
广西青梅幼苗与成年树木光合特性和叶片显微结构的比较研究
2025-02-16 00:00:00潘鑫峰邹蓉唐健民韦霄江海都杨一山
广西植物 2025年1期


摘 要: "广西青梅为国家一级保护野生植物和极小种群野生植物。该文利用Li-6400便携式光合测定系统分析仪和真空电子扫描镜对广西青梅幼苗和成年树木光合生理特性与结构特征进行比较分析,探究其不同生长发育阶段叶片的光合能力和叶片结构之间的差异。结果表明:(1)直角双曲线修正模型是广西青梅光响应拟合的最佳模型。(2)成年树木叶片的最大净光合速率(Pn max)、光饱和点(LSP)和暗呼吸速率(Rd)均显著高于幼苗,而光补偿点(LCP)低于幼苗(Plt;0.05)。(3)幼苗的叶绿素a、叶绿素b、叶绿素(a+b)和类胡萝卜素均显著低于成年树木(Plt;0.05)。(4)成年树木叶片的气孔器长轴、气孔面积和气孔密度显著高于幼苗;成年树木叶片的叶面积和比叶重(SLW)显著(Plt;0.05)大于幼苗,而比叶面积(SLA)小于幼苗。综上认为,广西青梅具有幼苗喜阴而成年树木喜阳的光合特性。幼苗的光适应能力、光能利用率较弱以及成年树木过度遮阴导致幼苗无法进行充分光合以满足生长需求,是幼苗无法成长为大树的重要原因。在广西就地保护中可通过人为的方式,对乔木层林木进行人工“开窗”,提供相对充足的光源,以利于广西青梅幼苗生长发育成大树及种群更新;在引种栽培中,广西青梅幼苗时期需要适当的遮阴处理,以及移栽至有一定荫蔽度的环境中。
关键词: 广西青梅, 拟合模型, 光合特性, 叶片显微结构, 叶绿素
中图分类号: "Q945
文献标识码: "A
文章编号: "1000-3142(2025)01-0133-14
基金项目: "国家重点研发计划课题(2022YFF1300703); 国家自然科学基金(32160091,32060248); 中国科学院“西部之光”计划(2022); 广西林业科技推广示范项目(2023LYKJ0 [2022] GT23); 广西植物功能物质与资源持续利用重点实验室自主项目(ZRJJ2022-2); “桂科学者”创新团队启动项目(CQI-C-192)。
第一作者: 潘鑫峰 (1997—),硕士,研究方向为保护生物学研究,(E-mail)panxinfeng@fafu.edu.cn。
*通信作者: "邹蓉,副研究员,研究方向为药用植物学和保护生物学研究,(E-mail)175183030@qq.com。
Comparative study on photosynthetic characteristics
and leaf microstructure of Vatica guangxiensis
seedling and adult tree
PAN Xinfeng, ZOU Rong*, TANG Jianmin, WEI Xiao, JIANG Haidu, YANG Yishan
( Guangxi Institute of Botany, Guangxi Zhuang Autonomous Region and Chinese Academy of Sciences, Guilin 541006, Guangxi, China )
Abstract: "Vatica guangxiensis is a national first-class protected wild plant and the wild plant of extremely small populations. This paper provides a comparative analysis of the photosynthetic physiological characteristics and structural features of V. guangxiensis seedling and adult tree. The Li-6400 portable photosynthesis measurement system analyzer and vacuum electron scanning microscope were utilized to investigate the differences between the photosynthetic capacity and leaf structure of leaves at different stages of growth and development. The results were as follows: (1) The right-angled hyperbolic modified model was the best model to fit the light response of V. guangxiensis. (2) The maximum net photosynthetic rate (Pn max), light saturation point (LSP) and dark respiration rate (Rd) of adult leaves were significantly higher than those of seedling, while the light compensation point (LCP) of adult tree was lower than that of seedling (Plt;0.05). (3) Chlorophyll a, chlorophyll b, chlorophyll (a+b) and carotenoids in seedlings were significantly lower than those in adult tree (Plt;0.05). (4) The stomatal long axis, stomatal "area and stomatal density of adult tree were significantly higher than those of seedling; the leaf area and specific leaf weight (SLW) of adult tree were significantly larger than those of seedling (Plt;0.05), and the specific leaf area (SLA) was smaller than that of seedling. In summary, V. guangxiensis exhibits photosynthetic characteristics of a shade-loving seedling and a sun-loving adult tree. The seedlings have weak light adaptation ability and light energy utilization, and are unable to photosynthesize sufficiently to meet their growth needs due to excessive shading by adult tree. This is a significant reason why seedlings are unable to grow into large trees. Therefore, for in situ conservation of V. guangxiensis, human interference can be used to artificially create ‘windows’ in the tree layer to provide sufficient light sources. This is conducive to the growth of V. guangxiensis seedlings into large trees and the renewal of the population. On the other hand, for the introductory cultivation of V. guangxiensis, the seedlings need to be appropriately shaded and transplanted to a certain degree of shade. During the initial cultivation process, it is important to provide appropriate shading for seedlings and transplant them to an environment with a suitable level of shade.
Key words: Vatica guangxiensis, fitting model, photosynthetic characteristics, leaf microstructure, chlorophyll
广西青梅(Vatica guangxiensis)为落叶乔木,隶属于龙脑香科青梅属,是我国一级保护野生植物(肖云学等,2023)。其树干通直圆满,材色美观,木材坚硬,纹理直,结构细致,耐腐性强,是我国特有的热带珍贵用材树种,在建筑、桥梁、造船、车厢以及制造各种高级家具中作为优良用材(彭玉华等,2015)。广西青梅具有地域分布狭窄,种群退化和数量稀少等极小种群的特征。极小种群野生植物是重要的生物遗传多样性资源,若不及时保护可能导致它们的生物特征和遗传多样性资源的消失,因此保护极小种群植物对于我国的生物多样性保护具有至关重要的意义(张则瑾等,2018)。广西青梅主要分布于广西和云南,生于海拔 500~600 m 的北热带沟谷雨林中的丘陵地带,广西境内仅在那坡县发现1个野生种群。广西青梅作为青梅属在广西分布的唯一种类,对研究我国热带植物区系有十分重要的意义。在那坡县生境中,广西青梅成年树木生长良好,然而幼苗自然更新进展困难,无法正常生长为成树,种群繁衍与扩展受到严重桎梏。为探究其原因并对广西青梅进行合理的保育措施,研究人员对广西青梅的群落结构(蒋迎红等,2016)、土壤理化性质(谭长强等,2016)、遗传多样性(Zhu et al., 2023)等方面进行了相关研究,但并未从光合特性和叶片显微结构的层面进行研究和报道。
光合作用是植物生长和发育的基础,可根据其特性判断和确定植物最适宜的环境条件和生态需求(Adamec, 1997)。光合特性并非在植物的整个生命周期中都保持不变,而是随着树龄的变化而动态变化(Bond, 2000)。研究濒危植物不同时期的光合特性差异,为评估和选择适宜的栖息地提供重要参考,也为濒危植物的保育与精细化栽培奠定基础。对于木本植物,幼苗和成年植株的光合特性差异较为显著。相关研究发现从夹竹桃的幼苗与成年植株在光合特性和光保护特性存在巨大差异,导致在应对不同季节时呈现出不同生理特性(Chondrogiannis et al., 2023);在热带雨林中,对17个物种的幼树与成年植株的叶片特性进行了对比分析,发现在所有物种的幼苗与成年植株之间的光合特性与叶片结构均存在显著差异,而这些差异与其环境适应性以及生长状态存在相关性(Houter amp; Pons, 2012);另外,叶片显微结构是观察植物差异性重要方法,而表皮细胞厚度、栅栏组织和海绵组织等叶片显微结构的变化会影响植物的光合作用调节(许爱祝等,2024)。因此,了解和分析单一植物不同生长发育阶段的不同叶片解剖结构以及对光合特性的不同影响,这也是保护濒危植物面临的巨大挑战(罗鸣等,2019)。在香木莲的幼苗与成年株中存在较大的光合特性差异,该差异主要由海绵组织厚度、叶片厚度以及叶绿素含量所造成(潘李泼等,2023);在新西兰圣诞树叶片显微结构的研究中,成熟叶片植株中较厚的栅栏组织和海绵组织有助于减少光通过叶片的穿透,从而防止光诱导的损伤,但也导致光合速率低于幼叶(Kubien et al., 2007)。从上述研究可以发现,研究同种植物不同时期的光合特性和生理结构之间存在的差异,对于阐明其濒危机制及开展合适的保育措施具有重要意义。本研究推测广西青梅种群更新困难是成年树木与幼苗的光合特性差异所造成。为验证此推测,对广西青梅幼苗和成年树木的光合特性、不同色素含量和叶片结构进行测定和分析,拟解决以下几个问题:(1)广西青梅成年树木与幼苗的光合特性是否存在差异;(2)若光合能力存在差异,该差异是否与叶片结构和色素含量有关。本研究结果旨在了解广西青梅不同时期的光合特性和差异,揭示广西青梅叶片在不同生长发育时期的光响应机制,为广西青梅的迁地保护和种群恢复提供理论依据并探索如何对广西青梅幼苗进行保护性种植。
1 材料与方法
1.1 研究区概况
研究区域位于百色市那坡县百合乡平坛村的那芝山上(105°49′47″ E、23°9′36″ N)。研究区域属于亚热带季风气候,年均温度18.8 ℃,年均降水量 1 408.3 mm 。广西青梅的群落为常绿阔叶林,树木高大繁茂,郁闭度0.8以上,土壤为砂页岩发育而成的黄红壤,土层疏松深厚,林下枯枝落叶层厚,腐殖质丰富。主要伴生树种有乌榄(Canarium pimela)、苹婆(Sterculia nobilis)、香楠(Aidia canthioides)、柄果木(Mischocarpus sundaicus)等。
1.2 材料
分别选定同一生境下的生长健康、无病虫害的广西青梅幼苗(1年生幼苗)与10年生成年树木各3株,在成年树木与幼树的树冠中部向阳叶片中各选取3片生长良好的成熟叶片进行野外原位光合参数测定。叶绿素含量测定和叶片解剖结构的实验同样选择树冠中部向阳的健康成熟叶片。
1.3 方法
1.3.1 光合-光响应曲线的测定
利用Li-6400便携式光合测定系统分析仪测定叶片的光响应曲线。由于幼苗和成年树生长的光环境差别太大,测定光响应曲线时幼苗采用800 μmol·m-2·s-1,成年树木幼苗用1 200 μmol·m-2·s-1光强下诱导30 min(仪器自带的红蓝光源)以充分活化光合系统。使用开放气路,空气流速为0.5 L·min-1,叶片温度为28 ℃,CO2浓度为400 μmol。设定的光强梯度为1 800、1 500、1 200、1 000、800、600、400、200、150、100、50、20、0 μmol·m-2·s-1,测定时每一光强下停留120~200 s。测定3种光合参数,即净光合速率(Pn)、蒸腾速率(Tr)和气孔导度"(Gs),利用公式计算水分利用率(water use efficiency,WUE):WUE=Pn/Tr。
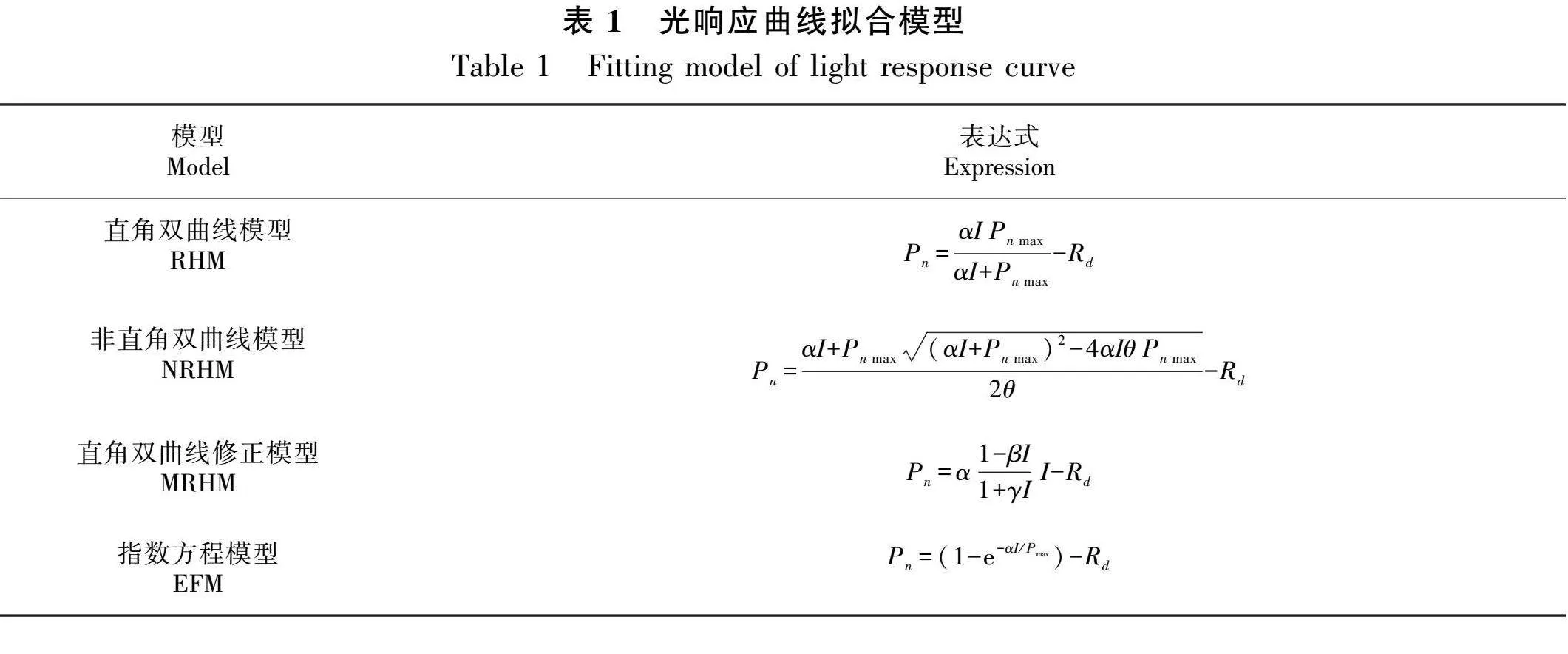
1.3.2 光响应曲线模型 直角双曲线模型(rectangular hyperbola model,RHM)、非直角双曲线模型(non-rectangular hyperbola model,NRHM)、直角双曲线修正模型(modified rectangular hyperbola model,MRHM)和指数方程模型(exponential function model,EFM)为常见的光响应曲线拟合模型(Ye et al., 2012)。利用这4种模型对叶片实测值进行拟合,模型表达式见表1。根据相关文献(Ye et al., 2012; 叶子飘等,2016)分别计算4种模型拟合光饱和点(light saturation point,LSP)、光补偿点(light compensation point,LCP)、表观量子效率(apparent quantum yield,AQY)和暗呼吸速率(Rd)。
1.3.3 叶绿素含量的测定 分别选定3株幼苗与成年树木,每株各选取3片生长良好的叶片。清理干净叶片后准确称量0.2 g,参考Lichtenthaler和Buschmann(1987)的方法,用紫外可见分光光度计Alpha 1502(上海谱元仪器有限公司)在470、649、665 nm波长下测定吸光值,计算出叶绿素a(Chla)、叶绿素b(Chlb)、类胡萝卜素(Car)的含量,以及叶绿素a与叶绿素b的比值(Chla/Chlb),叶绿素与类胡萝卜素的比值(Chl/Car)。
1.3.4 叶片解剖结构 将新鲜叶片沿着横截面精准地切成4小块,随后即刻放入2.5%的戊二醛固定液中进行固定。固定后的叶片采用乙醇梯度脱水技术,分别经过30%、50%、70%、85%、90%的乙醇脱水处理各1次,以及100%乙醇处理2次,每次处理时长15 min。脱水完成后,进行临界点干燥和镀金处理,在真空电子扫描镜(ZEISS EVO18)下观察和拍摄。使用Axio Vision SE64 Rel.4.9.1扫描电镜软件,对叶片结构的各项指标进行精确测定,包括叶片厚度(leaf thickness,LT)、上表皮厚度(upper epidermal thickness,UET)、下表皮厚度(lower epidermal thickness,LET)、栅栏组织厚度(palisade thickness,PT)、海绵组织厚度(spongy thickness,ST)以及气孔面积(stomata area,SA)。在计算气孔密度时,采用视野气孔个数除以视野面积的公式进行计算。将每个叶片剪成小块测定叶片微形态指标,随机重复测定5次以上。因此,每个叶片微形态指标,共有150个数据(1个生境 × 2个树种 × 3株树 × 5片成熟叶片 × 5 次重复=150个数据)
1.3.5 叶片性状参数测定 分别从幼苗与成年树木中各选取50片叶片,先用Li-3000叶面积仪测定其叶面积,再在110 ℃下处理30 min,80 ℃烘干24 h后,用电子天平称其干重,利用叶片干重除以叶面积(leaf area,LA)获得比叶重(specific leaf weight,SLW),比叶面积(specific leaf area,SLA)为SLW的倒数。
1.4 数据处理与分析
利用叶子飘等(2016)方法中的RHM、NRHM、MRHM和EFM进行光合参数的计算。其中,MRHM通过软件直接计算LSP,而RHM和NRHM需要通过公式Pn max = AQY×LSP - Rd来计算LSP,式中AQY 为光强小于200 μmol·m-2·s-1拟合直线方程的斜率。运用Excel软件、SPSS 23.0软件进行数据处理和分析,并运用独立样本t检验进行显著性分析,随后使用Origin 2023b软件作图。
2 结果与分析
2.1 光响应模型对光响应曲线的拟合效果
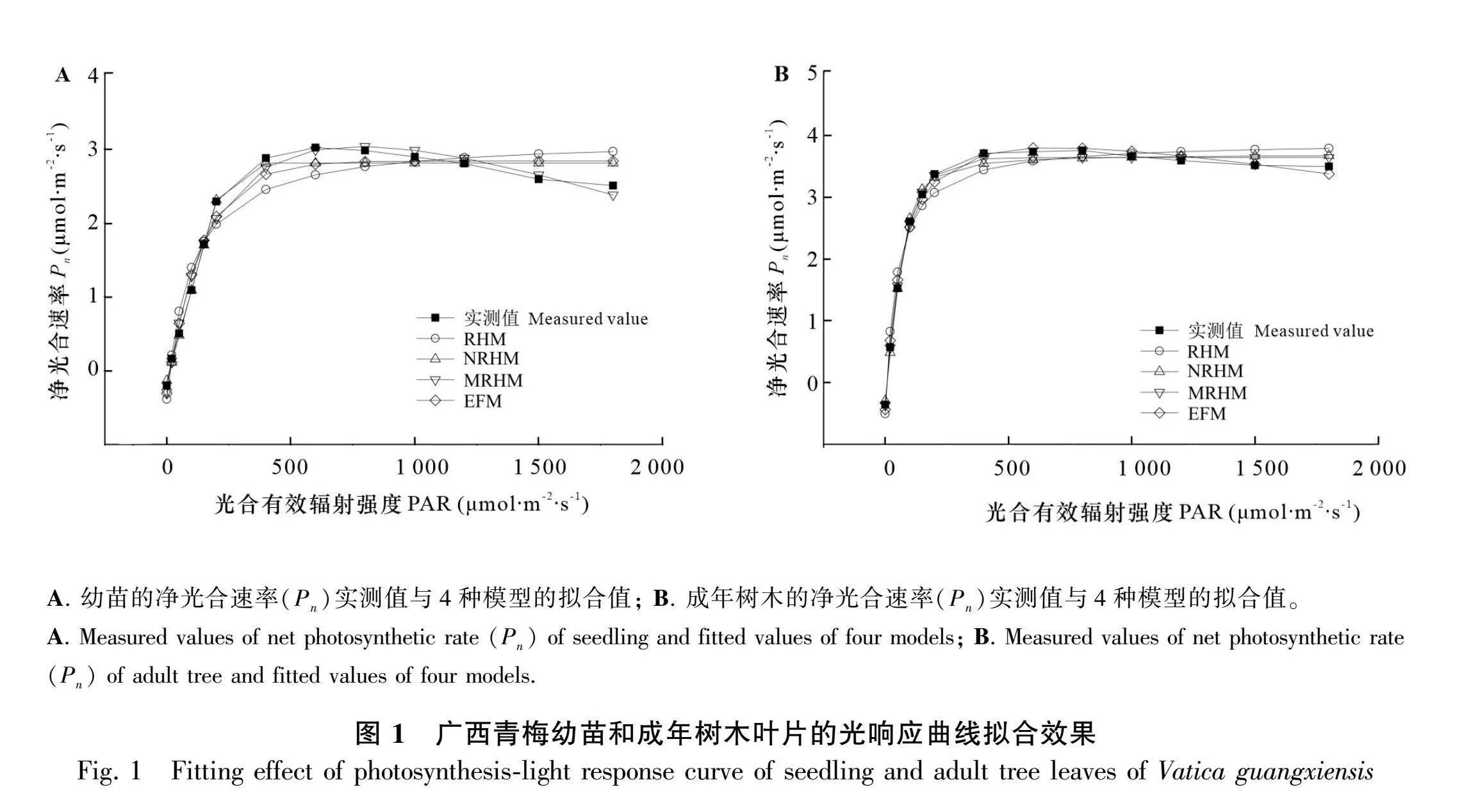
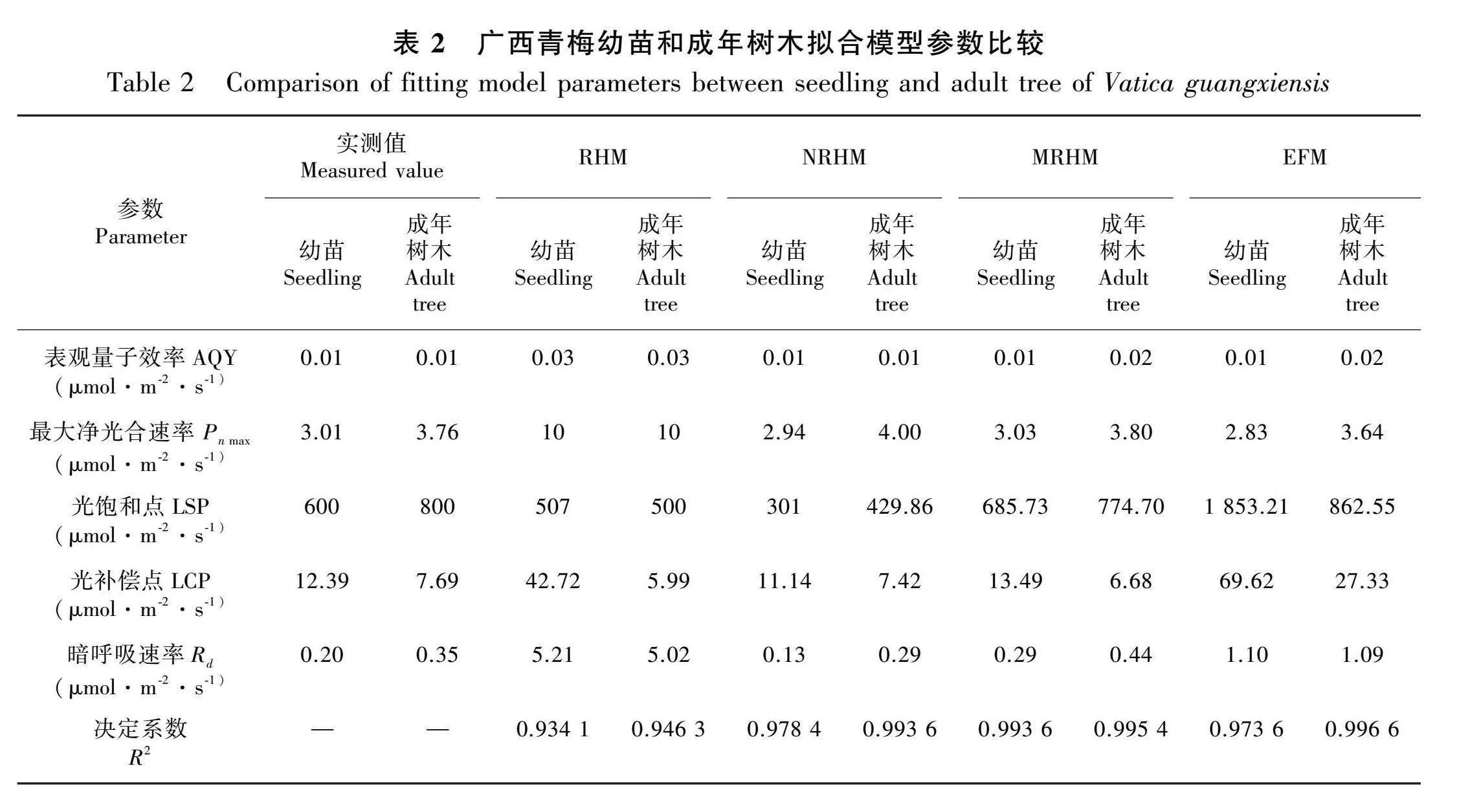
将广西青梅成年树木和幼苗的净光合速率(Pn)实测值与4种模型的拟合值作图,由图1:A可知,MRHM的拟合值与幼苗的实测值最为接近。当光合有效辐射(photosynthetically active ratiation,PAR)为0~200 μmol·m-2·s-1时,Pn呈线性增长趋势;当PAR达到200~800 μmol·m-2·s-1时,Pn的增长速率放缓;当PAR大于800 μmol·m-2·s-1时,Pn停止增长,并有缓慢下降的趋势。由图1:B可知,成年树木与NRHM、MRHM和EFM的拟合值都较为接近。当PAR为0~200 μmol·m-2·s-1时,Pn呈线性增长趋势;当PAR为400~600 μmol·m-2·s-1时,Pn的增长速率放缓;当PAR大于600 μmol·m-2·s-1时,Pn停止增长,并有缓慢下降的趋势。由表2可知,在幼苗的模型拟合中除RHM的决定系数(R2)较低外(0.934 1),其余3种模型均较高,其中MRHM的R2最高(0.993 6),NRHM次之(0.978 4),EFM排第三(0.973 6)。成年树木的拟合结果与幼苗相似,RHM的R2最低(0.946 3),其余3种模型的R2高于0.99,与幼苗拟合不同的是,EFM的R2最高(0.996 6),MRHM次之(0.995 4),NRHM排在第三(0.993 6)。幼苗的NRHM与EFM在PAR范围内无法拟合出Pn的极值,而MRHM能够在PAR为600 μmol·m-2·s-1强度下拟合出极值。
2.2 幼苗和成年树木拟合模型参数比较
对4种模型计算出的幼苗和成年树木的光合参数和实测值进行比较。由表2可知,幼苗和成年树木实测值的表观量子效率(AQY)均为0.01 μmol·m-2·s-1,最大净光合速率(Pn max)分别为3.01 μmol·m-2·s-1、3.76 μmol·m-2·s-1,LSP分别为 600 "μmol·m-2·s-1、800 μmol·m-2·s-1,LCP分别为 12.39 μmol·m-2·s-1、7.69 μmol·m-2·s-1。从Pn max、AQY、LCP、LSP这 4个参数看,幼苗和成年树木的光响应曲线拟合效果最差的是RHM,拟合效果较好的是MRHM和EFM,其次为RHM;根据LCP和LSP可知,幼苗光照度适应范围(12.39 ~600 μmol·m-2·s-1)"低于成年树木(7.69~800 μmol·m-2·s-1)。
2.3 幼苗和成年树木光合-光响应曲线
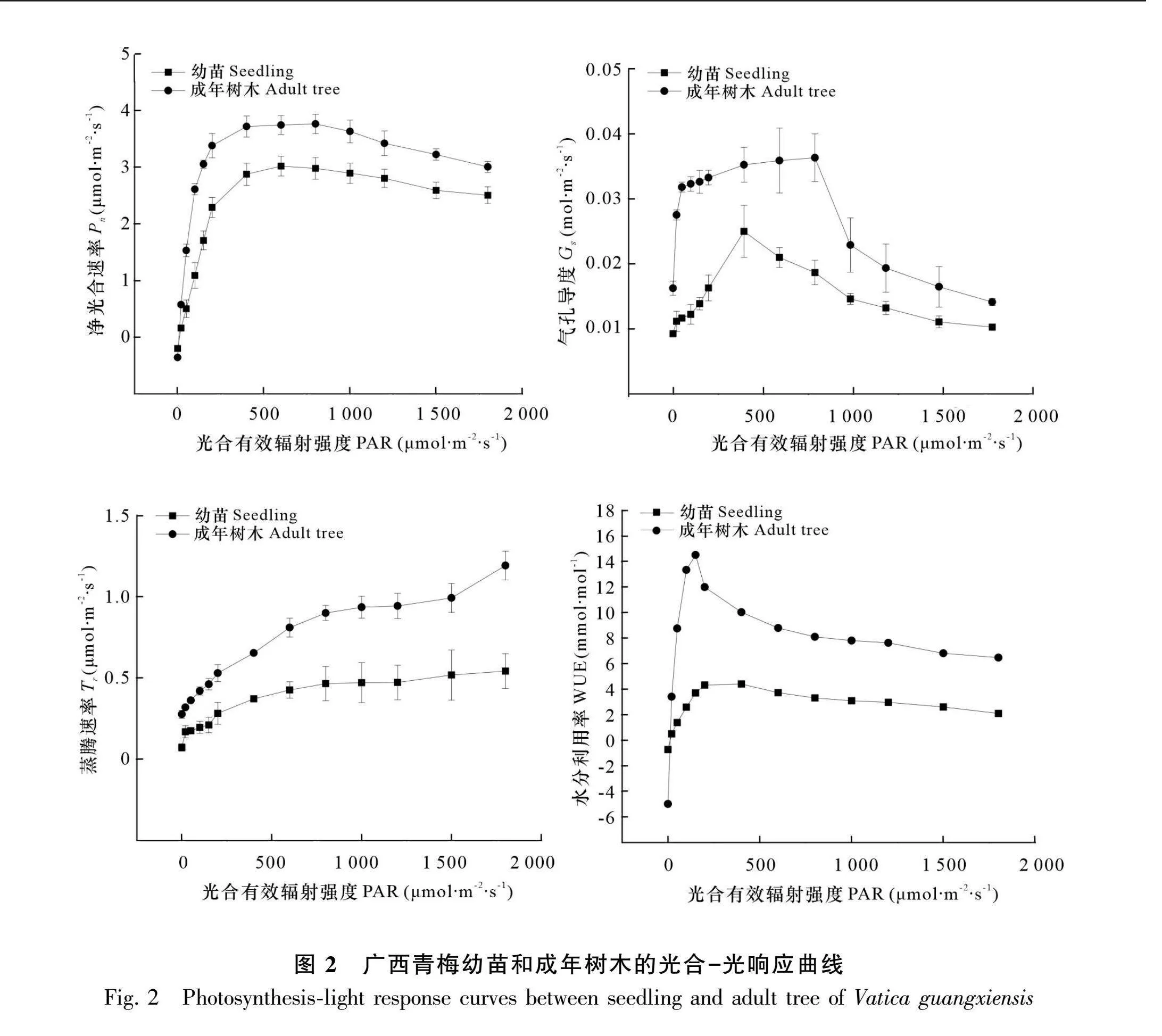
由图2可知,在PAR为0~1 800 μmol·m-2·s-1范围内成年树木的Pn、Tr、Gs和WUE均高于幼苗。幼苗和成年树木的Pn均随PAR先升高后下降,当PAR为0~200 μmol·m-2·s-1时,幼苗和成年树木的Pn均快速上升。幼苗和成年树木的Pn在PAR为400~800 μmol·m-2·s-1范围内时逐渐稳定,而当PAR大于800 μmol·m-2·s-1以后,幼苗和成年树木的Pn呈下降趋势。幼苗和成年树木的Gs均随PAR升高呈先升高后下降的趋势,在PAR为0~500 μmol·m-2·s-1时,幼苗和成年树木Gs均呈上升趋势;当PAR超过400 μmol·m-2·s-1时,幼苗Gs呈下降趋势,而成年树木持续到800 μmol·m-2·s-1后才出现下降趋势。幼苗和成年树木的Tr随PAR升高呈持续上升趋势。幼苗和成年树木的WUE随PAR升高而上升,在PAR为250 μmol·m-2·s-1"时均达到极值,随后随PAR上升而下降。
2.4 幼苗和成年树木的光合色素比较
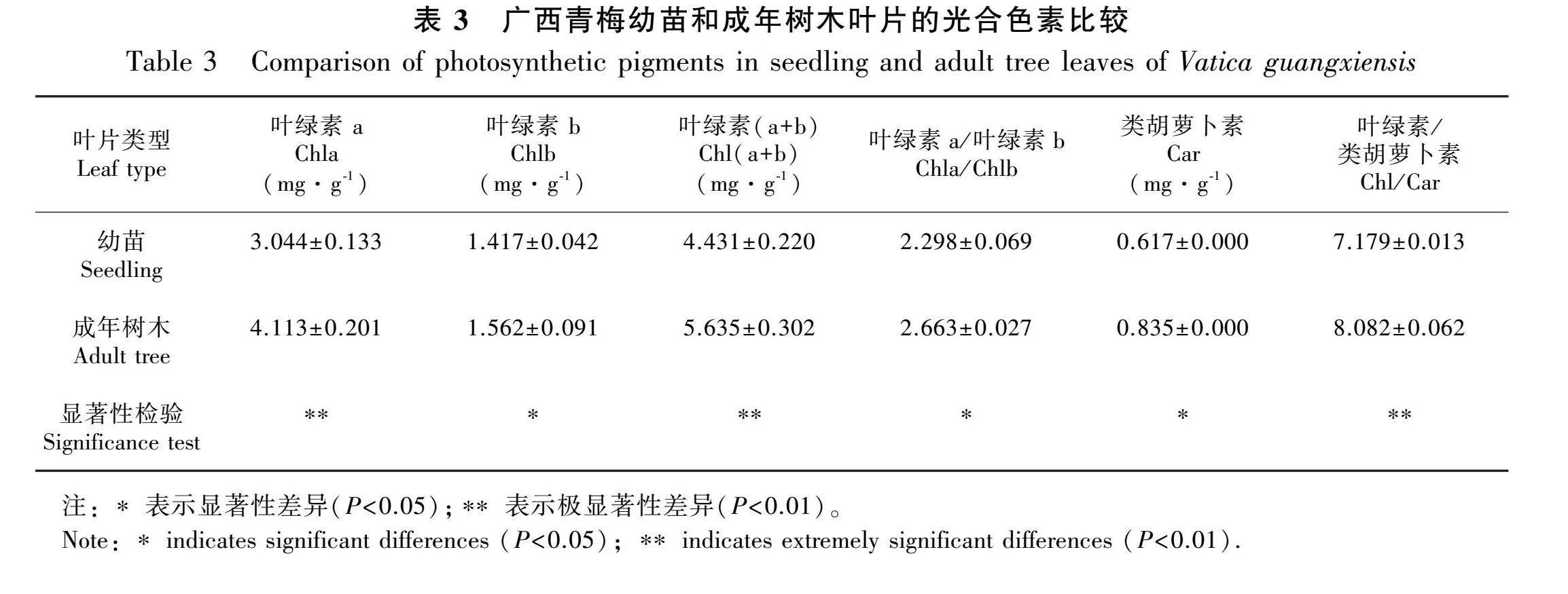
幼苗和成年树木的光合色素含量如表3所示:幼苗Chla含量为(3.044±0.133) mg·g-1"极显著低于成年树木(4.113±0.201) mg·g-1(Plt;0.05);幼苗Chlb含量(1.417±0.042)显著低于成年树木(1.562±0.091 mg·g-1)(Plt;0.05);幼苗Car含量(0.617±0.000)"mg·g-1显著低于成年树木(0.835±0.000) mg·g-1)(Plt;0.05);幼苗Chla/Chlb(2.298±0.069)显著低于成年树木(2.663±0.027)(Plt;0.05)。
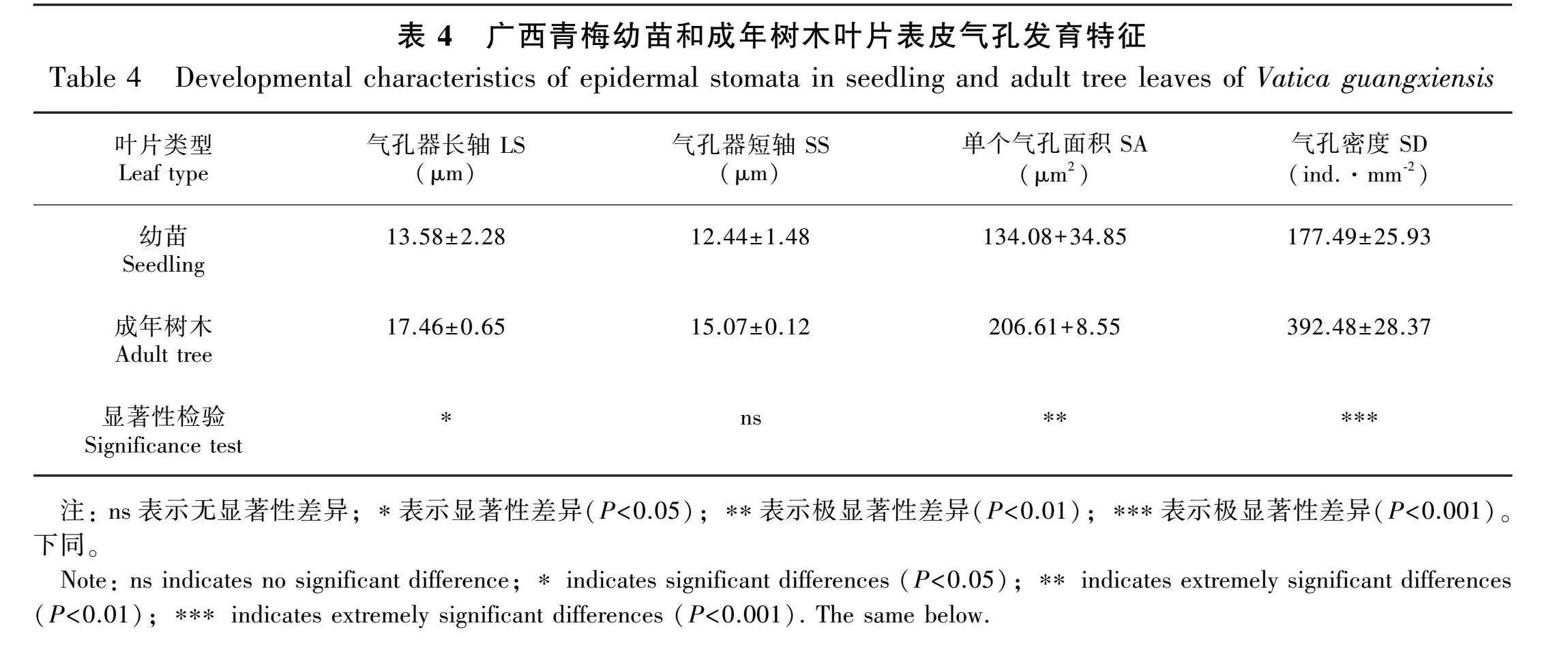
2.5 成年树木和幼苗的气孔发育比较
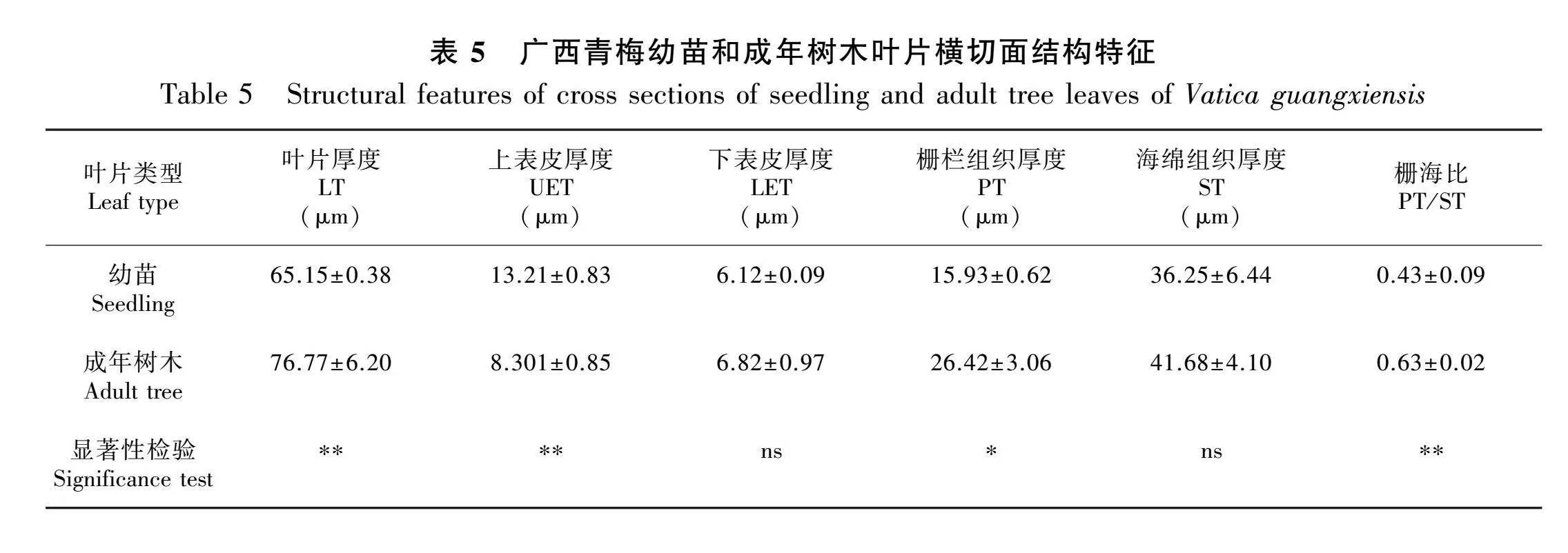
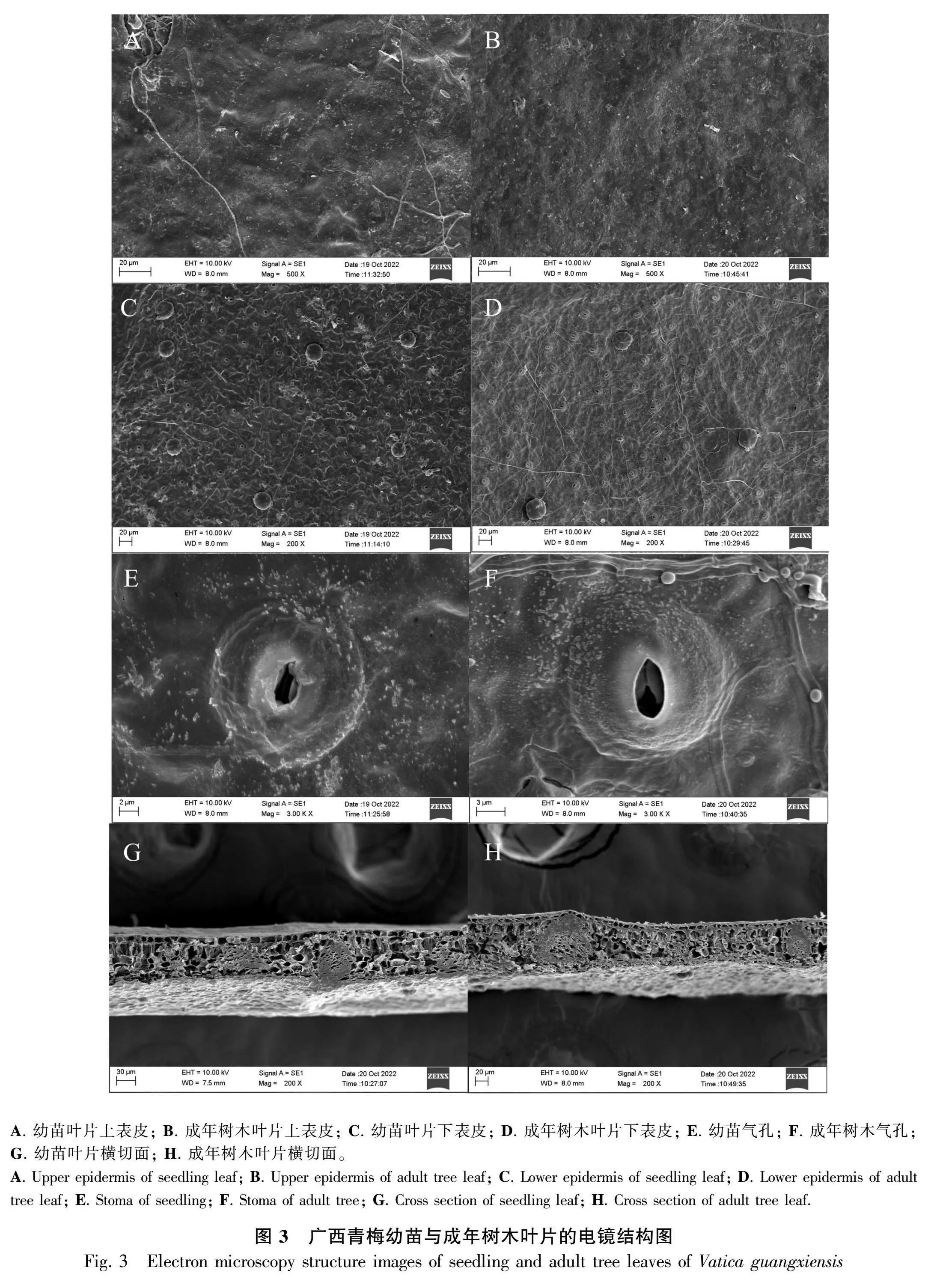
如图3:A、B所示,成年树木的上表皮褶皱明显多于幼苗且两者的上表皮均未发现气孔的分布。如图3:C、D所示,成年树木和幼苗的下表皮存在大量的气孔组织。成年树木的气孔密度(392.48±28.37)ind.·mm-2极显著高于幼苗(177.49±25.93) ind.·mm-2(Plt;0.001)(表4)。通过图3:C-F和表4的结合分析可知,除气孔器短轴(short axis of stomata,SS)无显著性差异外,成年树木的气孔器长轴(long axis of stomata,LS)、单个气孔面积(SA)、气孔密度(stomatal density,SD)等指标均显著高于幼苗(Plt;0.05)。无论是幼苗或成年树木都有明显结构分层,其结构由上至下为上表皮、栅栏组织、海绵组织和下表皮(图3:G、H)。 根据表5的横截面结构特征参数可知,成年树木的LT、PT以及PT/ST均显著高于幼苗(Plt;0.05或Plt;0.01),而幼苗的UET极显著高于成年树木(Plt;0.01)。
2.6 幼苗与成年树木的叶面积、叶比重和比叶面积比较
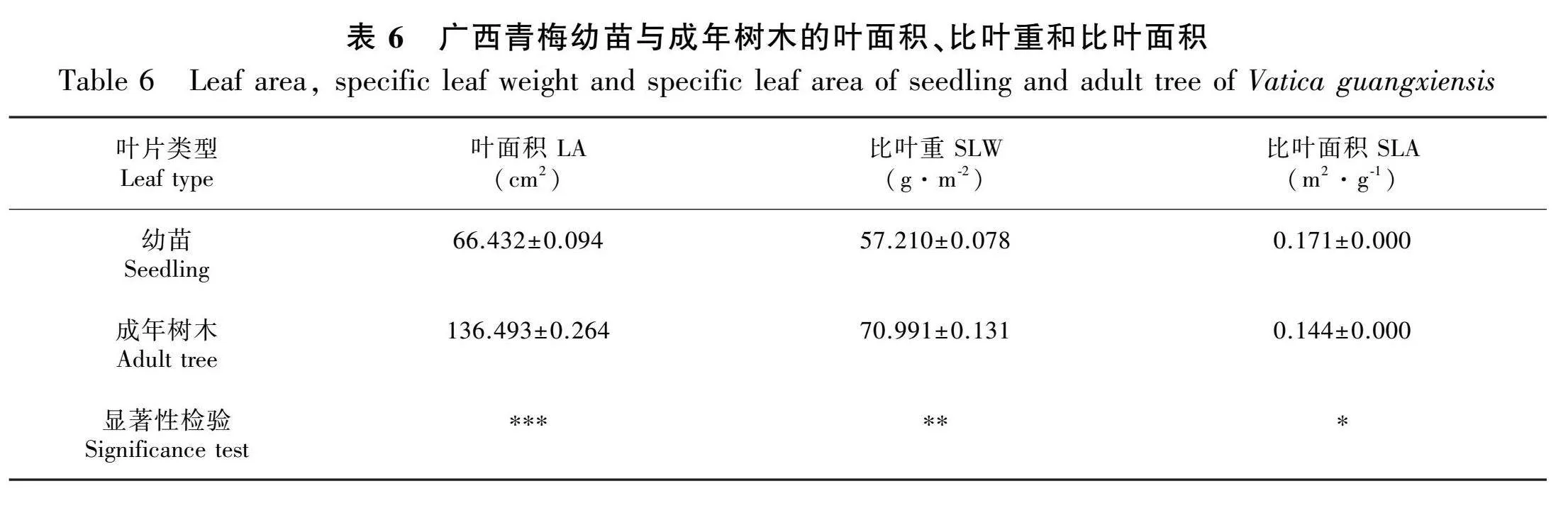
如表6所示,广西青梅幼苗的叶面积(LA)[(66.432±0.094)"cm2]极显著低于成年树木[(136.493±0.264) cm2](Plt;0.001),幼苗的比叶重(SLW)[(57.210±0.078) g·m-2]极显著低于成年树木[(70.991±0.131) g·m-2](Plt;0.01),幼苗的比叶面积(SLA)[(0.171±0.000) m2·g-1]显著高于成年树木[(0.144±0.000) m2·g-1](Plt;0.05)。
2.7 相关性分析
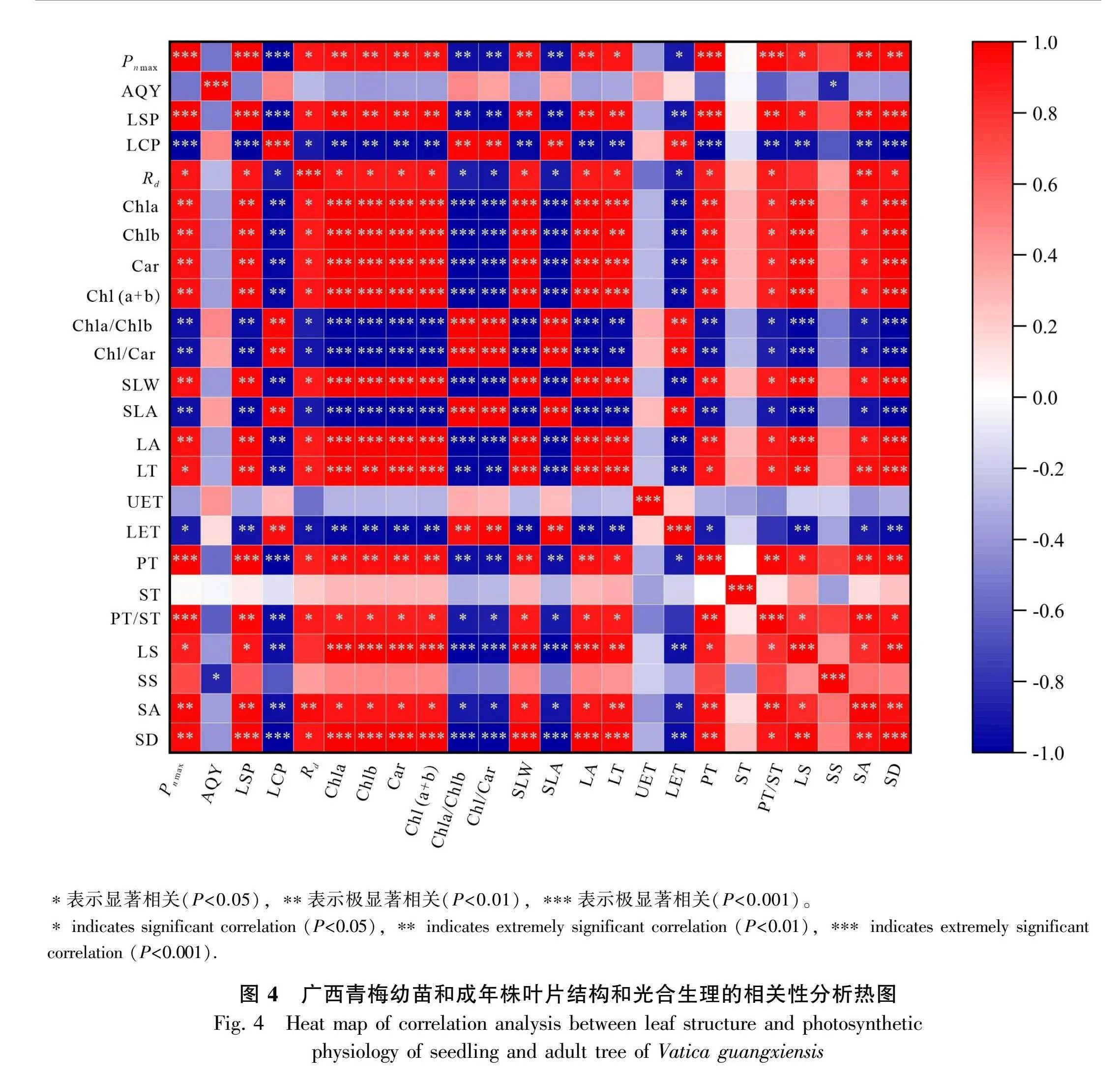
由广西青梅的幼苗和成年株叶片结构和光合生理的相关性分析热图(图4)可知,幼苗和成年树木叶片色素含量参数、结构参数和光合生理参数之间存在较强的相关性,其中叶片色素参数Chla、Chlb、Car、Chl(a+b) 与光合参数Pn max、LSP均存在显著正相关,Chla/Chlb、Chl/Car与Pn max存在显著负相关。叶片结构参数SLW、LA、LT、PT、PT/ST、LS、SA与光合参数Pn max、LSP存在显著正相关,LET、LA与Pn max存在显著负相关。
3 讨论
3.1 4种模型光响应曲线拟合
RHM、NRHM、MRHM和EFM作为在植物中常用的4种光响应曲线拟合模型适用于不同植物。其中,MRHM和EFM为大多数植物的最适模型。黑葡萄、枸杞、水稻等植物最适的光合响应模型是MRHM(闫小红等,2013),EFM常用于花生、藻类和光合能力较弱的水生植物中(刘瑞显等,2013)。本研究中,4种模型中仅有MRHM能较好地拟合出光抑制后的曲线部分,并且能够准确地计算出光补偿点。虽然EFM在幼苗的拟合中决定系数R2高于MRHM,但计算光饱和点时需要将光合速率假定为0.9或者0.99的最大净光合速率,结果的可信度较差,并且在幼苗中EFM模拟出较异常的光饱和点,在黄枝油杉中EFM也计算出异常的光饱和点(柴胜丰等,2015)。综上所述,MRHM是广西青梅叶片光响应拟合的最佳模型。
3.2 光合特性
成年树木的净光合速率、蒸腾速率、气孔导度和水分利用率都高于幼苗,在荔枝(张红娜等,2016)和辣木中同样观察到了此现象(王瑞苓等,2022),但在巴山榧中3年生树龄叶片的蒸腾速率和气孔导度高于15年生树龄叶片(高岚等,2018),在国槐中80年树龄的净光合速率、蒸腾速率和气孔导度3个指标均弱于20年树龄(程程等,2018)。从生理生态过程和适应机制推测成年树木通常具有较高的净光合速率,是由于成年树木叶面积更大、叶绿素相对较多并且光合器官更成熟(Kinose et al., 2019)。较大的叶片面积也使得成年树木的气孔数量高于幼苗,气孔是植物叶片表皮上的小孔,可控制进出叶片的气体交换,因此叶片面积与气孔可能是影响蒸腾速率和气孔导度的重要因素(Tuzet, 2014)。本研究的相关性分析也发现最大净光合速率与气孔密度和叶面积存在正相关性。光合特性差异是树木各种生理生态过程和适应机制之间复杂相互作用的一部分。全面了解这些差异还需要进一步对生理指标、叶片解剖结构及功能性状进行分析。
植物的光饱和点和光补偿点是评估植物光强适应性的重要指标。本研究发现, 成年树木和幼苗到达光饱和点后,净光合速率随光强的增加而减小,推测广西青梅的成年树木和幼苗在过高的光强下均出现光抑制效应。在蕨麻、杨树、白及等植物中也观察到同样的现象(罗鸣等,2019;李雪琴等,2022;陈兴浩等,2022)。成年树木的光饱和点高于幼苗,而光补偿点低于幼苗,说明成年树木的光环境适应性强于幼苗,这与濒危植物银缕梅研究结果相似(朱汤军等,2008)。推测成年树木的光补偿点低于幼苗的原因有以下三点:第一,可能是成年树木的叶片通常比幼苗的叶片更大且更厚,这意味着它们的光合作用系统更为发达,能够在较低的光照强度下进行有效的光合作用。第二,成树的叶片可能具有更高效的光能捕获和利用机制,如更多的叶绿体、更优化的叶绿体排列以及更有效的光合色素。此外,成树的叶片可能已经适应了其所在环境的光照条件,通过调整其生理和生化过程来降低光补偿点。第三,成树的叶片可能在气孔调节和水分利用效率方面也更为优化,进一步降低了光补偿点。在本研究的相关性分析中,叶片厚度、气孔数量、叶绿素含量与光补偿点呈显著负相关,但与净光合速率呈正相关,并进一步对这些原因进行论证。蒋迎红等(2016)和我们的野外调查发现,广西青梅种群中大量的幼龄个体能够存活在群落中其他优势乔木占据的主林层下,说明广西青梅种子发芽及幼苗的生长需要在荫蔽环境下完成,但这些幼苗要长成小树、中树较为困难。本研究结果表明广西青梅具有幼苗喜阴而成年树木喜阳的光合特性,幼苗难以长成中树或大树,可能由于其他乔木树种长期占据了上层,而广西青梅植株随年龄的增加对光的需求变化快,使得个体较难通过强烈的环境筛选进入种群的更替层。光适应的生态幅度较窄导致广西青梅种群扩大困难,是其濒危的重要原因之一。在广西就地保护中可通过人为干扰的方式,对乔木层林木进行人工“开窗”,提供相对充足的光源,以利于广西青梅大苗生长发育成大树和种群的更新。
3.3 叶片解剖结构及功能性状
比叶重的大小反映了叶片干物质的累积(刘明秀和梁国鲁,2016;刘琏等,2022 )。本研究中,广西青梅成年树木的比叶重显著大于幼苗。这可能与成年树木相对较强的光合作用以及广西青梅作为地区中的顶层优势植物从而能获得较多的光照辐射有关。广西青梅群落结构的相关研究结果侧面印证了此观点(蒋迎红等,2016)。本研究发现幼苗的比叶面积大于成年树木,推测幼苗可能受到了顶层大树遮光影响,林内的光合辐射强度大幅下降,为弥补光强的减少,幼苗以此进行适应性反应,这从侧面说明了广西青梅幼苗的耐阴性。在红果榆幼苗、蒙古栎和大叶女贞的光合研究中也获得相似的结果(李东胜等,2017;金雅琴和李冬林,2023;刘俊言等,2023)。本研究中比叶重与净光合速率呈正相关,而比叶面积与净光合速率呈负相关,说明二者与光环境的关联性,是植物对于不同环境条件作出的一种形态学反应,这种反应有利于植物在各种环境条件下更好地进行光合作用,从而保证其生长和生存。因此,引种栽培中广西青梅幼苗时期需要适当的遮阴处理,以及移栽至有一定荫蔽度的环境中。
叶绿素作为光合作用的关键基础, 其在植物细胞中的含量和比例是植物应对和利用环境的关键性指标(Rodríguez-calcerrada et al., 2008)。幼苗的叶绿素含量均低于成年树木,根据实际生长环境推测幼苗通常生长在成树的树荫下,光照条件较差。在这种环境下,叶绿素含量过高会导致光抑制,破坏光合作用。因此,幼苗可能会通过降低叶绿素含量来适应弱光环境。此结果与广西青梅光响应参数的结果相一致。在幼苗成长为成年树木的发育过程中,叶片逐步塑造出与生态环境相匹配的形态、结构和生理习性。叶片作为光合作用的核心器官,直接与外界环境接壤,对环境因子的敏感性极高(Meziane amp; Shiplep, 1999)。其中,叶片的叶面积与叶片发育阶段有着密切关系,气孔主要分布于下表皮,在幼苗阶段,为177 ind.·mm-2,随着叶片发育,植物为了更高效地进行水分和CO2的吸收和交换,气孔快速地形成和发育,到成年树木阶段,叶面积可达392 ind.·mm-2。有研究发现气孔特性影响植物的蒸腾作用和净光合速率(Salvucci amp; Crafts-brandner, 2004;Xu amp; Zhou, 2008),单位叶面积高,气体更容易通过气孔,有助于提高气孔导度。气孔导度越大,则CO2交换量大,而CO2是光合作用的原料,证明参与光合作用反应的CO2较多,从而影响净光合速率。本研究中,成年树木的叶面积以及单个气孔面积显著高于幼苗且净光合速率与蒸腾速率同样高于幼苗,相关性分析也发现气孔特性参数均与净光合速率呈现正相关性,说明在青梅的生长与光合特性中,气孔特性扮演着重要角色,这与臭椿的研究结果相似(赵芸玉等,2016)。本研究中成年树木的叶片厚度明显高于幼苗,相关研究表明叶片厚度增加能增强叶片的储水能力,有利于减小叶片的光能损失(Heredia-guerrero et al., 2012;陈健辉等,2015),水分利用率和净光合速率的结果也支持此论点。栅栏组织是叶绿体合成的关键区域,随着组织厚度增加,组织中叶绿素的含量也随之上升,进而提高叶片的光合作用能力,成年树木的叶片相较于幼苗叶片,栅栏组织逐渐增大,细胞排列变得紧密,栅海比升高。同时,相关性分析结果显示叶绿素含量、栅海比与最大净光合速率三者之间均存在显著正相关。因此,推断栅海比和栅栏组织厚度是影响叶绿素含量以及光合速率的重要叶片结构参数,这与油橄榄(刘露等,2016)的光合特性的研究结果相似。结合生境的实际情况推断广西青梅成年树木与其他乔木树种对幼苗的过度遮阴,导致其光能吸收量不足,影响了栅栏组织的发育,进而无法形成足够的叶绿素。较低的叶绿素与孱弱的栅栏组织仅能维持其基本的生存但无法满足植物生长,最终造成了广西青梅始终维持幼苗状态,自然更新困难。
4 结论
综上所述,不同的叶绿素含量和叶片结构是造成广西青梅成年树木与幼苗光合特性差异的重要原因。广西青梅具有幼苗喜阴而成年树木喜阳的光合特性。幼苗光适应能力和光能利用率较弱以及成年树木过度遮阴导致幼苗无法进行充分光合以满足生长需求,是幼苗无法成长为大树的重要原因。因此,在广西就地保护时,可通过人为干扰的方式,对乔木层林木进行人工“开窗”,提供相对充足的光源,以利于广西青梅幼苗生长发育成大树和种群的更新;在引种栽培中,广西青梅幼苗时期需要适当的遮阴处理,以及移栽至有一定荫蔽度的环境中。进入成年期,光适应范围区间变大,光合能力增强,可移植至荫蔽度较小的环境中,以促进广西青梅生长发育。
参考文献:
ADAMEC L, 1997. Photosynthetic characteristics of the aquatic carnivorous plant Aldrovanda vesiculosa [J]. Aquatic Botany, 59: 297-306.
BOND BJ, 2000. Age-related changes in photosynthesis of woody plants [J]. Trends in Plant Science, 5(8): 349-353.
CHEN XH, LIU HQ, ZHANG XJ, et al., 2022. Comparative analysis of photosynthetic and physiological characteristics of red-leafed ‘Quanhong’ and ‘Xuanhong’ poplar cultivars [J]. Acta Horticulturae Sinica, 49(2): 437-447. "[陈兴浩, 刘晗琪, 张新建, 等, 2022. 彩叶杨‘全红’和‘炫红’光合生理特性的比较分析 [J]. 园艺学报, 49(2): 437-447.]
CHENG C, ZHOU W, WANG XB, et al., 2018. Photosynthetic characteristics and antioxidant enzyme activity of Sophora japonica with different ages [J]. J Northwest Forestry University, 33(3): 7-13. "[程程, 周威, 王晓冰, 等, 2018. 不同树龄国槐光合特性和抗氧化酶活性比较研究 [J]. 西北林学院学报, "33(3): 7-13.]
CHAI SF, TANG JM, YANG X, et al., 2015. "Fitting analysis for 4 photosynthesis light response curve models of Keteleeria calcarea [J]. Journal of "Guangxi Academy Science, 31(4): 289-291. "[柴胜丰, 唐健民, 杨雪, 等, 2015. 4种模型对黄枝油杉光合光响应曲线的拟合分析 [J]. 广西科学院学报, 31(4): 286-291.]
CHEN JH, MIAO SY, HUANG LY, et al., 2015. Comparative study on the leaf structures of Sonneratia caseolaris and S. apetala [J]. Plant Science Journal, 33(1): 1-8. "[陈健辉, 缪绅裕, 黄丽宜, 等, 2015. 海桑和无瓣海桑叶片结构的比较研究 [J]. 植物科学学报, 33(1): 1-8.]
CHONDROGIANNIS C, KOTSI K, GRAMMATIKOPOULOS G, et al., 2023. Seasonal differences in leaf photoprotective potential between adults and juveniles of two mediterranean perennials with distinct growth forms: A comparative field study [J]. Plants, 12(17): 3110.
GAO L, LE JX, ZHANG W, et al., 2018. Response to light intensity of Torreya fargesii in two kinds of tree age [J]. Journal of Beijing Forestry University, 40(10): 34-42. "[高岚, 乐佳兴, 张文, 等, 2018. 2种树龄巴山榧对光照的响应 [J]. 北京林业大学学报, 40(10): 34-42.]
HOUTER NC, PONS TL, 2012. Ontogenetic changes in leaf traits of tropical rainforest trees differing in juvenile light requirement [J]. Oecologia, 169: 33-45.
HEREDIA-GUERRERO JA, DE LARA R, DOMNGUEZ E, et al., 2016. Chemical-physical characterization of isolated plant cuticles subjected to low-dose γ-irradiation [J]. Chemistry Physics Lipids, 165(8): 803-808.
JIN YQ, LI DL, 2023. Effects of shading on photosynthetic characteristics and leaf anatomical structure of Ulmus szechuanica seedlings [J]. Acta Botanica Boreali-Occidentalia Sin, 43(6): 1006-1016. "[金雅琴, 李冬林, 2023. 遮光对红果榆幼苗光合作用及叶片解剖结构的影响 [J]. 西北植物学报, 43(6): 1006-1016.]
JIANG YH, SHEN WH, TAN CQ, et al., 2016. The population structure and dynamics analysis and protection strategy of Vatica guangxiensis extremely small population [J]. Ecological Science, 35(6): 67-72. "[蒋迎红, 申文辉, 谭长强, 等, 2016. 极小种群广西青梅种群结构、动态分析及保护策略 [J]. 生态科学, 35(6): 67-72.]
KINOSE K, FUKAMACHI Y, WATANABE M, et al., 2019. Ozone-induced change in the relationship between stomatal conductance and net photosynthetic rate is a factor determining cumulative stomatal ozone uptake by Fagus crenata seedlings [J]. Trees, 34(2): 445-454.
KUBIEN DS, JAYA E, CLEMENS J, 2007. Differences in the structure and gas exchange physiology of juvenile and adult leaves in Metrosideros excelsa [J]. International Journal Plant Sciences, 168(5): 563-570.
LUO M, LIU H, YANG PF, et al., 2019. Study on the characteristics of photosynthetic light response in leaves of Bletilla striata seedlings at different acclimation stages [J]. Seed, 38(9): 52-56. "[罗鸣, 刘海, 杨平飞, 等, 2019. 白及幼苗不同驯化期叶片光合光响应特征研究 [J]. 种子, 38(9): 52-56.]
LIU JY, HE BH, LU XH, 2023. Study on the changes of leaf functional traits of Ligustrum macrophylla with different tree ages [J]. Contemporary Horticulture, 46(3): 38-41. "[刘俊言, 何宝辉, 路兴慧, 2023. 大叶女贞叶功能性状随不同树龄的变化规律研究 [J]. 现代园艺, 46(3): 38-41.]
LIU RX, WANG XJ, YANG CQ, et al., 2018. The study on different model fitting comparison of photosynthesis light response curves for peanut (Arachis hypogaea L.) [J]. Journal of Peanut Science, 47(4): 55-59. [刘瑞显, 王晓婧, 杨长琴, 等, 2018. 花生不同光响应曲线拟合模型的比较 [J]. 花生学报, 47(4): 55-59.]
LIU MX, LIANG GL, 2016. Research progress on leaf mass per area [J]. Chinese Journal of "Plant Ecology, 40(8): 847-860. "[刘明秀, 梁国鲁, 2016. 植物比叶质量研究进展 [J]. 植物生态学报, 40(8): 847-860.]
LIU L, LU LJ, HU YY, et al., 2022. Changes of photosynthesis in leaves of Torreya grandis ‘Merrillii’ in different months and different tree ages [J]. Journal of "Zhejiang A amp; F University, 39(1): 32-40. "[刘琏, 陆吕佳, 胡渊渊, 等, 2022. 不同月份不同树龄香榧叶片光合特性的变化 [J]. 浙江农林大学学报, 39(1): 32-40.]
LI XQ, LU YM, HUANG AM, et al., 2022. Light response model fitting and photosynthetic characteristics of ten different fern species in subtropics [J]. Acta Ecologica Sinica, 42(8): 3333-3344. [李雪琴, 卢艺苗, 黄爱梅, 等, 2022. 亚热带10种蕨类植物光响应模型拟合及光合特性研究 [J]. 生态学报, 42(8): 3333-3344.]
LI DS, BAI QH, LI YJ, et al., 2017. Effects of light conditions on the growth characteristics and photosynthetic traits of Quercus mongolica seedlings [J]. Chinese Journal of "Ecology, 36(10): 2744-2750. "[李东胜, 白庆红, 李永杰, 等, 2017. 光照条件对蒙古栎幼苗生长特性和光合特征的影响 [J]. 生态学杂志, 36(10): 2744-2750.]
LICHTENTHALER HK, BUSCHMANN C, 1987. Chlorophyll fluorescence spectra of green bean leaves [J]. Journal of "Plant Physiology, 129(1): 137-147.
MEZIANE D, SHIPLEY B, 1999. Interacting components of interspecific relative growth rate: Constancy and change under differing conditions of light and nutrient supply [J]. Functional Ecology, 13(5): 611-622.
PAN LP, TANG JM, JIANG HD, et al., 2023. Comparison of photosynthesis and structure of leaves between Manglietia aromatica seedlings and adult plants [J]. Molecular Plant Breeding: "1-11 [2024-02-26]. http://kns.cnki.net/kcms/detail/46.1068.S.20231024.1320.004.html. "[潘李泼, 唐健民, 江海都, 等, 2023. 濒危植物香木莲幼苗与成年植株叶片光合作用及结构比较 [J]. 分子植物育种: 1-11 [2024-02-26]. http://kns.cnki.net/kcms/detail/46.1068.S.20231024.1320.004.html.]
PENG YH, TAN CQ, HE F, et al., 2015. Community compositions and spatial structure feature of Vatica guangxiensis small population in Guangxi [J]. Journal of "Northeast Forestry University, 43(9): 18-22. "[彭玉华, 谭长强, 何峰, 等, 2015. 极小种群广西青梅群落物种及空间结构特征 [J]. 东北林业大学学报, 43(9): 18-22.]
RODRGUEZ-CALCERRADA J, REICH PB, ROSENQVIST E, et al., 2008. Leaf physiological versus morphological acclimation to high-light exposure at different stages of foliar development in oak [J]. Tree Physiology, 28(5): 761-771.
SALVUCCI ME, CRAFTS-BRANDNER SJ, 2004. Inhibition of photosynthesis by heat stress: the activation state of rubisco as a limiting factor in photosynthesis [J]. Physiologia Plantarum, 120(2): 179-186.
TAN CQ, PENG YH, YANG GF, et al., 2016. The physiochemical characteristics of soil and litter in extremely small population of Vatica guangxiensis community [J]. Guangxi Forestry Science, 45(3): 259-265. "[谭长强, 彭玉华, 杨桂芳, 等, 2016. 极小种群广西青梅群落凋落物及土壤理化特征 [J]. 广西林业科学, 45(3): 259-265.]
TUZET A, 2014. Stomatal conductance, photosynthesis, and transpiration, modeling [M]. Netherlands: Springer: 855-858.
WANG RL, LIU JX, CHEN S, 2022. The physiological characteristics of Moringa oleifera Lam. leaves in different growing days in spring [J]. Journal of "Yuxi Normal University, 38(3): 42-46 . [王瑞苓, 刘建祥, 陈诗, 2022. 辣木不同叶龄的叶片在春季的生理特性研究 [J]. 玉溪师范学院学报, 38(3): 42-46.]
XU AZ, JIANG HD, PU QK, et al., 2024. Comparative study on the leaf anatomical structures and photosynthetic characteristics of three species of Geodorum [J]. Guihaia, 44(1): 113-125. "[许爱祝, 江海都, 浦乾琨, 等, 2024. 三种地宝兰属植物叶片解剖结构及光合特性的比较研究 [J]. 广西植物, 44(1): 113-125.]
XIAO YX, HE ZX, SU QZ, et al., 2023. Current distribution and conservation strategy of Vatica guangxiensis (Dipterocarpaceae), a rare and endangered species with extremely small population [J]. Journal of "West China Forestry Science, 52(3): 1-7. "[肖云学, 何在新, 苏启照, 等, 2023. 我国极小种群广西青梅的资源现状及其保护策略 [J]. 西部林业科学, 52(3): 1-7.]
XU ZZ, ZHOU GS, 2008. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass [J]. Journal of "Experimental Botany, 59(12): 3317-3325.
YAN XH, YIN JH, DUAN SH, et al., 2013. Photosynthesis light response curves of four rice varieties and model fitting [J]. Chinese Journal of "Ecology, 32(3): 604-610. "[闫小红, 尹建华, 段世华, 等, 2013. 四种水稻品种的光合光响应曲线及其模型拟合 [J]. 生态学杂志, 32(3): 604-610.]
YE ZP, YANG XL, KANG HJ, 2016. Comparison of light-use and water-use efficiency for C3 and C4 species "[J]. Acta Agriculturae Zhejiangensis, 28(11): 1867-1873. ""[叶子飘, 杨小龙, 康华靖, 2016. "C3 和 C4 植物光能利用效率和水分利用效率的比较研究 [J]. 浙江农业, 28(11): 1867-1873.]
YE ZP, YU Q, KANG HJ, 2012. Evaluation of photosynthetic electron flow using simultaneous measurements of gas exchange and chlorophyll fluorescence under photorespiratory conditions [J]. Photosynthetica, 50(3): 472-476.
ZHANG ZJ, GUO YP, HE JS, et al., 2018. Conservation status of wild plant species with extremely small populations in China [J]. Biodiversity Science, 26(6): 572-577. "[张则瑾, 郭焱培, 贺金生, 等, 2018.中国极小种群野生植物的保护现状评估 [J]. 生物多样性, 26(6): 572-577.]
ZHANG HN, SU ZX, CHEN HB, et al., 2016. Research advances in physiology and molecular biology of flower and fruit development in litchi [J]. South China Fruits, 45(5): 62-64. "[张红娜, 苏钻贤, 陈厚彬, 等, 2016 . 荔枝成花诱导期幼苗和成年树木光合功能的比较 [J]. 中国南方果树, 45(5): 62-64.]
ZHU TJ, YUE CL, JIN SH, 2008. Ecophysiological trait comparison of Shaniodendron subaequale and accompanying species [J]. Journal of "Zhejiang A amp; F University, 25(2): 176-180. "[朱汤军, 岳春雷, 金水虎, 2008. 银缕梅和伴生植物光合生理生态特性比较 [J]. 浙江林学院学报, 25(2): 176-180.]
ZHAO YY, XIA XF, XIONG B, et al., 2016. Leaf epidermal features of Ailanthus altissima(Mill)swingle in different development stages [J]. Plant Science Journal, 34(2): 182-190. "[赵芸玉, 夏晓飞, 熊彪, 等, 2016. 臭椿不同发育阶段叶片表面结构特征 [J]. 植物科学学报, 34(2): 182-190.]
ZHU XL, ZOU R, TANG JZ, et al., 2023. Genetic diversity variation during the natural regeneration of Vatica guangxiensis, an endangered tree species with extremely small populations [J]. Global Ecology and Conservation, 42: e02400.
(责任编辑 周翠鸣)
猜你喜欢
阅读(科学探秘)(2020年8期)2020-11-06 06:22:48
绿色科技(2019年2期)2019-05-21 07:43:20
中国果业信息(2019年1期)2019-01-05 17:41:42
生物学教学(2017年9期)2017-08-20 13:22:32
江苏农业科学(2016年8期)2017-02-15 20:34:22
热带农业科学(2016年11期)2017-01-21 15:10:04
山东农业科学(2016年12期)2017-01-21 14:41:54
山东农业科学(2016年12期)2017-01-21 14:39:43
山东农业科学(2016年11期)2016-12-17 21:16:16
吉林农业·下半月(2016年10期)2016-12-16 23:45:53
