
新基因MAGE-n表位肽与人HSP70融合基因的构建及表达纯化
2010-06-08 10:31张秀敏史琪琪李增山叶菁王军伟林慧曲萍胡沛臻隋延仿
中国医药生物技术 2010年4期
张秀敏,史琪琪,李增山,叶菁,王军伟,林慧,曲萍,胡沛臻,隋延仿
肿瘤多肽疫苗因其具有化学性质稳定、易于制备和保存、肿瘤型别通用性强、治疗费用相对较低等优势而成为肿瘤免疫治疗的新方法。众所周知,疫苗疗效的关键在于其抗原,而有效的抗原递呈是肿瘤免疫治疗中不可缺少的重要环节。MAGE-n 基因是我室首次发现的 MAGE 家族中的新成员[1],肝癌阳性率达 40%,该基因的表达模式与其他MAGE 基因相同,可作为肿瘤特异性免疫治疗的靶分子。后续研究中,我们应用表位预测结合免疫重建等技术筛选出一条具有特异性免疫活性的HLA-A2 限制性 CTL(cytolytic T lymphoeyte)表位 MAGE-n(159-167)QLVFGIEVV,经免疫学实验证实,体内外均可有效地诱导特异性 CTL 的产生,有效杀伤肿瘤细胞[2]。文献[3]和我们以往的工作表明,将 HSP70(heat shock protein,HSP)与肿瘤肽复合物联合制成疫苗能够激发抗原特异的CTL 反应。本研究选择 HSP70 为载体,构建MAGE-n 表位肽与 HSP70 的融合基因,并进行表达纯化。期望利用 HSP70 的分子伴侣作用加强APC(antigen presenting cell)细胞对抗原的加工递呈,以增强其免疫原性,为构建新型的肿瘤疫苗提供更有效的抗原奠定实验基础。
1 材料和方法
1.1 材料
菌种E.coliBL21(DE3)、原核表达载体pET-28a(+)、含 HSP70 基因全长片段的质粒pCDNA3.1(+)/HSP70 均由本室保存;质粒小样快速提取试剂盒和小量 DNA 片段快速胶回收试剂盒购自北京博大泰克生物基因技术有限责任公司;Pyrobest DNA 聚合酶、T4 连接酶、限制性内切酶购自日本 TaKaRa 公司;Ni Sepharose 6FF 亲和层析填料购自美国 GE 公司。
1.2 方法
1.2.1 采用 PCR 法构建 QLVFGIEVV 与 HSP70的融合基因 根据引物设计原则以及 HSP70 基因的核苷酸序列,分别设计上游引物 Mn70-1 及下游引物 Mn70-2,在下游引物中加入 QLVFGIEVV的核苷酸序列(共 27 个碱基),并在引物两端分别加入 Sac I、Hind III 酶切位点,引物由上海生工生物工程技术有限公司合成。Mn70-1:5’ CCG GAG CTC ATG GCT CGT GCG GTC GGG ATC 3’;Mn70-2:5’ CGG AAG CTT TCA GAC GAC CTC TAT GCC GAA CAC TAA TTG CTT GGC CTC CCG GCC GTC 3’。此序列中方框标记处分别为Sac I、Hind III 酶切位点,划线部分为表位肽QLVFGIEVV。反应条件为:94℃预变性 5min,94℃变性 1min,57℃退火 1min,72℃延伸150 s,循环 1 次;94℃变性 1min,67℃退火及延伸 210 s,循环 30 次,72℃延伸 7min,PCR产物进行琼脂糖凝胶电泳,回收。
1.2.2 重组质粒 pET-28a(+)/QLVFGIEVVHSP70 的构建 用 Sac I、Hind III 双酶切 PCR扩增的融合基因 QLVFGIEVV-HSP70 和表达载体pET-28a(+)进行琼脂糖凝胶电泳。分别回收目的片段和表达载体,以 T4 DNA 连接酶于 16℃连接过夜,转化感受态E.coliBL21(DE3)。卡那平板筛选阳性克隆。提取阳性克隆菌质粒,以 Sac I和 Hind III 酶切和 DNA 序列分析鉴定阳性克隆。
1.2.3 融合基因 QLVFGIEVV-HSP70 的诱导表达及纯化 将含 pET-28a(+)/QLVFGIEVV-HSP70质粒的E.coliBL21(DE3)接种于 2 ml LB/Kan+的培养液中,37℃振荡培养至OD600约 0.6,加入终浓度为1mmol/L 的 IPTG 诱导,不加 IPTG 者为阴性对照,继续培养 4 h 取出,离心收菌,加入SDS-PAGE 上样缓冲液,混匀,煮沸 5min,在离心半径 8.4cm,12 000 r/min 条件下离心 10min,取上清经 SDS-PAGE 鉴定表达融合蛋白的菌落。
对 SDS-PAGE 鉴定有 QLVFGIEVV-HSP70融合蛋白表达的菌落进行大量(100~150 ml)扩增诱导,在离心半径 8.4cm,5 000 r/min,4℃条件下离心 5min,将菌体重悬于 10 ml 细菌裂解缓冲液(50mmol/L 磷酸缓冲液,pH 7.5),超声破碎,18 000×g,4℃离心 10min,取上清过 Ni Sepharose 6FF 亲和层析柱,平衡液包含 40mmol/L咪唑,0.5 mol/L NaCl,50mmol/L 磷酸缓冲液,pH 7.4;洗脱液包含 300mmol/L 咪唑,0.5 mol/L NaCl,50mmol/L磷酸缓冲液,pH 7.4。用 10%SDS-PAGE 电泳鉴定。
2 结果
2.1 MAGE-n159-167(QLVFGIEVV)表位肽与 HSP70融合基因的构建
以 pCDNA3.1(+)/HSP70 质粒为模板,在下游引物的 5’ 端引入 27 个碱基的模拟表位基因,经 PCR 将 QLVFGIEVV 与 HSP70 的序列进行连接,获得了 2.0 kb 的融合基因,与预期目的片段大小一致(图1)。
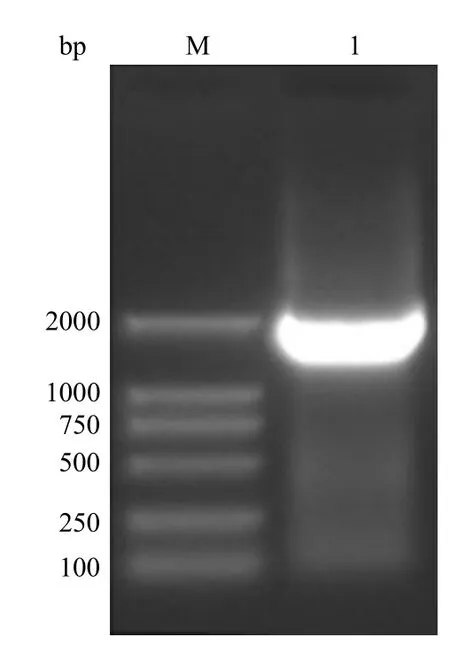
图1 QLVFGIEVV-HSP70 融合基因 PCR 结果的琼脂糖凝胶电泳Figure 1 Agarose gel electrophoresis of QLVFGIEVV-HSP70 fusion gene PCR product
2.2 原核表达载体 pET-28a(+)/QLVFGIEVV-HSP70的鉴定
pET-28a(+)/QLVFGIEVV-HSP70 重组质粒经 Sac I/Hind III 双酶切后,从 pET-28a(+)载体上消化释放出唯一的一条约 2.0 kb 的酶切片断,与目的片断大小相符(图2)。其 cDNA 序列测定结果表明:肝癌抗原肽基因序列已连接于 HSP70的 3’ 末端,HSP70 序列与基因库中的一致。在HSP70 的 5’ 端有起始码,抗原肽基因序列的 3’端有终止码,序列分析的结果与设计相符。
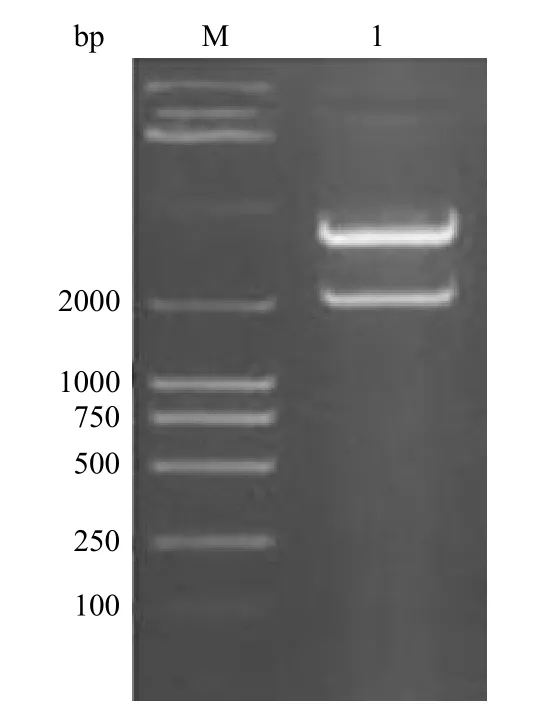
图2 pET-28a(+)/QLVFGIEVV-HSP70 重组质粒的酶切鉴定Figure 2 Identification of recombinantpET-28a(+)/QLVFGIEVV-HSP70
2.3 大肠杆菌的融合蛋白表达及纯化
SDS-PAGE 电泳显示经 IPTG 诱导后,重组载体 PET-28a(+)/QLVFGIEVV-HSP70 在约 71 kD处有一条高效表达的蛋白条带,与预期大小一致,未经诱导的重组载体未见此现象。经 Ni Sepharose 6FF 亲和层析后,在分子量约为71 kD 处有 1 条纯化的蛋白带(图3)。经 BandScan 软件分析,目的蛋白纯度为93.2%。
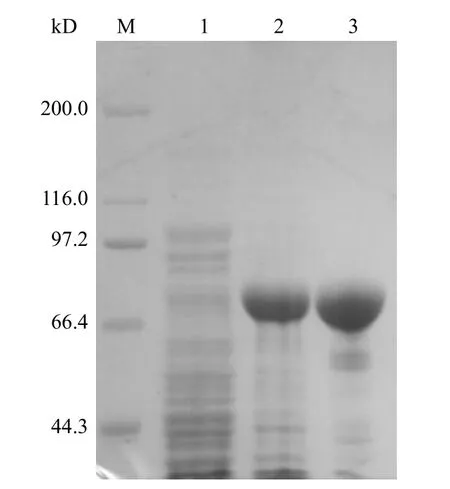
图3 原核表达 QLVFGIEVV-HSP70 融合蛋白 SDSPAGE 电泳分析Figure 3 Analysis of prokaryotic expression QLVFGIEVVHSP70 by SDS-PAGE
3 讨论
肿瘤抗原的研究不仅对阐明免疫应答机制的肿瘤免疫逃避机制具有十分重要的理论意义,而且对肿瘤的发病学、诊断、治疗以及疫苗制备都具有重要的应用意义,是当今国际肿瘤免疫学研究的一个热点,也是一个难点。目前,国内外已经对黑色素瘤、前列腺癌、脑胶质瘤、乳腺癌等多种肿瘤的肽疫苗进行了临床试验并取得了一定效果。肿瘤多肽疫苗虽具备诸多的优势但也因存在单独免疫时免疫原性较弱、体内易降解、只能激发低水平的CTL 反应等问题而难以获得理想的抗肿瘤效果。
有实验证实,MAGE 蛋白及其 CTL 表位不仅能在体外有效诱导特异性细胞毒性 T 淋巴细胞的产生,而且也能在体内诱导特异性 CTLs 的产生,并使部分患者的肿瘤有所缩小或完全消退[4]。目前,已有数十个 CTL 表位被鉴定,且这些表位均可在体内、外激发特异性 CTL 抗瘤免疫应答,部分多肽表位已进入临床 I 期和 II 期实验,并取得较好的疗效。其中本课题组筛选鉴定出的 MAGE-n 第159~167 位(QLVFGIEVV)为HLA-A2 分子递呈,并可激活 CD8+ 和 CD4+ T 细胞[2]。后续的工作中我们将 MAGE-n 与 MAGE-3 的 HLA-A2限制性表位联合应用进行研究,证实了联合表位肽较单个表位肽具有更强的免疫效应[5]。CTL 表位肽疫苗抗肿瘤的机制在于用 CTL 表位肽刺激机体后,可引起抗原特异性的 CTL 细胞的克隆增殖,而 CD8+ 的 CTL 是目前认为抗肿瘤免疫的主要效应细胞[6],其向胞外释放穿孔素、细胞溶解素等物质或通过细胞表面的死亡受体等杀灭靶细胞,同时亦可分泌 IFN-γ 等细胞因子间接杀伤肿瘤细胞,为杀伤肿瘤创造一个良好的微环境。
虽然 MAGE 为肿瘤的免疫治疗提供了良好的肿瘤特异性抗原,但是对抗原的有效加工提呈也是肿瘤免疫治疗中的一个关键问题。HSP70 具有类似 MHC 结合抗原肽的结构域,可作为抗原递呈分子直接将抗原肽递呈至细胞表面激发特异的 γδ T 细胞反应。更为重要的是,HSP 本身无免疫原性,但所结合的多肽具有免疫原性,HSP 不具有多态性,HSP-肽复合物在同种内的相互作用是可行的[7]。有研究者将 HSP 肽-复合物用于人的肿瘤治疗,I 期临床试验结果表明没有明显的自身免疫效应和细胞毒性,可使 50%患者的 CD8+ T 细胞增加,部分患者还有 NK 细胞扩增趋势,显示出了临床应用前景[3]。我们以往的研究结果也证实HSP70 能够增强 MAGE3/HSP70 蛋白疫苗的CTL 效应[8]。相比其他免疫佐剂而言,将 HSP70与肿瘤肽复合物联合制成疫苗能够激发抗原特异的 CTL 反应,具有更广阔的临床应用前景。本研究中我们采用 PCR 方法将 MAGE-n159-167(QLVFGIEVV)的 cDNA 序列简便、快捷地融合到 HSP70 的 3’ 末端,经酶切鉴定及测序证实,融合基因构建成功,抗原肽 QLVFGIEVV 与HSP70 基因的开放读框正确,并在大肠杆菌中表达出与预期 Mr 大小一致的蛋白,该表达产物为肽疫苗的研制提供了实验基础。
[1]Wu W, Sui YF, Ye J, et al.The cloning of tumor-associated gene MAGE in human hepatocellular cell line.Chin J Cell Mol Immunol,2002, 18(3):270-273.(in Chinese)武文, 隋延仿, 叶菁, 等.人肝癌细胞系中肿瘤相关基因MAGE的克隆.细胞与分子免疫学杂志, 2002, 18(3):270-273.
[2]Dong HL, Sui YF, Li ZS, et al.Efficient induction of cytotoxic T lymphocytes specific to hepatocellular carcinoma using HLA-A2-restricted MAGE-n peptide in vitro.Cancer Lett, 2004,211(2):219-215.
[3]Blachere NE, Li Z, Chandawarkar RY, et al.Heat shock protein-peptide complexes, reconstituted in vitro, elicit peptide specific cytotoxic T lymphocyte response and tumor immunity.J Exp Med, 1997, 186(8):1315-1322.
[4]Marchand M, van Baren N, Weynants P, et al.Tumor regressions observed in patients with metastatic melanoma treated with an antigenic peptide encoded by gene MAGE-3 and presented HLA-A1.Int J Cancer, 1999, 80(2):219-230.
[5]Zhang XM, Zhang YF, Huang Y, et al.The anti-tumor immune response induced by a combination of MAGE-3/MAGE-n-derived peptides.Oncol Rep, 2008, 20(1):245-252.
[6]Schmidt-Weber CB, Blaser K.Immunological mechanisms in specific immunotherapy.Springer Semin Immunopathol, 2004, 25(3/4):377-390.
[7]Srivastava PK.Immunotherapy for human cancer using heat shock protein-peptide complexes.Curr Oncol Rep, 2005, 7(2):104-108.
[8]Ma JH, Sui YF, Ye J, et al.Heat shock protein 70/MAGE-3 fusion protein vaccine can enhance cellular and humoral immune responses to MAGE-3 in vivo.Cancer Immunol Immunother, 2005, 54:907-914.
猜你喜欢
生命科学研究(2021年4期)2021-09-08
江西农业学报(2021年4期)2021-04-20
智慧健康(2021年2期)2021-03-15
昆明医科大学学报(2021年1期)2021-02-07
水生生物学报(2021年1期)2021-02-04
三农资讯半月报(2020年11期)2020-06-21
温州医科大学学报(2019年4期)2019-04-28
中国免疫学杂志(2017年1期)2017-01-17
肿瘤影像学(2015年3期)2015-12-09
中国当代医药(2015年9期)2015-03-01
