
荔枝花芽分化期间光合特性与碳氮物质变化
2017-01-21 15:10张红娜苏钻贤陈厚彬
热带农业科学 2016年11期
张红娜+苏钻贤+陈厚彬
摘 要 荔枝花芽分化分为诱导、发端和分化3个阶段。试验以早中熟品种‘妃子笑和中熟品种‘桂味为试材,对荔枝花芽分化不同物候阶段叶片净光合速率(Pn)、叶绿素指数(SPAD)、荧光参数和碳氮物质等生理指标的变化规律进行研究。结果表明:‘妃子笑的成花进程早于‘桂味品种,但2个品种在整个花芽分化期间的相关生理指标均呈相似变化趋势,Pn和SPAD在诱导期和抽穗期水平较低,“白点”期和花蕾期有所增加,而荧光参数恰好相反;荔枝成花期间,其可溶性总糖含量在“白点”期阶段水平最低,而淀粉和总氮含量呈相反趋势,在“白点”期达到最高值。可见,不同荔枝品种的光合特性与碳氮物质变化受其花芽分化的阶段性影响更大,而“白点”期是荔枝碳氮物质变化较为关键的转折点。
关键词 荔枝 ;花芽分化 ;光合特性 ;碳氮物质
中图分类号 S667.2 文献标志码 A Doi:10.12008/j.issn.1009-2196.2016.11.014
Abstract The phase changes in floral induction, evocation and differentiation take place in litchi. In order to elucidate the physiological changes during the phase changes, the photosynthetic rate (Pn), chlorophyll index (SPAD), fluorescent parameters, carbohydrate and nitrogen contents of litchi leaves in the field were studied with 'Feizixiao', a early-medium cultivar, and 'Guiwei', a medium cultivar. Results showed that the flowering progress of the 'Feizixiao' litchi trees was earlier than that of 'Guiwei'. But the related physiological indexes of two cultivars had similar change trend during the flower bud differentiation. During the flowering, the changes of Pn and SPAD were lower at floral induction and heading stage, higher at the stage of "whitish millets" and alabastrum, but the fluorescent parameters were just opposite to them. At the "whitish millets" stage, the contents of the soluble sugars were the lowest, whereas starch and total nitrogen were the highest. Therefore, the changes of photosynthetic characteristics and carbon-nitrogen content were greatly influenced by the flowering phase, and the period of "whitish millets" was the key turning point of the changes of carbohydrate and nitrogenous compounds in litchi.
Keywords Litchi (litchi chinensis Sonn.) ; Flower differentiation ; Photosynthetic characteristics ; Carbon and Nitrogen
光合作用是果树生长发育的基础,而碳水化合物是光合作用的直接产物,对植物的成花座果具有十分重要的意义。荔枝(Litchi chinensis Sonn.)花芽分化期是其生长发育的关键时期,也是一个复杂的形态建成过程,荔枝成花诱导和分化期间碳同化及再分配可能对成花过程产生重要的影响[1]。前人关于荔枝光合特性方面的研究已有很多报道[2-12],主要集中在外部因素和栽培措施等方面,如不同品种、不同发育期的光合作用[2,4,6-7],钾[5]、水分胁迫[8]、遮荫[9-10]、环剥[11]和间伐回缩[12]等对光合作用的影响,但针对荔枝花芽分化这一关键时期的光合生理研究尚未见报道。
随着对荔枝花芽分化“阶段性”的提出[13],国内外学者对荔枝成花期间碳水化合物积累和再分配模式分析变得更加具体和有针对性[14-17]。Nakata等[14]研究发现,在荔枝花芽分化前的低温诱导期间叶和茎内淀粉含量达到最高值。陈厚彬等[15]也发现,荔枝成花诱导过程可导致碳水化合物的再分配,高成花树与低成花树中淀粉的积累模式不同,出现多种淀粉积累的分配方向。袁沛元等[14]针对‘妃子笑现蕾开花期叶片碳水化合物的变化进行研究,发现其结果母枝叶片总糖含量在现蕾开花期不断减少。肖华山等[16]研究分析了荔枝雄蕊和雌蕊发育期碳氮化合物的动态变化,发现C/N较大有利于雄蕊分化,反之则有利于雌蕊分化。吴志祥等[17]比较分析不同品种在花芽分化过程中不同方位、不同部位枝条顶芽和花序中碳氮化合物的动态变化,认为C/N比值较高有利于荔枝花芽分化。
前人的研究大多数集中在成花某一具体阶段的光合同化物积累动态上,但关于荔枝整个成花过程中其光合能力和碳氮营养储备规律的系统研究尚未见报道。因此,本文拟通过分析2个主栽品种在荔枝成花各个关键时期叶片光合、荧光参数和碳水化合物含量等生理指标的变化,弄清荔枝成花各时期的光合能力和碳氮营养积累规律,以期能为花芽分化不同阶段的营养积累与调控提供指导。
1 材料与方法
1.1 材料
试验于2009年12月至2010年3月在华南农业大学荔枝园内进行。供试品种为2个主栽品种‘妃子笑(Litchi chinensis Sonn. cv. Feizixiao)和‘桂味(Litchi chinensis Sonn. cv. Guiwei)各5株,2003年春季种植,自根苗,常规管理。
1.2 方法
1.2.1 成花物候期及成花状况观测统计
自2009/2010年度11月起到3月,每隔5~10 d记载各植株的枝梢老熟状况、现“白点”、抽花序和现花蕾等物候期。物候观察标准参考Menzel and Simpson[18]。在2个品种成花时,调查花序枝类型,单株小区,每株调查的末端枝梢总数量大于50枝。
1.2.2 田间荔枝树光合作用指标测定
在‘妃子笑和‘桂味2个品种成花的不同阶段,分别选取晴朗、无风天气的上午,诱导期(2009-12-05;2009-12-15)、现“白点”(2009-12-25;2010-01-05)、抽花序(2010-01-28;2010-02-09)和现花蕾(2010-02-11;2010-02-22),每个单株从东南西北4个方向,选树体中部成熟叶片(从顶部向下数第3、4片复叶中部的小叶)测定光合作用和叶绿素指数。气体交换参数采用美国Li-COR公司生产的Li-6400便携式光合仪进行测定。测定时光合有效辐射(PAR)设为1 200 μmol/(m2·s)、大气CO2浓度(Ca)设为400 μmol/mol,大气温度(Ta)、叶温(TL)、相对湿度(RH)等参数均采用仪器自控系统控制,随外界环境变化而变化。
1.2.3 叶绿素荧光参数和叶绿素指数的测定
在测定光合参数的同一时间,使用FM S-2便携调制式荧光仪活体测定叶绿素荧光动力学参数。在自然光下测定初始荧光(Fo)、最大荧光(Fm)、最大光化学效率(Fv/Fm)等,再将叶片用暗适应夹暗适应20 min后测PSⅡ等指标。测定上述指标时随机选取不同方向的健壮枝条,从顶部向下数第3、4片复叶中部的小叶。每株选取4个枝条,重复测定3次。
因SPAD可以表征叶片叶绿素的含量[19],是活体测定叶绿素含量的一种简捷方法。在每次测定光合和荧光参数后,使用SPAD-502型号便携式叶绿素仪测定叶绿素指数。
1.2.4 叶片中碳氮物质的测定
在每次测定光合和荧光参数后的11:00左右,迅速将当天测定的叶片周围枝条类似位置叶片采集下来,每个品种各采20片。 放置冰盒中带回实验室用清水、蒸馏水清洗干净,置烘箱中105℃杀青15 min,65℃烘干24 h,再用粉碎机粉碎,过100目筛,装入封口袋,并置于干燥器中备用。可溶性总糖(以干重计)采用蒽酮-硫酸法进行测定[20],淀粉采用碘显色法测定[21],总氮含量(以干重计)测定用凯式定氮法[22]。
1.2.5 统计分析
数据统计分析使用Microsoft Office 2010和SPSS 16.0等软件进行处理。
2 结果与分析
2.1 荔枝成花物候期调查
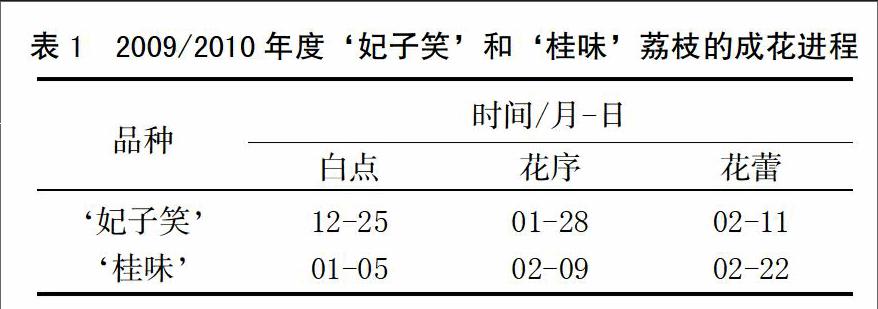
‘妃子笑和‘桂味成花进程存在一定差异(表1),‘妃子笑出现“白点”、花序抽出及花蕾显现的时间均早于‘桂味。‘妃子笑在2009年12月25日左右出现“白点”,而‘桂味在2010年1月5日前后顶芽才开始现“白点”;‘妃子笑在2010年1月28日前后抽出花序,而‘桂味则推迟至2010年2月9日左右;‘妃子笑在2010年2月11日前后小花蕾开始显现,而‘桂味则在2010年2月22日左右才能肉眼分辨出单独的花蕾。
2.2 荔枝成花不同阶段叶片光合生理特性分析
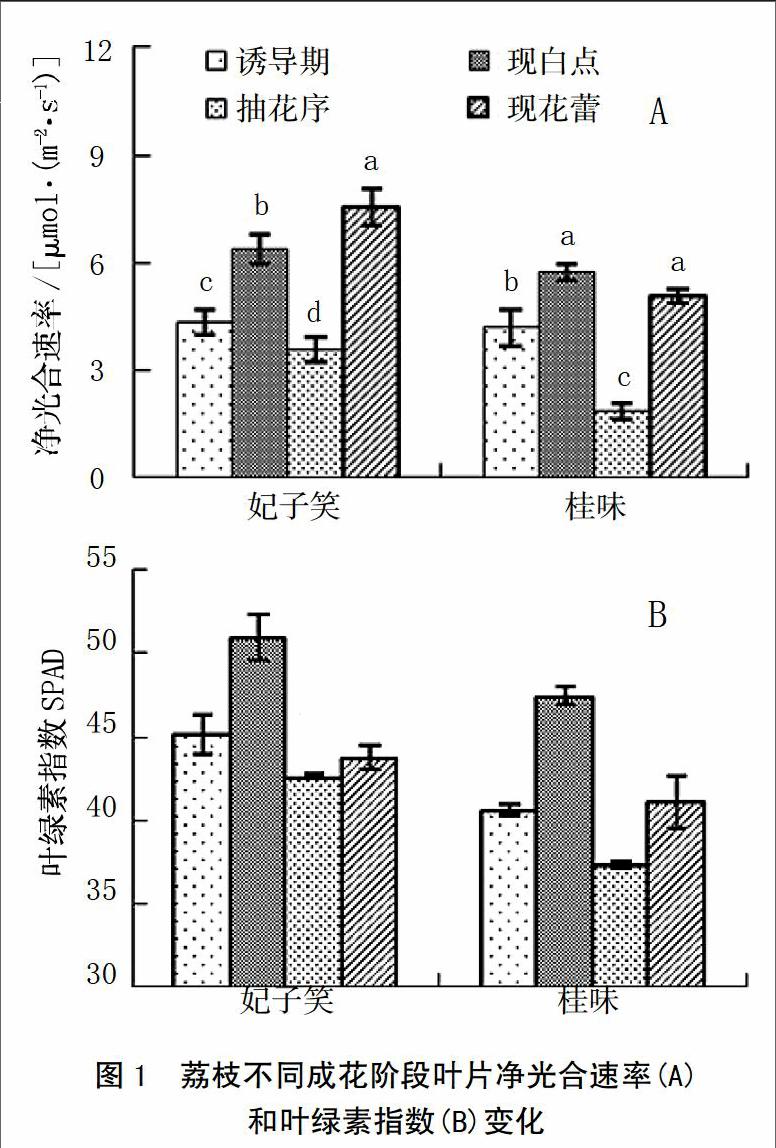
从图1-A可以看出,在整个成花期,2个荔枝品种Pn变化趋势表现一致,在成花诱导期较低,现“白点”期时有所上升,而抽花序期Pn出现短暂的下降,花蕾期又有所升高。其中‘妃子笑4个阶段之间的Pn值存在显著差异,现花蕾时的Pn较其它几个阶段的高,为7.57 μmol/(m2·s),而抽花序时的Pn则最低3.59 μmol/(m2·s)。2个品种在成花期间的SPAD也呈现相似的变化趋势,在“白点”期时达到最高值(图1-B)。
2.3 荔枝成花不同阶段叶片叶绿素荧光参数分析
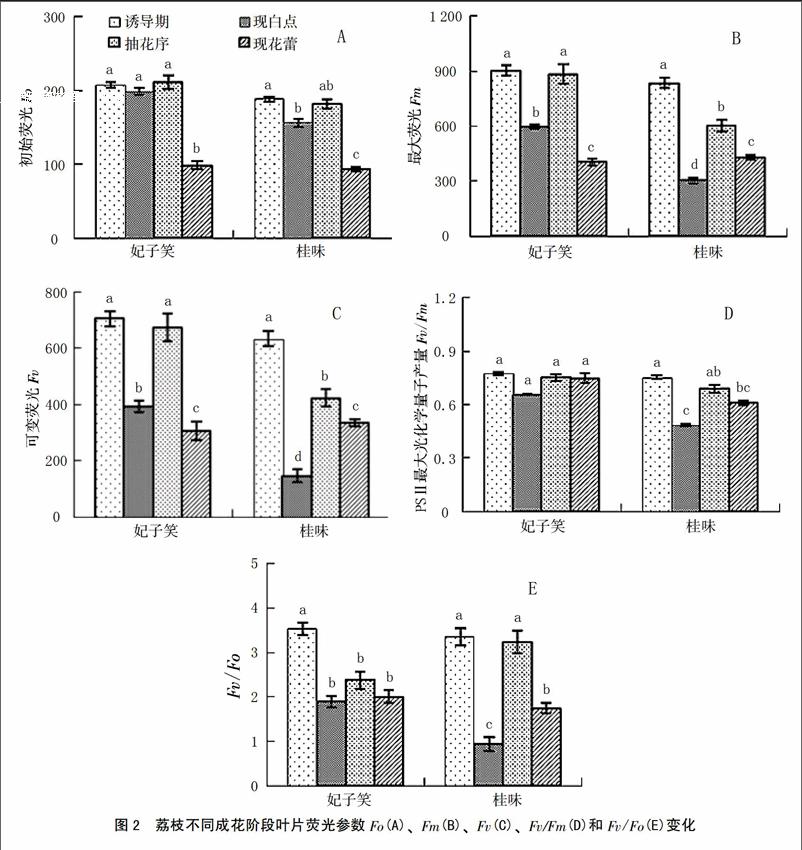
叶绿素荧光动力学测定结果表明:成花过程中‘妃子笑和‘桂味2个品种现花蕾阶段的Fo分别为98.41和93.39,显著低于该品种其它3个阶段的Fo(198.59~211.75;156.50~188.19)(图2-A);与Pn和SPAD的变化趋势相反,2个品种的Fm、Fv和Fv/Fo在现“白点”和现花蕾阶段均较其它2个阶段低;其中,‘桂味的Fm和Fv在诱导期值最高,分别为833.97和633.12,至现“白点”时急剧下降,抽花序时有所上升,随后又有所下降(图2-B、2-C、2-E)。‘妃子笑的Fv/Fm在不同成花阶段无显著差异,而‘桂味现“白点”阶段的Fv/Fm为0.49,显著低于该品种其它3个阶段(0.61~0.76)(图2-D)。
2.4 荔枝成花不同阶段叶片碳水化合物含量变化
从图3-A可以看出,在荔枝成花阶段的各个时期,可溶性总糖含量的变化趋势一致,均在现“白点”时达到最低点,之后随着花序抽出和花蕾的出现又逐渐上升。但与可溶性总糖的变化趋势恰好相反,‘妃子笑和‘桂味叶片中淀粉含量在现“白点”时最高,分别为12.75和18.21 mg/g,随着花序的抽出又逐渐下降(图3-B)。在不同成花阶段,2个品种叶片中总氮含量变化趋势基本一致,在花芽诱导期到“白点”期时,经历了一个上升的过程,至现“白点”时含量最高,分别为22.28 和21.50 mg/g,进入形态分化后逐渐下降(图3-C),这与淀粉的变化规律较为相似,但与可溶性总糖的变化趋势恰好相反。
3 结论与讨论
3.1 荔枝不同成花阶段光合和荧光特性变化
在整个成花过程中,‘妃子笑和‘桂味Pn和SPAD的变化趋势相似,均是在成花诱导期较低,由生理分化转向形态分化(现“白点”)时有所上升,在抽穗期出现短暂下降,花蕾期又回升。叶片光合作用效率受季节和外界环境影响很大,尤其是光照和温度[23-24]。荔枝成花诱导期需要较低的温度,导致Pn和SPAD较低;“白点”出现期,外界温度回暖,Pn和SPAD也随之上升;而花序分化期一般昼间温和、夜间冷凉,导致Pn和SPAD有所下降;随着温度升高,光照增强,花蕾期Pn和SPAD又有所上升。叶绿素荧光参数是反映植物光合能力强弱的重要指标[25]。从图2可以看出,在不同成花阶段,2个品种的Fo、Fm、Fv、Fv/Fo和Fv/Fm在现“白点”和现花蕾阶段的值较低。Fv/Fm、Fv/Fo是反映PSⅡ最大光化学效率和潜在活性的重要参数[25]。Genty等[26]的研究表明,Fv/Fm在非胁迫条件下比较恒定,一般介于0.80~0.85,不受物种和生长条件的影响,但在胁迫条件下,该参数明显下降。
3.2 荔枝不同成花阶段叶片碳氮物质的变化
Kozlowski指出,花芽分化期最重要是涉及碳水化合物的源库方向交换,即成花诱导期间,花发端部位必须成为调运碳水化合物的库[27]。因此,果树的花芽分化需要较多的可溶性总糖[28]。从图3可以看出,在荔枝成花的各个阶段,2个品种的可溶性总糖含量均是在诱导期含量较高,花发端时(“白点”期)处于低谷,之后随着花序抽出和花蕾的出现,又逐渐上升,这与在枇杷[29]、龙眼[30]和杨梅[31]上的研究结果基本一致。荔枝在花芽诱导期可溶性糖含量较高,这可能是贮藏的淀粉降解增加了可溶性糖的含量导致。进入“白点”期后其含量迅速下降,可能是由于顶端花芽生理分化消耗了大量的可溶性糖来合成各种含碳物质。随着花芽形态分化的完成,可溶性糖含量又逐渐升高,这可能与当时叶片的光合作用密切相关。生产中常通过间伐并结合适度回缩及疏剪等处理,通过改善树体间通风透光性、提高光合能力来提高树体营养及成花座果率[32]。‘妃子笑叶片中可溶性总糖含量在成花的各个阶段均高于‘桂味,这可能与‘妃子笑的光合能力比‘桂味高有关,也暗示了高水平可溶性总糖可能是‘妃子笑较‘桂味更早进入花芽分化阶段的重要原因。
淀粉是果树叶片合成碳水化合物的常见积累形式,分解为可溶性糖后被植物相关器官利用。本研究结果表明,2个品种叶片中淀粉含量的变化与可溶性糖的变化趋势恰好相反,说明淀粉和可溶性糖之间可以相互转化(图3-B)。荔枝采收后通过2~3次梢的营养生长,在花芽生理分化期前积累了大量淀粉等碳水化合物,为花芽分化做了充足的营养储备。随着进入花芽的形态分化,淀粉的含量逐渐降低,是由于淀粉转化成可溶性糖,来满足花芽分化期间较为旺盛的新陈代谢。图3中,总氮含量与可溶性糖含量的变化呈相反趋势,在进入花芽形态分化后,叶片中的总氮含量持续下降,可能由于花芽分化合成大量蛋白质消耗所致。荔枝在成花诱导期总氮含量较低,表明在花芽生理分化期总氮含量高不利于花芽分化。因此,荔枝在生产上若想提早花芽分化,必须在其诱导期严格控制氮肥的施用。在苹果梨[33]上有类似结论。荔枝的花芽分化一般处于冬春季节,而冬春季荔枝的光合作用处于一个相对较弱的时期,碳水化合物的积累和再分配不仅影响荔枝花芽的诱导与分化,还可能间接影响到荔枝的开花座果。在“白点”出现之前,光合产物的积累可能是影响荔枝花诱导、发端及分化的原因之一,而现白点之后,花芽进入分化阶段,使花芽成为一个强“库”,需要消耗大量的碳氮营养,从而可能反过来间接影响“源”叶的光合作用。
参考文献
[1] 陈厚彬,苏钻贤,张 荣,等.荔枝花芽分化研究进展[J]. 中国农业科学,2014,47(9):1 774-1 783.
[2] 张祝平,招晓东,叶耀雄,等.荔枝的光合特性[J]. 应用与环境生物学报,1995,1(3):225-231.
[3] 孙谷畴. 荔枝的光合特性[J]. 武汉植物学研究,1987,5(2):165-172.
[4] 张规富. 荔枝幼树光合特性的研究[D]. 长沙:湖南农业大学,2004.
[5] 邓义才,倪耀源,陈乃荣. 钾对荔枝光合作用和呼吸作用的影响[J]. 华南农业大学学报,1994,15(4):80-84.
[6] Hieke S, Menzel C M, Lüdders P. Effects of leaf, shoot and fruit development on photosynthesis of lychee trees (Litchi chinensis)[J]. Tree Physiology, 2002, 22(13): 955-961.
[7] 范 妍,尹金华,罗 诗,等. 荔枝新品种岭丰糯的光合性能指标研究[J]. 中国南方果树,2011,40(4):13-15.
[8] 宋世文,王泽槐,赵晓勤,等. 水分胁迫对成年荔枝不同季节光合速率日变化的影响[J]. 热带亚热带植物学报,2007,15(6):482-486.
[9] 莫伟平,周琳耀,张静逸,等. 遮荫和环剥对荔枝枝梢生长和光合生理的影响[J]. 园艺学报,2013,40(1):117-124.
[10] 周琳耀,莫伟平,张静逸,等. 遮阴对荔枝光合特征及矿质营养积累的影响[J]. 果树学报,2014,31(2):270-275.
[11] 戴宏芬,邱燕萍,袁沛元,等. 螺旋环剥对幼龄‘桂味荔枝果期光合和蒸腾作用的影响[J]. 园艺学报,2010,37(8):1 241-1 246.
[12] 戴宏芬,邱燕萍,袁沛元,等. 间伐回缩修剪对荔枝叶片光合和蒸腾作用的影响[J]. 果树学报,2016,33(6):701-708.
[13] 黄辉白,陈厚彬.以阶段观剖视荔枝的花芽分化[J]. 果树学报,2003,20(6):487-492.
[14] Nakata S, Watanabe Y. Effects of Photoperiod and Night Temperature on the Flowering of Litchi chinensis[J]. Botanical Gazette, 1966, 127(2/3):146-152.
[15] 陈厚彬,黄辉白,刘宗莉.荔枝树成花与碳水化合物器官分布的关系研究[J]. 园艺学报,2004,31(1):1-6.
[16] 肖华山,吕柳新,肖祥希.荔枝花雄蕊和雌蕊发育过程中碳氮化合物的动态变化[J].应用与环境生物学报,2002,8(1):26-30.
[17] 吴志祥,王令霞,陶忠良,等. 2个荔枝品种花芽分化期碳氮营养的变化[J]. 热带作物学报,2006,27(4):25-28.
[18] Menzel C M, Simpson D R. Effect of temperature on growth and flowering of litchi (Sonn.) cultivars[J]. Journal of Horticultural Science, 1988, 63(2): 349-360.
[19] 徐照丽,李天福.SPAD-502叶绿素仪在烤烟生产中的应用研究[J]. 贵州农业科学,2006,34(4):23-24.
[20] 邹 琦. 植物生理学实验指导[M]. 北京:中国农业大学出版社,2000:110-123.
[21] 徐昌杰,陈文峻,陈昆松,等. 淀粉含量测定的一种简便方法-碘显色法[J]. 生物技术,1998,8(2):41-43.
[22] 母建华. 基于光谱分析的茶鲜叶全氮含量快速检测技术[D]. 镇江:江苏大学,2008:23-24.
[23] 曾光辉,郭延平,刘 辉.果树光合作用的环境调控[J]. 北方园艺,2004(3):32-33.
[24] 谷 镜,程晓建,张 敏. 环境因子对常绿果树光合作用的影响[J]. 福建林业科技,2009,36(3):270-274.
[25] 陈建明,俞晓平,程家安. 叶绿素荧光动力学及其在植物抗逆生理研究中的应用[J]. 浙江农业学报,2006,18(1):51-55.
[26] Genty B, Briantais J M, Baker N R. The relationshipbetween the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence[J]. Biochimica et Biophysica Acta (BBA)-General Subjects, 1989, 990(1): 87-92.
[27] Kozlowski T. Carbohydrate source and sinks in wood plant[J]. Botanical review, 1992, 58(2):107-222.
[28] 钟晓红,罗先实,陈爱华.李花芽分化与体内主要代谢产物含量的关系[J]. 湖南农业大学学报,1999,25(1):31-35.
[29] 胡 军. 枇杷花芽分化和某些物质代谢关系的研究[J]. 浙江农业大学学报,1983,9(4):307-314.
[30] 陈清西,李松刚. KClO3诱导龙眼成花及其叶片碳水化合物与蛋白质的变化[J]. 福建农林大学学报(自然科学版),2004,33(2):182-185.
[31] 李兴军,李三玉,汪国云,等. 杨梅花芽孕育期间酸性蔗糖酶活性及糖类含量的变化[J]. 四川农业大学学报,2000,18(2):164-166.
[32] 邱燕萍,袁沛元,凡 超,等. 间伐回缩修剪对荔枝树体营养及成花坐果的影响[J]. 中国热带农业,2015(4):74-76.
[33] 郭金丽,张玉兰. 苹果梨花芽分化期蛋白质、淀粉代谢的研究[J]. 内蒙古农牧学院学报. 1999,20(2):80-82.
猜你喜欢
科普童话·学霸日记(2021年6期)2021-09-05
汉语世界(The World of Chinese)(2021年1期)2021-02-22
北方音乐(2020年9期)2020-07-09
果农之友(2020年5期)2020-06-21
农业与技术(2019年5期)2019-05-29
农民致富之友(2018年21期)2018-12-27
艺术评鉴(2017年1期)2017-03-24
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年11期)2016-12-17
