
二倍体和三倍体虹鳟外周血细胞的比较研究
2010-08-09 06:45陈侠君王炳谦范兆廷
东北农业大学学报 2010年1期
陈侠君,王炳谦,刘 奕,范兆廷*
(1.东北农业大学动物科学技术学院,哈尔滨 150030;2.中国水产科学研究院黑龙江水产研究所,哈尔滨 150070)
虹鳟(Onchorynchus mykiss)属于鲑形目(Salmoniformes),鲑科(Salmonidae),是世界性的冷水性养殖鱼类。由于育性不良或者不育,三倍体鱼类可以将能量完全用于营养生长,性成熟后在生长和肌肉质量方面表现出较强的优势。三倍体育种在鲤鲫杂种及虹鳟、团头鲂、罗非鱼、牡蛎、鲍鱼和扇贝等水产动物中得到广泛的应用[1-5]。虹鳟的卵子体积大、胚胎发育时间长,易于进行各种染色体倍性的遗传操作,三倍体苗种生产容易,易于普及推广。三倍体虹鳟在幼鱼阶段时生长速度稍慢,但雌性的性腺不发育,几乎没有成熟死亡现象,三倍体虹鳟的个体大,肌肉质量好,可加工为上乘的三文鱼;同时三倍体虹鳟对水霉病和传染性造血器官坏死病(IHN)抵抗力强[6-8],受到生产者和消费者的青睐。
在对多倍体的血液研究中,往往侧重于红细胞的DNA含量或细胞核体积的测量,对于基因组成倍增加后,红细胞核的形态发生的变异及变异的原因和构成外周血的主要成分白细胞的变化则罕有研究。文章对二倍体和三倍体虹鳟的红细胞、白细胞的形态进行了比较分析,以其为丰富虹鳟血细胞形态学、血液生理学、人工诱导三倍体虹鳟以及三倍体虹鳟的鉴定提供理论基础。
1 材料与方法
1.1 材料
三倍体虹鳟(Onchorynchus mykiss)由中国水产科学研究院黑龙江水产研究所王炳谦课题组通过静水压力方法获得并培育;二倍体虹鳟系同期人工繁殖获得。二者均饲养于中国水产科学院黑龙江水产研究所渤海冷水鱼类实验站。2008年3月随机选择体表无伤、发育正常的三倍体及同期二倍体虹鳟各20尾,其体重及叉长见表1。

表1 试验用虹鳟基本情况Table 1 Basic circumstance of Onchorynchus mykiss
1.2 方法
用1%肝素浸润的1 mL注射器从鱼尾静(动)脉采血,制作血涂片,自然风干后用甲醇固定,瑞氏染液染色,清水冲洗干净,自然干燥。
在光学显微镜下观察各种细胞的形态特征,测量各种白细胞、成熟红细胞和成熟红细胞核的长径(b)与短径(a)。每个样本各测量50个细胞,取其平均值,根据公式S=πab/4计算红细胞和红细胞核的面积;根据公式V=a2b/1.91计算白细胞、红细胞和红细胞核的体积[9],根据公式核质比=细胞核体积/细胞质体积[10]计算红细胞的核质比;统计三倍体虹鳟1 000个红细胞中异常红细胞的百分比,并对异常红细胞的种类及所占比例进行分析;计数二倍体和三倍体虹鳟血涂片中的白细胞,并分类。
1.3 数据处理与分析
试验数据均采用SPSS进行统计学处理,采用t检验进行差异显著性分析。
2 结果与分析
2.1 二倍体与三倍体虹鳟红细胞大小及形态异常红细胞的比较
二倍体与三倍体虹鳟的红细胞及核的大小测量结果及二者之间的差异显著性检验结果见表2。

表2 二倍体和三倍体虹鳟红细胞及其细胞核大小比较Table 2 Comparison on size of erythrocyte and cellular nucleus in diploid and triploid rainbow trout (μm)
由表2可以看出,三倍体虹鳟各项数值都高于二倍体,三倍体虹鳟红细胞长径、红细胞短径、红细胞体积、红细胞面积、核长径、核体积以及核面积分别为二倍体的1.30、1.11、1.58、1.43、1.38、1.45、1.34倍,差异极显著(P<0.01);三倍体虹鳟红细胞核短径与二倍体差异不显著(P>0.05)。其中二倍体个体的红细胞长径的平均值为 12.12 μm,而三倍体个体的红细胞长径的平均值为15.71 μm;三倍体红细胞短径/长径比值为0.57,小于二倍体的比值0.67;三倍体虹鳟和二倍体虹鳟的核质比均为0.82。三倍体虹鳟中63%的红细胞呈长椭圆形,17%的红细胞有哑铃形细胞核或分裂的细胞核,其中哑铃形红细胞占所有形态异常红细胞的47.92%,均等分裂的红细胞占20.83%,不均等分裂红细胞占31.25%。本研究中未见二倍体虹鳟的红细胞有哑铃形细胞核和分裂的细胞核现象。
2.2 二倍体与三倍体虹鳟的白细胞分类记数及大小比较
由表3可知,虹鳟白细胞构成特点是以血栓细胞为主,其次为淋巴细胞、嗜中性粒细胞,单核细胞极少,没有发现嗜酸性粒细胞和嗜碱性粒细胞,其中二倍体和三倍体在淋巴细胞和嗜中性粒细胞上有较大差异。从表4中可以看到,三倍体虹鳟白细胞的长径和短径都大于二倍体;嗜中性粒细胞、单核细胞的体积与倍性的1.5:1比例相接近;淋巴细胞、血栓细胞的大小虽然没有表现出相应的比例,但三倍体虹鳟的淋巴细胞、血栓细胞与二倍体虹鳟相应细胞之间在体积上存在显著性差异。
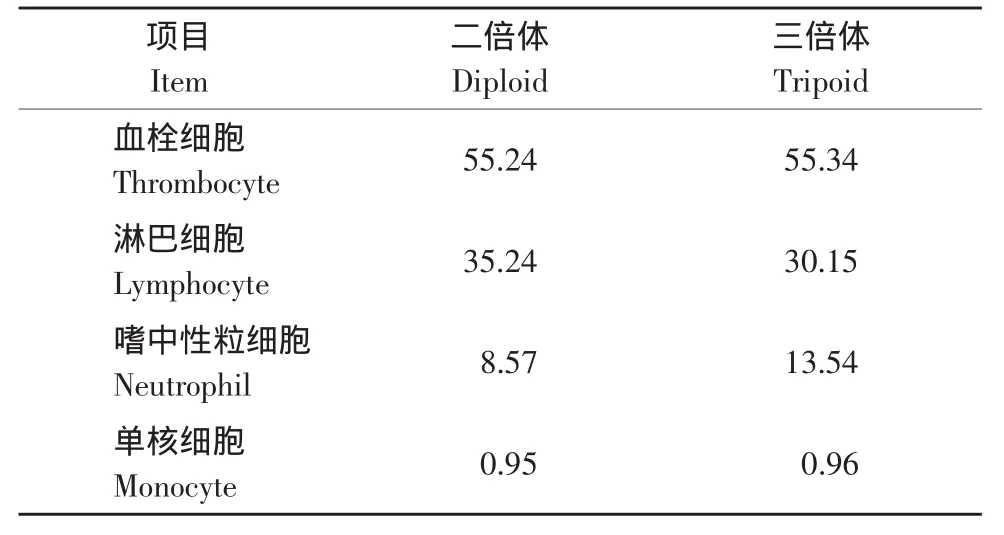
表3 白细胞分类计数值Table 3 Differential leucocyte counts(DLC)(%)

表4 二倍体和三倍体虹鳟白细胞大小比较Table 4 Comparison on size of leucocyte in diploid and triploid rainbow trout (μm)
2.3 二倍体与三倍体虹鳟血细胞形态的观察
2.3.1 红细胞形态观察
在二倍体和三倍体虹鳟的血涂片上均可以观察到原红细胞、幼红细胞以及成熟的红细胞,偶然可见衰老的红细胞和核影细胞。
两种倍性虹鳟的原红细胞呈圆形或近圆形,细胞核也呈圆形,居中,核质比大,染色质成网状,染成浅紫色,细胞质较少染成浅蓝色(见图版Ⅰ-1、2);幼红细胞圆形或略成椭圆形,和成熟的红细胞形状相差不大,常成群出现,其核比成熟的红细胞稍大,圆形或椭圆形,核质比较大,染色质结构疏松,着色较浅,细胞质明显增多,染成较浅蓝色(见图版Ⅰ-3、4)。

图版Ⅰ 二、三倍体虹鳟血细胞的形态PlateⅠ Morphology of blood cells in diploid and triploid rainbow trout
无论是三倍体还是二倍体,成熟的红细胞长椭圆形,表面光滑,核椭圆形,位于细胞中央,染色质致密,核质比小,染成绛紫色,细胞质染成非常浅的蓝色(见图版Ⅰ-5、6);三倍体中可见到一些异常的红细胞,有的呈哑铃状,细胞质内陷(见图版Ⅰ-7),有的细胞内有两个分开的小核,一大一小(见图版Ⅰ-8),或大小均等(见图版Ⅰ-9),有的是细胞质已分开但核没分开(见图版Ⅰ-10)。
衰老的红细胞较正常的红细胞大,核也大,染色质成疏松的网状,染成浅紫红色,细胞质着色很浅接近白色(见图版Ⅰ-11、12)。
核影红细胞似裸核,染色质疏松网状,染成淡紫红色,无细胞质(见图版Ⅰ-13、14)。
2.3.2 白细胞形态观察
血栓细胞:细胞极小,是血细胞中最小的细胞,呈圆形、不规则圆形、短棒状或纺锤形;细胞核占细胞的大部分,呈肾形(见图版Ⅰ-15、16)、纺锤形(见图版Ⅰ-17、18)、圆形(见图版Ⅰ-19、20)或短棒状(见图版Ⅰ-21、22);核中有稠密的染色质,着色很深,染成深紫色;细胞质仅一薄层,或有时不清,绕核零星分布,边界模糊,淡蓝色或透明色。血栓细胞在血涂片中常单个、成队、数个或数十几个聚集在一起(见图版Ⅰ-23、24),在显微镜下十分容易辨认。在三倍体中还能观察到很多哑铃形的血栓细胞(见图版Ⅰ-25),另外,三倍体以纺锤形血栓细胞为主,而二倍体以圆形血栓细胞为主。
淋巴细胞:仅比血栓细胞略大,细胞有大有小,多为圆形、肾形或不规则形状(见图版Ⅰ-26、27);多数核有清晰可见的缺刻(见图版Ⅰ-28);核染色质浓密,呈粗网状,着色较深,染成紫色。淋巴细胞核几乎占据整个细胞,位于细胞中央或与质膜相切;细胞质较少,在核外围成窄环或仅在核的一侧出现,呈弱嗜碱性,染成淡蓝色或天蓝色;有的边缘平整,有的可见钝形的胞质突起。胞质极少的淋巴细胞与裸核状球形血栓细胞有时很难区分。
嗜中性粒细胞:细胞呈圆形或不规则圆形,边缘较为平整。核的形状不十分规则,呈肾形、马蹄形、螺旋形或分叶形,位于细胞中央,或偏居于细胞一侧,常与质膜相切,约占细胞的1/3。核中染色质浓密,染色较深;胞质着色较浅,染成灰蓝色,有的细胞能看见其中有微小的蓝紫色颗粒。血涂片上,嗜中性粒细胞有时与单核细胞难以区分,但前者胞体较圆,表面较光滑,胞核较小,胞质染色较淡。二倍体和三倍体中均有中有螺旋形(见图版Ⅰ-29、30)、二分叶(见图版Ⅰ-31、32)、三分叶形(见图版Ⅰ-33、34)核嗜中性粒细胞,而二倍体中肾形核嗜中性粒细胞(见图版Ⅰ-35)、三倍体中有四分叶形(见图版Ⅰ-36)、马蹄形(见图版Ⅰ-37)、杆状(见图版Ⅰ-38)核。
单核细胞:在外周血液中数量很少,圆形或近圆形,边缘较粗糙,核呈不规则的圆形或不规则多边形,较大,位于细胞中央或偏心位,核中染色质分布疏松,成网状,胞质丰富,着色较浅,染成灰蓝色,有蓝紫色颗粒分布(见图版Ⅰ-39、40)。
3 讨论与结论
由于基因组大小的不同,不同倍性细胞核体积比值存在较大的差异。马涛等报道二倍体与四倍体虹鳟细胞核体积的比值为1:2.42[11];朱蓝菲等在研究人工同源和异源三倍体鲢时,发现同源三倍体红细胞核体积为二倍体的1.63倍[9];鲢×鳙及鲢×团头鲂异源三倍体红细胞核体积则分别是二倍体的2.04倍和1.85倍。俞小牧等则发现异源四倍体、四倍体和倍间三倍体白鲫(Carassius curieri)的细胞核体积分别是二倍体的1.83,1.98和1.43倍,接近理论值2.00倍和1.50倍[12]。本研究结果表明,三倍体虹鳟的核体积约为二倍体的1.5倍,表明通过测量红细胞核体积能较准确反映出多倍体的倍性。
一般认为,细胞大小与DNA的含量或染色体数量有关。为维持恒定的核质比例,随着染色体数目增多,多倍体的细胞及细胞核通常要比二倍体大[13]。本研究结果表明,虹鳟三倍体的核体积约为二倍体的1.5倍,并且三倍体和二倍体的红细胞体积分别为658.50和417.25,比值接近1.5,依倍性增加而且同比例增大。
研究发现三倍体虹鳟的红细胞形态与二倍体相比发生了一定的变化,有近17%的哑铃状或双核红细胞。俞小牧等对异源四倍体白鲫及倍间三倍体白鲫的研究也发现异常红细胞(哑铃形或双核)出现率分别为7.6%和10.12%[12];刘巧等报道的异源四倍体鲫鲤有33%的红细胞核为哑铃形、三倍体湘云鲫有25%的红细胞核为哑铃形[1]。因此可将该现象作为区别多倍体与二倍体的一个重要标志。虹鳟三倍体中产生异常红细胞的原因目前尚不能得到合理的解释,可能是由于三倍体红细胞存在一些异常无丝分裂,而使得部分红细胞停留在无丝分裂的不同阶段。若要对此现象作出明确的结论,还需要对三倍体鱼类造血器官的生理特点以及红细胞的发生过程进行深入研究,以寻找其组织学和细胞学证据。
虹鳟外周血白细胞中的血栓细胞和淋巴细胞占有相当大的比例,在二倍体和三倍体虹鳟中均未发现嗜酸性粒细胞和嗜碱性粒细胞。在鱼类外周血液中是否存在嗜酸性粒细胞和嗜碱性粒细胞,以往的研究已有较多论述[14-18]。笔者倾向于认为嗜酸性粒细胞和嗜碱性粒细胞客观存在,但数量较少,或者我们制片过程中导致了嗜酸性粒细胞和嗜碱性粒细胞解体而无法得见。
白细胞的主要作用是保护机体,抵御病害的侵袭,在机体的免疫反应中起主要作用[19]。白细胞数量的差别可用于解释鱼类的抗病和在特定环境下的耐受能力。三倍体虹鳟白细胞中嗜中性粒细胞和淋巴细胞都显著多于二倍体,可能与三倍体抗病能力强有关。
二倍体虹鳟的血栓细胞在血涂片上表现出各种形态:有圆形、肾形、纺锤形、短棒形;除以上形状外,三倍体虹鳟还有哑铃形;四倍体鲫鲤的血栓细胞有卵圆形、长杆形、大头钉形三种形态[1];欧洲鳗鲡表现为5种形态[20];这显示了血栓细胞的形态因种类而异。
Barber等认为许多淋巴细胞的体积是连续变化的,不能用“大”“小”来界定[17]。Ellis指出,将淋巴细胞分为“大”“小”是武断的,因为它们可能代表了同一种细胞的不同功能状态,而不大可能是具有不同功能的两种细胞[18]。因此,有研究者将其归为一类进行描述[21-23]。但也有人认为它们是具有不同功能的两类细胞[24-25]。在本试验中,通过观察和测量,淋巴细胞的大小和染色深浅有一定的连续性,而且较大者胞质染色较深,这与“未成熟的细胞往往较已成熟者大(巨核细胞系除外)”,“细胞愈年幼,RNA含量愈多,与瑞氏染液中的碱性美蓝有亲和力,浆染色愈蓝”的规律相符合[26],因此可以认为是淋巴细胞的不同功能状态,故应归为一类。
二倍体虹鳟嗜中性粒细胞有肾形、螺旋形、二分叶、三分叶形核,而三倍体中无肾形核、有马蹄形、螺旋形、二分叶、三分叶,四分叶形核。肾形核、马蹄形核和四分叶核可作为区分二倍体虹鳟与三倍体虹鳟的另一重要鉴定标志。
由此可知,两种倍性虹鳟的红细胞、嗜中性粒细胞、血栓细胞的形态均存在差异,尤其以红细胞的差异最为明显。三倍体虹鳟外周血中近17%的红细胞有哑铃形细胞核或双核而二倍体虹鳟中未观察到这种畸形细胞,因此这种现象可以作为区别三倍体与二倍体的一个重要标志。
[1]刘巧,王跃群,刘少军,等.不同倍性鲫鲤鱼血液及血细胞的比较[J].自然科学进展,2004,14(10)∶1111-1117.
[2]龙强,宗虎民.虹鳟鱼三倍体的育种技术[J].北京水产,2004(5)∶36-37.
[3]邹曙明,李思发,蔡完其,等.团头鲂同源四倍体、倍间三倍体与二倍体红细胞的形态特征比较[J].中国水产科学,2006,13(6)∶891-895.
[4]Varad K,冯益民.全雌三倍体莫桑比克罗非鱼的生产[J].国外水产,1990(4)∶22-24.
[5]王昭萍,姜波,孔令峰,等.利用四倍体与二倍体杂交规模化培育全三倍体太平洋牡蛎苗种[J].中国海洋大学学报,2004,34(5)∶742-746.
[6]马作析.鱼类多倍体及在养殖中应用的可能性[J].现代渔业信息,1991,6(4)∶6-10.
[7]楼允东.鱼类育种学[M].北京∶中国农业出版社,1999∶117-152.
[8]Benfey T J,Sutterlin A M.The haematology of triploid landlocked Atlantic salmon,Salmo salar L.[J].J Fish Biol,1984,24∶333-338.
[9]朱蓝菲,桂建芳,梁绍昌,等.人工同源和异源三倍体鲢的红细胞观察[J].水生生物学报,1992,16(1)∶84-87.
[10]倪海儿,杜立勤.东海鳓.卵巢发育的组织学观察[J].水产学报,2001,25(4)∶317-322.
[11]马涛,朱才宝,朱秉仁.热休克诱导虹鳟四倍体[J].水生生物学报,1987,11(4)∶329-335.
[12]俞小牧,陈敏容,杨兴棋,等.人工诱导异源四倍体和倍间三倍体白鲫的红细胞观察及其相对DNA含量测定[J].水生生物学报,1998,22(2)∶291-295.
[13]Small S A,Benfey T J.Cell size in triploid salmon[J].Exp Zool,1987,241∶339-342.
[14]赵海鹏,赵海涛,金丽,等.华鲮外周血细胞纤维结构观察[J].水产科学,2004,24(6)∶24-27.
[15]Thorgaard G H,Gall G A E.Adult triploids in a rainbow trout family[J].Genetics,1979,93∶961-973.
[16]Arai K,Matsubara K,Suzuki R.Production of polypioids and viable gynogens using spontaneously occurring tetraploid loach,Misgumus anguillicaudatus[J].Aquaculture,1993,117∶227-235.
[17]Barber D L,Millis Westermann J E,White M G.Th blood cells of the antarcitic icefish Chaenocephalus aceratus Lonnberg light and electron microscopic abservations[J].Fish Boil,1981,19∶11-28.
[18]Ellis A E.The leucocytes of fish∶A review[J].Fish boil,1977(11)∶453-491.
[19]林浩然.鱼类生理学[M].广州∶广东高等教育出版社,1999∶82-87.
[20]周玉,郭文场,杨振国,等.欧洲鳗鲡外周血细胞的显微和超微结构[J].动物学报,2002,48(3)∶393-401.
[21]Roubal F R.Blood and other possible inflammatory cells in the sparid Acanthopagrus australis(Gunther)[J].Fish Biol,1986,28∶573-593.
[22]胡成钰.鳙鱼外周血细胞的细微结构[J].江西科学,1990,8(3)∶20-28.
[23]冯怀亮,李文武,王铁恒.鲤鱼血细胞显微和亚显微结构的观察[J].水产学报,1991,15(3)∶241-244.
[24]Parrott D M V,Desousa M A B.Thymus-dependent and thymusindependent populations.Origins,migratory patterns and life span[J].Clin Exp Immunol,1971(8)∶663-684.
[25]周玉,潘风光,李岩松,等.达氏鳇外周血细胞的形态学研究[J].中国水产科学,2006,13(3)∶480-484.
[26]刘志洁,宗英.野生动物血液细胞学图谱[M].北京∶科学出版社,2002.
猜你喜欢
中国海洋大学学报(自然科学版)(2023年2期)2023-02-21
科学养鱼(2021年11期)2022-01-11
养殖与饲料(2020年8期)2020-02-17
食品与生活(2019年12期)2019-05-15
生命科学研究(2018年1期)2018-05-29
江苏农业科学(2017年16期)2017-10-27
河北渔业(2017年9期)2017-10-16
现代农业科技(2017年8期)2017-06-10
中国饲料(2017年7期)2017-01-17
中国果业信息(2015年6期)2015-01-23
