
α-半乳糖苷酶在禽料中的应用研究进展
2010-08-09 11:14:28戴求仲
饲料工业 2010年2期
乔 伟 张 民 戴求仲 周 辉
豆粕、棉粕、菜籽粕等含有丰富的蛋白质,是家禽饲料中常见的主要蛋白原料,而豆科类种子、棉籽、菜籽、葵花籽中存在着一些由1个蔗糖与1个或多个半乳糖以α-1,6-糖苷键连接而成的不溶于水的低聚糖,如棉籽糖(三糖)、水苏糖(四糖)和毛蕊花糖(五糖)等。在大豆粕中棉籽糖和水苏糖的含量约为7.1%,占豆粕中碳水化合物总量的40%左右。棉籽粕蛋白质含量较高,氨基酸的平衡程度较好,是很好的蛋白质饲料来源,但在棉籽粕中的α-半乳糖苷含量比豆粕中的还要高。由于家禽不能分泌相应的降解酶,因此它们不能被家禽消化道的内源酶降解,只有经过消化道微生物发酵后才能被利用。这样不仅消化能大大下降,而且在发酵过程中会产生CO2、CH4和H2等气体,使家禽的采食量下降。另外,这些低聚糖还能刺激肠道蠕动,提高饲料通过消化道的速度,减少食糜在消化道停留的时间,从而降低养分的消化利用率。为了消除这些不溶性低聚糖的抗营养作用,长期以来一直采用高温、高压、高湿和延长时间等方法压榨大豆,或者采用有机溶剂萃取法消除这些低聚糖。但这些加工措施都不同程度地增加了豆粕的生产成本,而且效果并不理想。针对α-半乳糖苷的化学结构特点,国内外学者已成功开发出α-半乳糖苷酶制剂产品,并就α-半乳糖苷的抗营养作用机理以及α-半乳糖苷酶的生产应用进行了一系列研究,取得一定的成果。本文就α-半乳糖苷酶及其在家禽饲料方面的应用研究进行综述。
1 α-半乳糖苷结构、分布及抗营养特性
1.1 α-半乳糖苷及其在植物饲料中的分布
α-半乳糖苷是由一个蔗糖单位(果糖、葡萄糖)与一个(或多个)半乳糖分子以α-1,6糖苷键连接构成的寡糖(或低聚糖)类物质,又称低聚半乳糖,这些低聚糖类物质主要有棉籽糖(raffinose)、水苏糖(stachyose)和毛蕊花糖(verbascose),它们广泛存在于各种植物性饲料原料中,其中豆科植物中α-半乳糖苷含量最高,谷物中含量较低。表1为常见蛋白和能量饲料原料中α-半乳糖苷组成和含量。
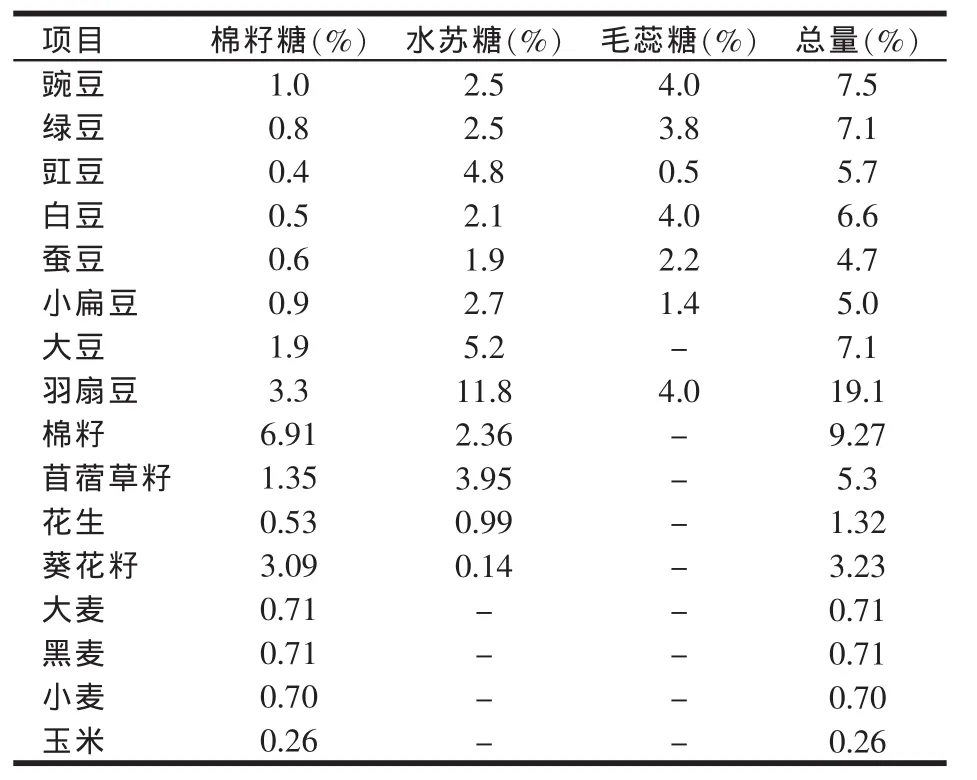
表1 常见蛋白和能量饲料原料中α-半乳糖苷的组成和含量
1.2 α-半乳糖苷对能量利用的不良影响
α-半乳糖苷不能被鸡的内源酶消化,也未发现鸡小肠黏膜中具有α-1,6半乳糖苷酶的活性,因此,在家禽小肠中没有分解α-半乳糖苷类物质中糖苷键的酶(Leske等,1993a),单胃动物饲料中的α-半乳糖苷,只有经后肠段消化道微生物发酵后才能被利用(Ohshima等,1997),这样由α-半乳糖苷提供给动物的净能就会少于其直接转化成单糖时所提供的能量,并且会产生CO2、NH3等气体,引起胀气 (Calloway等,1996;Leske 等,1999),甚至渗透性腹泻。有关 α-半乳糖苷抗营养作用的大量研究结果表明,α-半乳糖苷在体内大量积聚,达到一定量时,就会影响饲料的饲喂效果,对机体造成一定的影响(Anderson等,1995)。
经研究表明,用乙醇处理过的低寡糖豆粕进行试验,低寡糖豆粕的氮校正真代谢能值(TMEn)显著高于普通豆粕。Angel等(1988)利用强饲法对成年公鸡进行的一项试验结果也表明,低寡糖豆粕的TMEn确实高于普通豆粕。Coon等(1990)后来的另一项试验得出同样结果,同时还表明:碳水化合物的消化率从53.9%提高到67.3%,半纤维素和纤维素的消化率分别从9.2%和0%提高到61.6%和35.0%;低寡糖豆粕后肠道食糜的pH值要高于普通豆粕(Irish等,1995)。另外,还发现鸡对低寡糖豆粕日粮中α-半乳糖苷的全肠道消化率显著降低,而回肠表观消化率几乎没有变化,笔者认为这可能是由于α-半乳糖苷与肠道的营养物质牢固结合造成的。Leske等(1993b)仔细研究了棉籽糖或水苏糖含量与TMEn的回归关系,发现添加棉籽糖或水苏糖,或同时添加棉籽糖和水苏糖到抽提了寡糖的大豆粕日粮中,结果对大豆粕的代谢能产生了负面影响(见表2),经过回归分析得出了两个公式:
y=5 357.1-4 780.8x+2 572.8x2(r2=0.95)
其中:y为TMEn,x为棉籽糖的含量 (干物质基础,0.65%~1.05%)。
y=3 841.7-377.34x+46.225x2(r2=0.83)
其中:y为TMEn,x为水苏糖的含量 (干物质基础,0.36%~5.36%)。
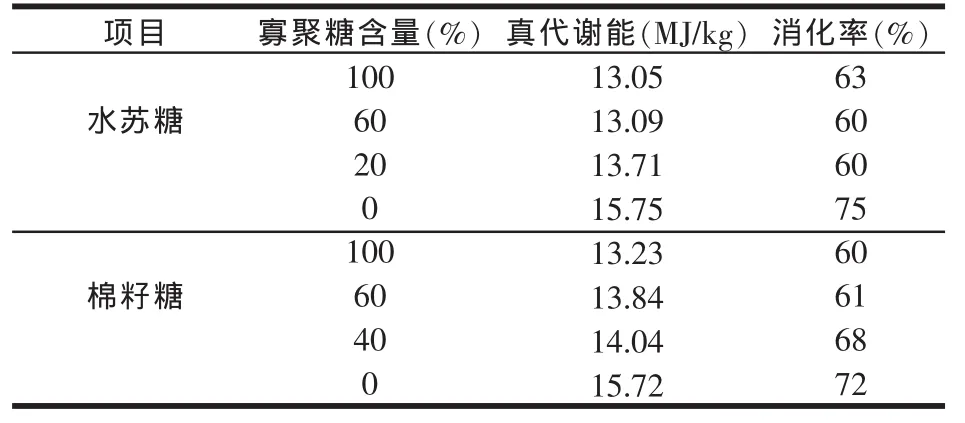
表2 提高棉籽糖或水苏糖含量对豆粕真代谢能(TME)及干物质(DM)消化率的影响
由表2可知,为了使豆粕获得较好的代谢能至少80%的水苏糖应被去除,而棉籽糖去除40%就显著提高代谢能值,去掉60%就能获得最好的效果。但从干物质消化率考虑,两者都应全部去除才有改善。
同时也有研究结果表明,由于α-半乳糖苷的存在使回肠末端的淀粉、粗蛋白质及无氮浸出物的消化率均有所下降。
1.3 α-半乳糖苷对家禽肠道pH值和食糜通过速度的影响
Goon等(1990)用乙醇作溶剂来浸提α-半乳糖苷进行肉鸡饲养试验,结果豆粕的代谢能增加了20%,其他非淀粉多糖,如阿拉伯木聚糖、果胶糖、β-葡聚糖等的消化率从5%提高到50%以上。另外还发现浸提α-半乳糖苷后,可减少饲料消耗量,增加消化时间和盲肠的pH值(见表3)。
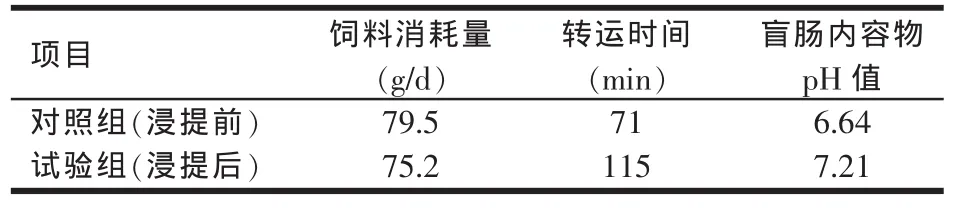
表3 α-半乳糖苷对肉鸡饲料消耗量、肠内运转时间和pH值的影响
Coon等在探讨α-半乳糖苷显著降低TMEn值的机理时发现,与普通豆粕相比,鸡对低寡糖豆粕中α-半乳糖苷的回肠表观消化率并没有变化,但是在全肠道的消化率显著降低。笔者认为,这是由于经过乙醇浸提后,棉籽糖和水苏糖在低寡糖豆粕中的含量非常显著地低于普通豆粕。这些残留的α-半乳糖苷可能与一些其它的物质非常牢固地结合在一起,从而导致了消化率的下降。经过比较后我们可以明显看出,α-半乳糖苷在回肠几乎不能被消化,而经过后肠的微生物作用后,全肠道的消化率明显提高,尤其是普通豆粕可以达到85%左右,因此,α-半乳糖苷的主要发酵部位是在后肠。笔者认为,普通豆粕的TMEn值较低的原因并不是由于微生物的水解能力不够,而恰恰相反,正是由于微生物的水解能力很强,改变了后肠的消化环境,非常突出的就是使后肠的pH值降低,食糜的通行速度过快。而低寡糖豆粕正相反,它的pH值较高,食糜的通过速度较慢,很可能创造了一个更适合分解碳水化合物的环境,从而使纤维素和半纤维素的消化率明显高于普通豆粕。这些被发酵的碳水化合物提供的挥发性脂肪酸可以作为能量被鸡利用。Irish等也发现,低寡糖豆粕的食糜在后肠的pH值有高于普通豆粕的趋势。
1.4 α-半乳糖苷对动物生长性能的影响
α-半乳糖苷对动物生长性能影响的研究结果争议很大。Trevin等将大豆寡糖的提取物重新添加到日粮中,而家禽的生长性能却没有受到任何影响。虽然在绝对值上,生长速度和饲料利用率有所提高,但是统计差异并不显著。lrish等的试验结果甚至表明去寡糖豆粕会明显降低日增重、饲料转化效率以及蛋白质的表观消化率。而Irish等的试验结果却说明肉鸡的生长性能与小肠食糜中寡糖的含量呈现负相关,即食糜中的寡糖含量越高,肉鸡的生长性能越差。
2 α-半乳糖苷酶的生理生化特性
2.1 α-半乳糖苷酶
α-半乳糖苷酶(EC3.2.1.22)属外切糖苷酶类,能专一地催化多糖、糖脂、糖蛋白中糖链末端的α-半乳糖苷键水解(Coon等,1990),它不仅能水解棉籽糖、水苏糖和毛蕊花糖等低聚糖,还能水解含α-半乳糖苷的杂多糖。α-半乳糖苷酶广泛存在于各种植物和动物体内,许多微生物的提取液中也发现有α-半乳糖苷酶活性。目前已从许多原核和真核生物中纯化得到α-半乳糖苷酶。α-半乳糖苷酶同其它糖苷酶一样属糖蛋白,其中蛋白质和糖苷的连接方式是通过低聚糖中的乙酰葡萄糖胺和蛋白质中的天冬氨酸连接的。不同方法制得的酶有不同的最适pH值和温度范围。一般细菌分泌的最适pH值范围是6.5~7.5,最佳温度在37~40℃,温度一般在50~60℃。同工酶之间以及同一种微生物在不同培养基上产生的α-半乳糖苷酶的稳定性也有显著差异。
α-半乳糖苷酶对于植物种子的萌发至关重要,其缺乏可能抑制种子的萌发。种子萌发初期,它首先催化含有α-半乳糖基的寡糖,如蜜三糖、水苏糖的水解,其终产物D-半乳糖通过糖酵解途径迅速转化和消耗,为种子的萌发提供最初的能量来源。后期,它主要参与细胞壁储藏多糖如半乳甘露聚糖等的水解。α-半乳糖苷酶的水解作用方式如下所示:
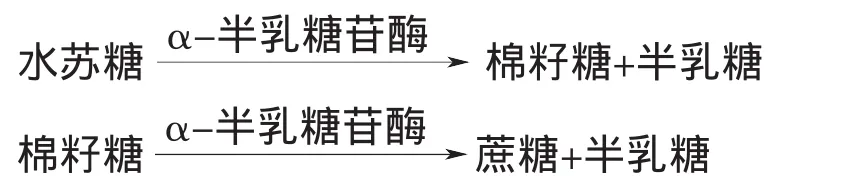
2.2 α-半乳糖苷酶的最适pH值
大多数α-半乳糖苷酶的最适pH值在4~7之间,丝状真菌及酵母菌分泌的最适pH值范围变化较大,相对于底物对硝基苯α-半乳吡喃糖苷(p-nitrophenylα-D-galactopyranoside,pNPG),在 4.5~8.0 之间。对于不同底物,最适pH值会发生相应的变化。咖啡豆来源的α-半乳糖苷酶对于人工底物pNPG的最适pH值为6.4;当底物是蜜二糖、蜜三糖或水苏糖时,最适pH值降到3.6~4之间。
2.3 α-半乳糖苷酶的底物专一性
α-半乳糖苷酶对于α-D-半乳糖苷键具有较广泛的底物专一性。咖啡豆的α-半乳糖苷酶不仅能够水解低分子量的底物,如pNPG、蜜二糖等,而且能够水解高分子量的寡糖(如水苏糖)和多糖(如半乳甘露聚糖)。
3 α-半乳糖苷酶的作用机理
3.1 控制肠道微生物发酵,增强免疫力
加入α-半乳糖苷酶后,抑制了肠道微生物的过度发酵,使肠道内的胺、甲酚等有害物质减少,同时可以提高内源性蛋白酶的活性,促进多肽类物质的产生,控制因饲料消化不良而引起的腹泻。酶制剂可以使多糖降解产生寡聚糖及其衍生物,这些物质可防止致病菌在肠道上聚集,减轻病原菌对机体的毒害,参与机体的免疫调节,增强免疫力。王春林(2004)研究表明,α-半乳糖苷酶的添加可以增加球蛋白水平及21日龄胸腺和脾脏的相对质量,表明α-半乳糖苷酶可增强机体的免疫功能。
3.2 破坏细胞壁结构,促进细胞内营养物质释放
细胞壁是由脂肪双分子层组成,其功能是调节细胞内的渗透压,参与细胞内外的物质交换,同时也能阻碍营养物质的吸收。加入酶制剂可破坏饲料中存在的植物细胞壁,使细胞的内容物淀粉、蛋白质释放出来,降低渗透压,破坏食糜周围的水化膜,使食糜运行的速度减慢,酶容易渗入食糜中,使酶有最佳的作用时间,增大食糜与小肠的接触面积,提高营养物质的吸收。
3.3 参与内分泌调节,影响血液中某些成分
酶制剂对一些激素具有调节作用,如甲状腺激素、胰岛素、生长激素、胰岛素样生长因子等,从而提高代谢水平。酶制剂影响内分泌的机理还不太清楚,可能有如下几条途径:①提高了营养物质的消化、吸收和利用,释放的营养物质引起相应的激素水平发生变化;②有的营养物质还有激素样活性,酶能促进或抑制某些激素的活性;③日粮中的蛋白质与多糖经酶制剂处理后,可能产生特殊活性的活性肽和寡糖,在体内有特殊的生理或药理作用,参与机体的内分泌调节,影响代谢。目前使用的酶制剂大多是来源于微生物发酵的粗制品,其成分复杂,除了含有水解酶外,还有培养基中多糖和蛋白质被降解的中间代谢产物,也包括微生物生长过程中的次生代谢物和小分子的代谢副产物,还有细菌的菌体,这些物质可能本身就含有一些调节机体神经内分泌功能的未知因子。研究发现,粗酶制剂可促进雏鸡血清中T3水平升高,而对T4水平的影响不大,说明酶制剂具有促进甲状腺素释放激素(TRH)的生理效用。禽类TRH-T4、T3轴通过生长激素(GH)对生长起作用;肝、肾组织的5'-脱碱酶的活性提高,促使T4转化为T3,从而增强机体的代谢功能。
4 α-半乳糖苷酶在禽料中的应用
4.1 α-半乳糖苷酶在禽料中的应用效果
Gdala等的研究结果发现,在羽扇豆粕的日粮中加入α-半乳糖苷酶,结果α-半乳糖苷回肠末端消化率从80%提高到97%,效果显著。Ghazi等(1997a、1997b)的两次试验结果均表明,α-半乳糖苷酶提高了豆粕的TME值和氮存留率,同时也使动物的日增重显著提高。Barcelona Autonoma大学曾用肉仔鸡作为试验材料,评价了α-半乳糖苷酶对玉米-豆粕型日粮中能量、蛋白质和其它养分消化率的影响,结果表明,在饲料中按照100 U/kg的量添加α-半乳糖苷酶,可以使代谢能提高5%~10%,氮存留率提高10%~15%,效果极其显著 (α-半乳糖苷酶的活力单位U定义为:37℃、pH值6.0的条件下,水解浓度为0.002 mol的对硝基苯-α-半乳吡喃糖苷,产生1 μmol对硝基苯酚所需要的酶量为一个酶活单位)。
西班牙巴塞罗纳Autonoma大学的一个肉鸡试验中,测定了以两个不同水平添加α-半乳糖苷酶对玉米-豆粕型日粮能量和氮存积的影响,结果见表4。
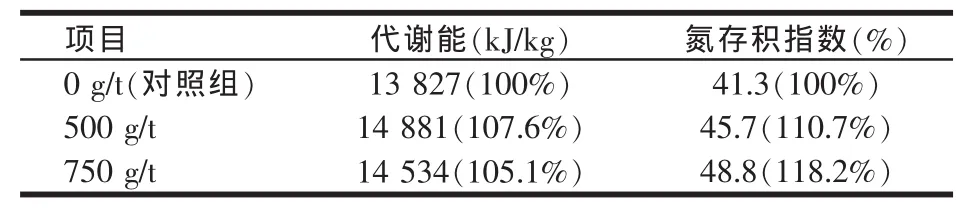
表4 不同α-半乳糖苷酶添加量对肉鸡日粮能量和蛋白质氮存积的影响
由表4可知,在添加了500 g/t α-半乳糖苷酶后,代谢能提高7.6%,氮的存积指数也提高了10.7%。表明添加α-半乳糖苷酶后,由于酶解作用,不仅对能量,而且对蛋白质消化率都有好处。为了测试酶和豆粕的用量关系,Autonoma大学还专门做了以下试验:酶的用量固定在500 g/t,改变日粮中豆粕的含量,来测定豆粕的表观消化能,结果见表5。

表5 α-半乳糖苷酶对不同含量豆粕日粮的表观代谢能影响
由表5可知,当豆粕比例较少,为30%、40%时,酶的作用比较明显,豆粕的表观代谢能提高了8%和11%;而当豆粕用量在50%、60%时,作用明显下降。Leske等早在1993年就指出,当α-半乳糖苷酶的水解作用低到一个临界水平时,对代谢能基本没有改善。豆粕在肉鸡日粮中正常用量一般不会超过40%,所以添加α-半乳糖苷酶后,预计可使豆粕的代谢能提高10%。
另外在酶的混合使用效果的研究中,Slominski等(1994)研究发现,用α-半乳糖苷酶和蔗糖酶并不影响双低菜籽粕和豆粕中非淀粉多糖(NSP)的消化率。Irish等(1995)将α-半乳糖苷酶和蔗糖酶加入日粮中进行研究,结果发现日粮的AMEn值反而有所下降。Igbasan等(1997)研究结果发现,在豌豆日粮中单独加入果胶酶没有明显的效果,但将果胶酶和α-半乳糖苷酶同时加入饲料中,肉鸡的生长速度和日粮的利用效果均有提高。
王春林等(2004)研究表明,在玉米-豆粕型日粮中添加α-半乳糖苷酶能显著提高肉公鸡的真代谢能、Met和Cys的真消化率,以及DM、OM、Ca和P的表观消化率,并提高21日龄肉公鸡采食量和日增重。有研究报道,以α-半乳糖苷酶为主的复合酶添加于肉鸡玉米-豆粕型日粮中,提高饲料效率1%~10%,胴体产量1%~20%,腹脂率降低10%~20%。表明α-半乳糖苷酶提高营养素的分配效率,有节省蛋白质和合成氨基酸的作用,降低饲料成本。
关于α-半乳糖苷酶的报道,之所以出现一些不一致的结果,这可能是由于不同的试验所用的试验条件、试验动物、日粮结构、α-半乳糖苷的水平等因素都不同,况且酶的来源、活力等又不相同,同时,酶引起的寡糖水解水平与日粮能量和蛋白质消化率提高之间存在一定的关系。
4.2 α-半乳糖苷酶最适添加量探讨
饲料中α-半乳糖苷酶最适添加量与配方原料中α-半乳糖苷的含量及α-半乳糖苷酶的活性有关。国外有学者做了日粮中豆粕比例为40%时酶不同添加量对豆粕表观代谢能的影响试验,结果见表6。

表6 不同α-半乳糖苷酶添加量对豆粕表观代谢能的影响
由表6可知,添加酶后,豆粕的代谢能均有不同程度的改善,可以明显地看出,用量500 g/t效果最好;当α-半乳糖苷酶的水解作用低于一定的阈值水平时,代谢能值不会明显提高。在含大豆粕40%的日粮中添加α-半乳糖苷酶500 g/t为宜,增大添加量反而降低大豆粕的代谢能值。然而,不同酶添加水平之间大豆粕代谢能值的增高并无显著差异。但豆粕在肉鸡饲粮中正常用量一般不会超过40%,在肉鸡日粮中采用实用水平的大豆粕时向日粮中添加α-半乳糖苷酶制剂,仍可期望大豆粕的代谢能值有8%~10%的提高。在豆粕比例为40%的饲粮中分别添加0、500、1 000、1 500 g/t的α-半乳糖苷酶对表观代谢能的研究表明,添加较低剂量的酶(500 g/t)是最经济有效的。
5 结语
豆粕、棉粕、菜粕等植物性蛋白原料中的α-半乳糖苷含量较高,这不仅降低了饲料的代谢能,影响了氮的存留和养分的消化吸收,而且也不利于家禽的生长健康,降低了家禽的生长性能和肉质。α-半乳糖苷的稳定性较高,一般的饲料加工方法难以破坏其结构,而加入α-半乳糖苷酶后,可以明显促进营养物质的消化吸收,提高饲料的代谢能值和氮存留率,改善动物生产性能,降低饲料成本。在实用日粮配方中利用这一提高的最好方法是修改大豆粕的代谢能和有效氨基酸模型。在加酶后饲料配方中,通过修改饲料原料营养含量值,可在既降低饲料成本,又提高家禽的生产性能的同时,还能节约饲料资源、减少环境污染。通常,可将加酶后粕类的代谢能值和有效氨基酸值提高5%~10%,然后带入配方数据库进行计算。
[1]王春林,陆文清,李德发.玉米-豆粕型日粮中添加α-半乳糖苷酶对肉鸡营养物质利用、生产性能、血清参数和器官重量的影响[M].中国畜牧兽医学会动物营养学分会——第九届学术研讨会论文集,2004.
[2]Anderson R L,W J Wolf.Compositional changes in trypsin inhibitors phytic acid saponins and isoflavones related to soybean processing[J].J.Nutr.,1995,125:5815-5885.
[3]Angel C R,Y L Sell,D R Zimmerman.Autolysis of α-galactosides of defatted soy flakes[J].Food Chem.,1988,36:542-546.
[4]Calloway D H,D J Colasito,R D Matthews.Gases produced by human intestinal microflora[J].Nature,1996,212:1238-1239.
[5]Coon C A,K L Leske,O Akavanichan,et al.Effect of oligasac charide-free soybean meal on true metabolizable energy and fibre digestion in adult roosters[J].Poultry Sci.,1990,69:787-793.
[6]Ghazi S,J A Rooke,H Galbraith,et al.Effect of adding protease and alpha galactosidase enzymes to soybean meal on nitrogen retention and true metabolizable energy in broilers[J].Bri.Poult.Sci.,1997a,38:S28.
[7]Ghazi S,J A Rooke,HGalbraith,et al.Effect of feeding growing chicks semi-purified diets containing soybean meal and amounts of protease and alpha-galactosidase enzymes[J].Bri.Poult.Sci.,1997b,38:S29.
[8]Igbasan F A,W Guenter,B A Slominski.The effect of pectinase alpha-galactosidase supplementation on the nutritive value of peas for broilers chickens[J].Can.J.Anim.Sci.,1997,77:537-539.
[9]Irish G G,G W Barbour,H L Classen.Removal of the alphagalactosidase of sucrose from soybean meal using ethier ethanol extraction or exogenous alpha-galactosidase and broiler performance[J].Poult.Sci.,1995,74:1484-1494.
[10]Kuo T M,J F V Middlesworth,W J Wolf.Content of raffinose oligosaccharides and sucrose in various plant seeds[J].J.agricul.Food Chem.,1988,32-36.
[11]Leske K L,C J Jevne,C N Coon.Effect of oligosaccharide additions on nitrogen-corrected true metabolizable energy of soy protein concentrate[J].Poultry Sci.,1993b,72:664-668.
[12]Leske K L,C J Jevne,C N Coon.Extraction methods for removing soybean alpha-galactosides and improving true metabolizable energy for poultry[J].Anim.Feed Sci.Technol.,1993a,41:74-78.
[13]Leske K L,C N Coon.Hydrogen gas production of broiler chicks in response to soybean meal and a-galactoside free,ethanolextracted soybean meal[J].Poult.Sci.,1999,78:1313-1316.
[14]Ohshima T,G T Murray,W D Swaim,et al.Alpha-galactosidase Adeficient mice:a model of Fabry disease[J].Pro.Natl.Acad.Sci.,USA,1997,94:2540-2544.
[15]Slominski B A,L D Campbell,W Guenter.Oligosaccharides in canola meal and their effect on non-starch polysaccharide digestibility and true metabolizable energy in poultry[J].Poult.Sci.,1994,73:156-162.
猜你喜欢
动物营养学报(2022年7期)2022-08-09 09:38:52
食品工业(2022年2期)2022-03-09 01:48:22
中国饲料(2020年16期)2020-11-06 06:57:54
吉林农业(2019年3期)2019-01-06 06:06:08
动物营养学报(2018年12期)2018-12-13 06:01:48
动物营养学报(2018年7期)2018-07-16 03:05:30
饲料博览(2016年6期)2016-04-05 14:11:14
中国药业(2014年12期)2014-06-06 02:17:29
食品与发酵工业(2012年5期)2012-04-14 15:43:05
饲料博览(2012年10期)2012-04-13 21:00:34
