
濒危植物油丹种群结构与动态特征
2025-02-16 00:00:00贵新丽叶储民陈玉凯张凯吴庭天
广西植物 2025年1期
关键词:存活曲线
摘 要: "油丹(Alseodaphnopsis hainanensis)是国家二级重点保护野生植物。为了探讨油丹种群的生存状况,该文根据样地调查数据,利用径级代替龄级,编制种群静态生命表,分析种群数量特征,同时应用种群动态变化指数及时间序列模型来分析预测该种群的动态及未来发展趋势。结果表明:(1)无论是原生林还是次生林,油丹的种群龄级结构均呈现倒“J”型,各龄级均有个体分布;与原生林相比,次生林的幼苗数量更多。(2)在原生林和次生林中,油丹种群的动态指数在相邻的龄级间有一定的波动,原生林和次生林的动态指数Vpi和V′pi都大于0,这表明该种群目前是增长型,但种群对外界干扰比较敏感,抗干扰能力较差;次生林的抗干扰概率相对较小,表明次生林油丹种群的抗干扰能力强于原生林。(3)静态生命表显示,原生林和次生林的油丹种群存活数和个体生命期望值都随着龄级的增加逐渐下降;种群存活曲线趋近于Deevey-Ⅱ型。(4)时间序列分析预测,在经历未来2~3个龄级时间后,原生林Ⅲ龄级的个体数出现下滑;在经历未来2个龄级时间后,次生林Ⅲ龄级的个体数出现下滑;在经历未来5个龄级时间后,原生林和次生林中所有龄级个体数均呈稳定增长。综上认为,不管是原生林还是次生林,油丹种群均为增长型,种群处于波动状态,对外界干扰敏感;种群幼龄个体能补充各龄级个体死亡造成的损失,维持种群稳定性。为促进油丹种群自然更新,应加强就地保护措施,深入开展幼苗更新、繁育和遗传多样性研究等工作,同时可以采取适度的人为措施,提高幼苗到幼树的转化率,促进油丹种群数量增长。
关键词: 油丹, 濒危植物, 种群结构, 存活曲线, 生存分析, 动态指数
中图分类号: "Q948
文献标识码: "A
文章编号: "1000-3142(2025)01-0161-11
收稿日期: "2024-07-29 接受日期: "2024-09-26
基金项目: "海南省重点研发计划项目(ZDYF2023RDYL01); 海南国家公园研究院资助项目(KY-24ZK02); 海南省自然科学基金项目(320MS038)。
第一作者: 贵新丽(1999—),硕士研究生,研究方向为海南濒危植物保育,(E-mail)1670952706@qq.com。
*通信作者: 陈玉凯,博士,副教授,研究方向为海南濒危植物的保育和恢复,(E-mail)chenyukai@hainnu.edu.cn。
Population structure and dynamic characteristics of
endangered plant Alseodaphnopsis hainanensis
GUI Xinli"YE Chumin"CHEN Yukai1*, ZHANG Kainbsp;WU Tingtian2
( 1. Ministry of Education Key Laboratory for Ecology of Tropical Islands, College of Life Sciences, Hainan Normal University,
Haikou 571158, China; 2. Hainan Academy of Forestry (Hainan Academy of Mangrove), Haikou 571199, China )
Abstract: "Alseodaphnopsis hainanensis is a national second-class key protected plant in China. In order to explore the survival status of its populations in Hainan Province, a field quadrat survey was conducted. Based on the sample site survey data, population dynamics and future development trend were predicted by replacing the age class with the diameter class, compiling the static life table of the population, analyzing the quantitative characteristics of the population, using the population dynamic index and the time series model. The results were as follows: (1) The age-class structure of A. hainanensis populations exhibited an inverted J-shape in primary and secondary forests, and the studied trees covered all age classes; compared to primary forests, secondary forests had more seedlings. (2) The dynamic indices of A. hainanensis populations in primary and secondary forests fluctuated between neighboring age classes, and the values of Vpi and V′pi were greater than 0, which indicated that the populations were the growth type, but it were more sensitive to external disturbances, and had poorer anti-disturbance ability; "the anti-interference probability of secondary forest was relatively small, which showed that secondary forest population was more resistant to disturbances than primary forest. (3) The static life table showed that number of surviving A. hainanensis populations and individual life expectancies decreased gradually with increasing age classes in both primary and secondary forests; the population survival curve tended to be Deevey-Ⅱ"type. (4) The time series prediction showed after experiencing the time of next 2-3 age classes, there was a decline in number of individuals in the age class Ⅲ of primary forest; while after experiencing the time of next 2 age classes, there was a decline in number of individuals in the age class Ⅲ of secondary forest; number of individuals at all age classes in both primary and secondary forests showed a steady increase after the the time of next "5 age classes. In summary, populations of A. hainanensis are growth-oriented, and the populations are in a fluctuating state and sensitive to external disturbances; young individuals of the population can replenish the losses caused by the death of individuals of all age classes and maintain population stability. Measures should be taken to promote the natural regeneration of A. hainanensis populations, such as strengthening in-situ conservation, strengthening research on seedling renewal and breeding, carrying out research on genetic diversity, and meanwhile, conducting moderate anthropogenic measures to increase the conversion rate of seedlings to saplings, and to promote population growth of A. hainanensis.
Key words: Alseodaphnopsis hainanensis, endangered plants, population structure, survival curves, survival analysis, dynamic indices
种群生态学的核心研究领域包括种群结构和数量动态(张婕等,2014;何佳宁等,2024)。种群结构由径级结构、高度结构和年龄结构等多方面组成(胡千惠等,2023),既可以反映种群内个体的年龄、大小和数量,又可以显示种群数量的动态及变化趋势。根据静态生命表建立的存活曲线,数量动态指数和时间序列预测模型等都是种群统计学的重点研究方法,这些方法不仅有助于分析植物种群当前的生存状况及其与环境的互动关系(范海兰等,2004),还能评估历史上的干扰因素,更重要的是它们能预测种群未来的发展趋势(Omelko et al.,2018;王泳腾等,2021)。通过对种群结构和数量动态的深入研究,能够为植物资源的保护提供有力的科学依据,并有效规划植物资源的合理利用,以确保其可持续发展。
油丹(Alseodaphnopsis hainanensis)是樟科(Lauraceae)乔木树种,是中国海南岛热带山地雨林树种,除越南北部外仅在中国海南岛有野生分布,主要生于海拔700~1 700 m的林谷或密林中,是国家Ⅱ级重点保护野生植物(陈玉凯等,2011)。油丹树干通直,材质优良,为热带名贵用材。由于长期利用和不合理的采伐,以及原生境的破碎化,加之种群更新经常失败,致使天然资源已面临枯竭(陈玉凯等,2011)。油丹群落的生境类型可分为两大类:第一类是受人为活动干扰很少或无人为干扰的原生林,这些原生林相对较为原始,受到的人为影响较小,仍然保持着较为自然的状态;第二类是经常受到人为活动(如砍伐、开垦和污染等)干扰的次生林。目前,关于油丹的研究主要集中在生境和生物学特征描述(黄桂华等,2011)、种间联结(陈玉凯等,2011)、化学成分提取(常海涛等,2011)、种苗培育(苟志辉等,2017)等方面。陶建平和臧润国(2004)对海南霸王岭热带山地雨林的林隙树种进行研究,发现油丹属于阳性树种,主要分布在幼年林隙或大林隙中。陈玉凯等(2011)对海南霸王岭油丹所在群落主要树种的种间联结性进行研究,发现油丹所在群落主要树种间在总体上表现出不显著的正联结关系,反映了该地区油丹所处群落曾受到一定的破坏且分布范围较窄,种群较小。然而,当前对海南岛油丹种群情况并不清楚,制约了该物种的保护成效。深入研究油丹的种群结构与动态,了解其生存现状及对未来的动态进行预测,有利于油丹种群的保护和管理。为此,本研究以海南热带雨林国家公园及周边为研究区域,采用实测法和典型抽样法对海南岛的油丹种群进行调查,通过分析原生林和次生林中油丹种群的龄级结构、静态生命表、存活曲线和时间序列预测,探讨两种生境中油丹的种群结构、数量动态变化规律、主要致濒原因等科学问题,揭示油丹种群的生存现状和未来发展趋势,旨在为油丹种群的保护和可持续利用提供科学依据。
1 研究区概况及研究方法
1.1 研究区概况
海南岛面积约3.39×104 km2(刘少军等,2015),气候类型为热带季风气候,地处中国的最南端,地理坐标为108°37′—111°03′ E、 18°10′—20°10′ N。该区域年均气温22~27 ℃,最炎热时月平均温度可至28.4 ℃,而每年最严寒时月平均温度可至17.2 ℃(李肇晨等,2015)。每年平均降水量充沛,约为1 639 mm,具分明的少雨季和多雨季,少雨季位于每年的11月到次年4月,多雨季位于每年5月到10月。玄武岩和花岗岩是海南岛土壤的主要成土母质岩,砖红壤、黄壤和赤红壤为该地区常见的3种基本土壤种类(龚子同等,2004)。油丹群落乔木层伴生树种主要有黄叶树(Xanthophyllum hainanense)、红鳞蒲桃(Syzygium hancei)、厚壳桂(Cryptocarya chinensis)、白颜树(Gironniera subaequalis)、长柄梭椤(Reevesia longipetiolata)、米槠(Castanopsis carlesii);灌木层植物主要有九节(Psychotria asiatica)、粗丝木(Gomphandra tetrandra)、琼岛染木树(Saprosma merrillii)、三角瓣花(Prismatomeris tetrandra)、亮叶猴耳环(Archidendron lucidum)、郎伞木(Ardisia crenata);草本层植物主要有双盖蕨(Diplazium donianum)、粗叶卷柏(Selaginella trachyphylla)、单叶新月蕨(Pronephrium simplex)、乌毛蕨(Blechnopsis orientalis)。
1.2 研究方法
1.2.1 样地设置与调查方法
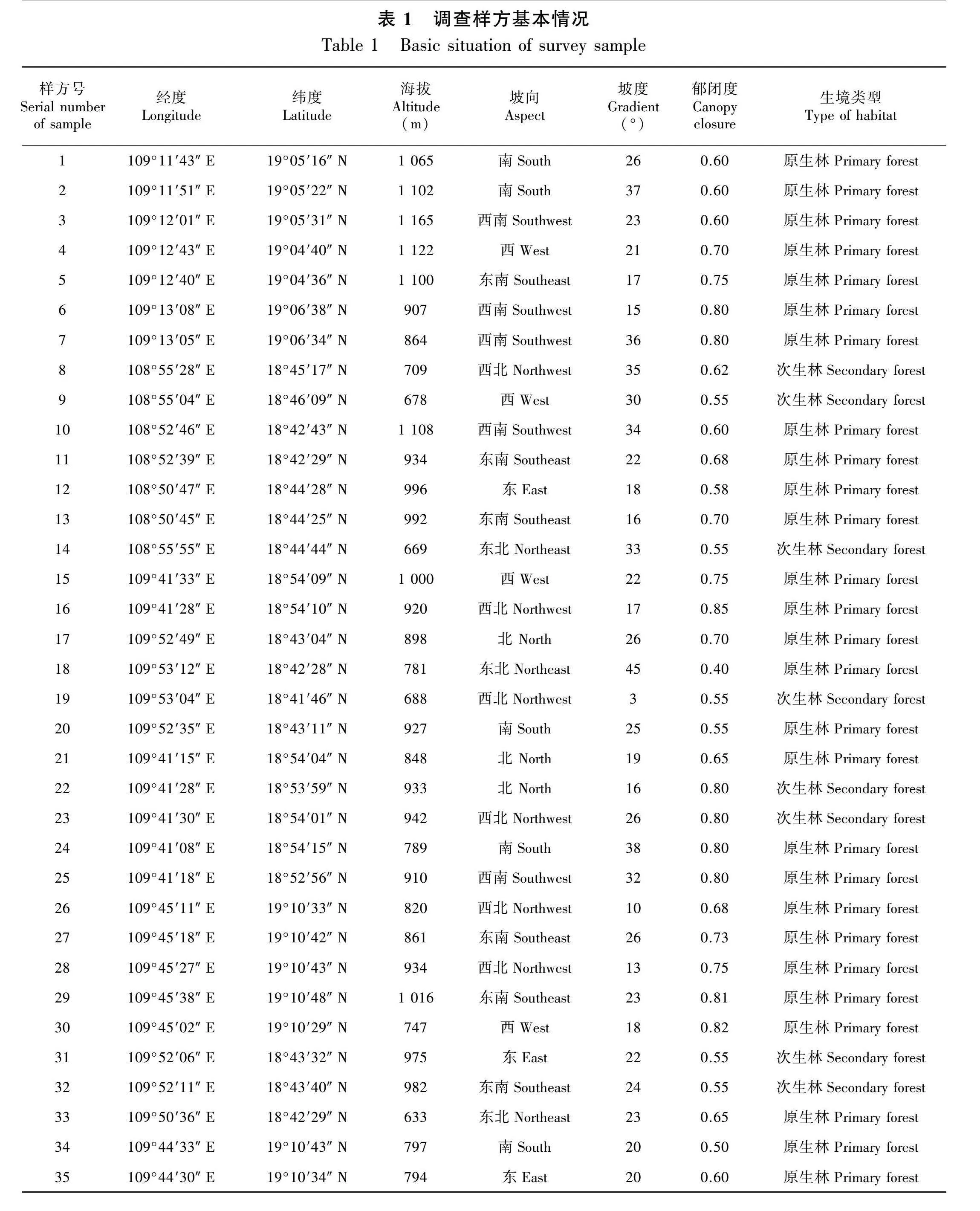
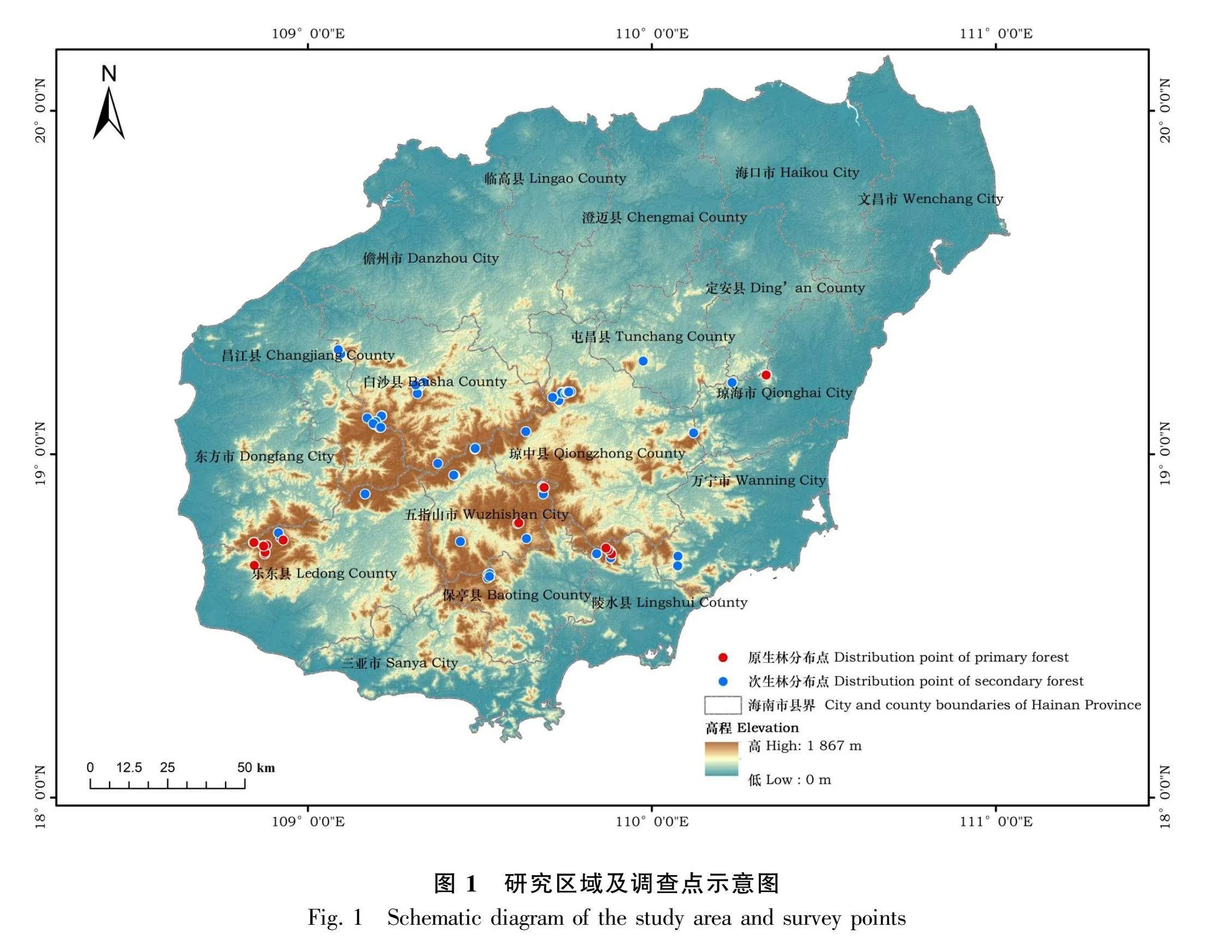
本研究采用实测法和典型抽样法对海南岛的油丹种群进行调查。海南热带雨林国家公园范围内,油丹连片分布,采用典型抽样法,在霸王岭、尖峰岭、五指山、吊罗山、黎母山设置样方进行调查,其中27个为次生林,8个为原生林(图1),每个样方大小为20 m × 20 m,总计35个样方14 000 m2;将每个大样方划分成4个10 m × 10 m的小样方,测量记录各样方内胸径(DBH)≥1 cm的油丹个体的胸径、树高、冠幅,同时用GPS定位样方,记录样地的海拔、坡度、坡向等基本信息。样地具体情况见表1。海南热带雨林国家公园范围外,由于油丹的分布范围有限且种群规模较小,适于直接计数,因此这些地区的油丹种群调查采用实测法,测定油丹个体的胸径、树高、冠幅等指标,同时采用GPS定位,记录物种所处的海拔、坡度、坡向等基本信息。
1.2.2 龄级结构划分 油丹在野外分布数量有限,不适合采用生长锥或解析木来测定植株年龄,因此采用空间替代时间,并结合应用生态学中的大小结构分析法来探究油丹种群的动态变化。本研究以胸径大小取代年龄大小来探究油丹种群动态(Frost amp; Rydin, 2000)。以其生物学特征及实地调查结果作为油丹龄级的划分依据,参照罗文等(2023)对坡垒径级的划分方法,油丹种群的龄级划分如下:幼苗阶段(龄级Ⅰ),包括DBH小于或等于2.5 cm的个体;幼树阶段(龄级Ⅱ),是指DBH在2.5 cm到7.5 cm之间(包括7.5 cm)的个体;小树阶段(龄级Ⅲ),是指DBH在7.5 cm到22.5 cm之间(包括22.5 cm)的个体;中树阶段(龄级Ⅳ),是指DBH在22.5 cm到42.5 cm之间(包括42.5 cm)的个体;大树阶段(龄级Ⅴ)则适用于DBH超过42.5 cm的个体。
1.2.3 种群龄级结构动态的量化方法 为客观、准确地评价油丹种群动态特征,利用陈晓德(1998)提出的种群动态量化法对油丹种群动态进行研究,Vn、Vpi、V′pi可以体现种群的变化趋势,正值表示种群数目增长,零表示种群数目稳定,负值表示种群数目衰退(申仕康等,2008)。
1.2.4 种群静态生命表建立 生命表为反映种群目前生存情况及判定油丹发展趋势的重要指标,根据静态生命表的编写体系(江洪,1992),编写油丹种群的静态生命表。油丹种群静态生命表由9个参数所组成(张悦等,2015)。由于静态生命表无法记录一个种群从诞生到消亡的完整过程,仅反映了特定时间内不同年龄阶段群体的动态变化情况,因此在野外调查中存在一定的非随机误差。由于在油丹种群静态生命表编制过程中会出现死亡率为负值的情况,因此采用匀滑处理技术(解婷婷等,2014;胡满等,2017)对各年龄阶段的油丹实际数量进行匀滑修正。
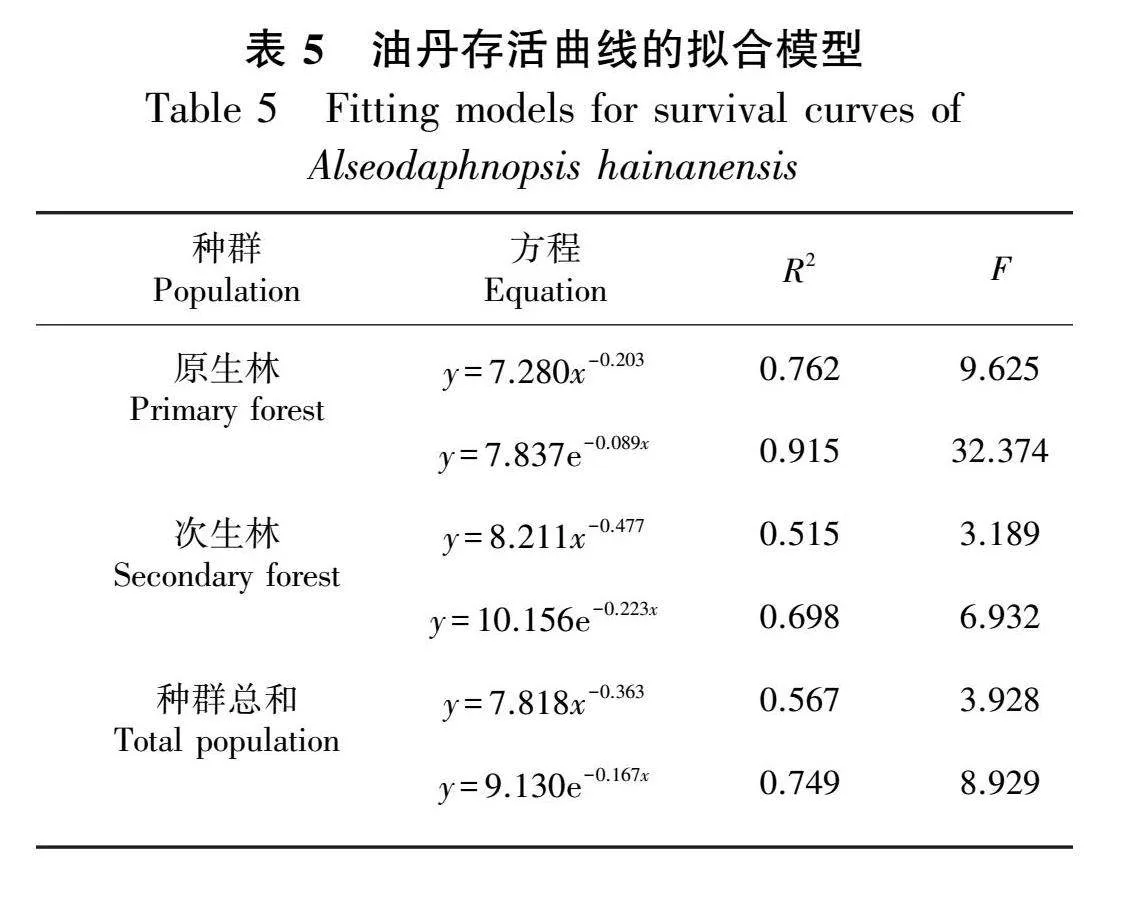
1.2.5 种群存活曲线 存活曲线为生命表数据提供了一种视觉表现形式,当无法准确获知特定年龄或年龄组的出生和死亡数据时,通过分析存活曲线的形态可以推断种群的增长和衰退模式(刘智慧,1990)。此研究通过将龄级置于横轴,将标准化存活数的自然对数(lnlx)置于纵轴,绘制了种群总体、原生林以及次生林的存活曲线。本研究中油丹存活曲线的分析采用Hett和Loucks(1976)的指数函数方程(y=y0e-bx)和幂函数方程(y=y0x-b)。
1.2.6 种群数量动态的时间序列预测 利用时间序列模型(张文辉,1998;谭一波等,2010;解婷婷等,2014)对经历未来2个、3个、4个、5个龄级时间后的油丹种群龄级结构进行模拟和预测。
2 结果与分析
2.1 油丹种群的龄级结构
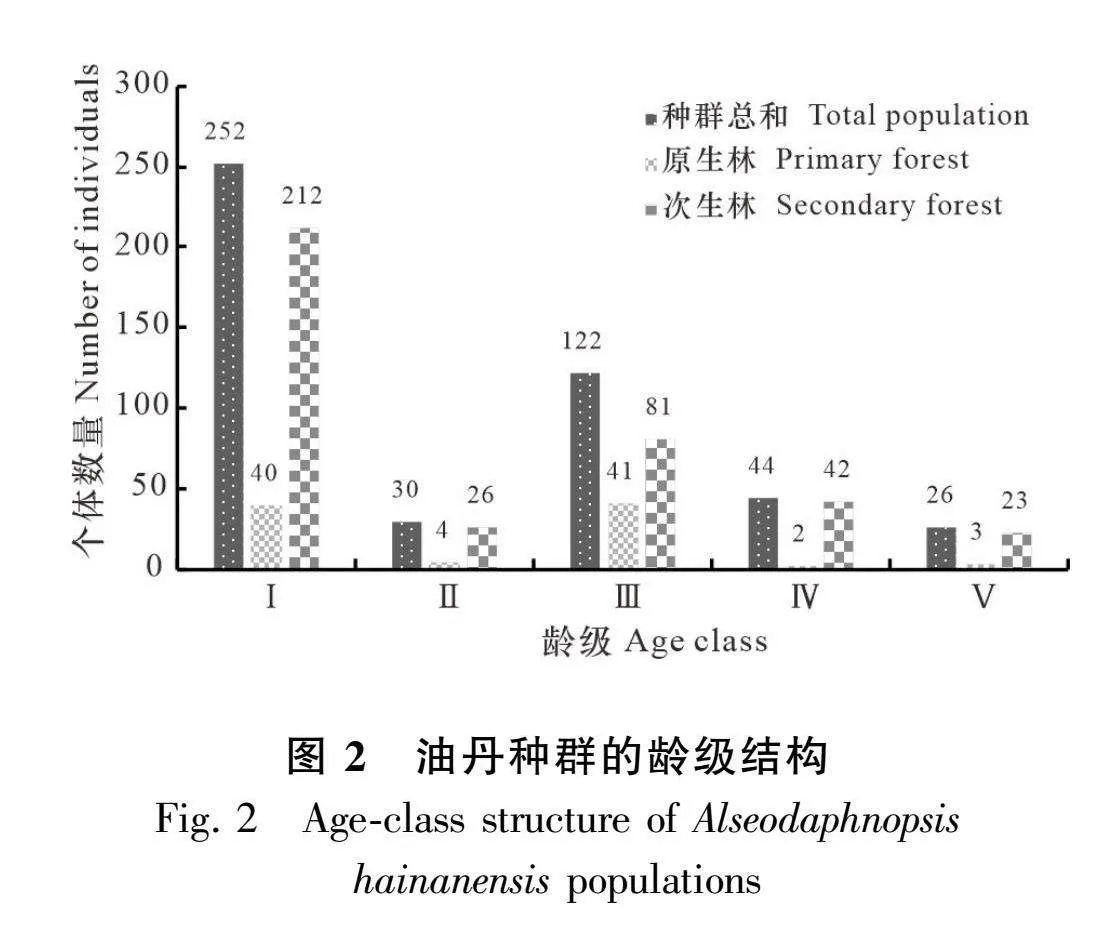
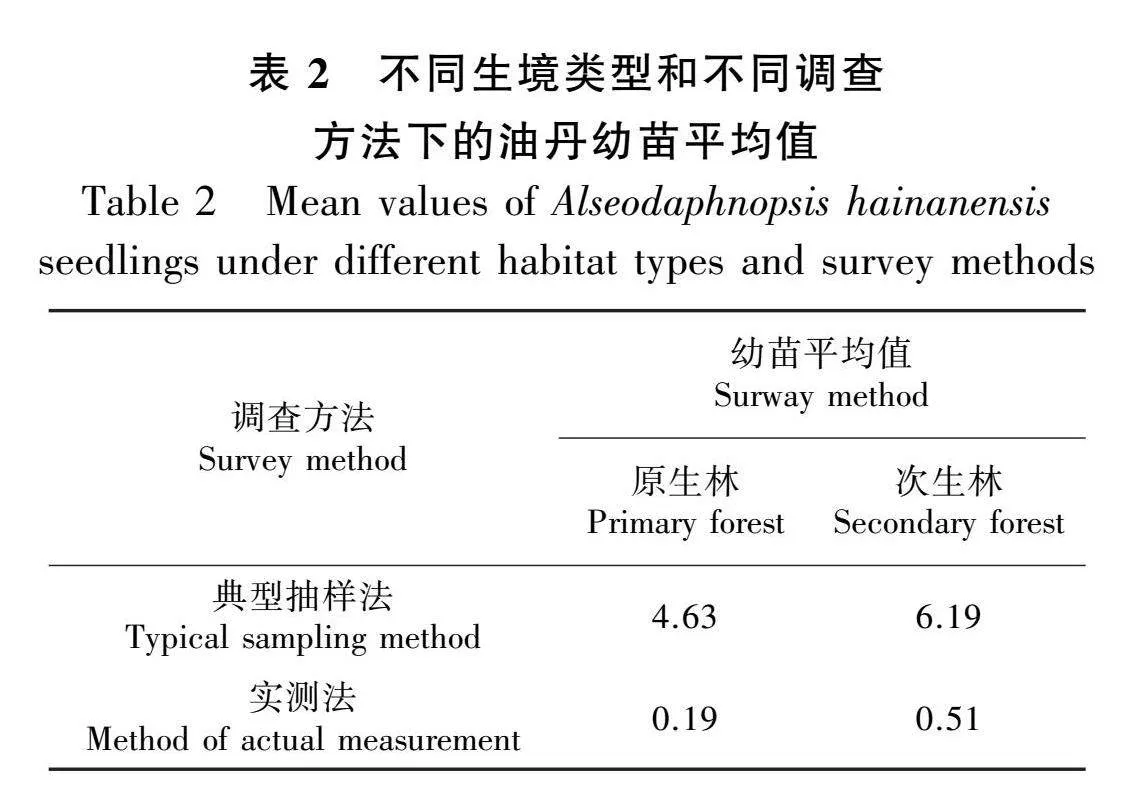
由图2可知,本次调查共统计到油丹个体474株。其中,原生林90株,最大胸径为71 cm;次生林384株,最大胸径为87 cm。由图2可知,原生林和次生林中油丹种群各龄级均有个体分布。整体来看,油丹种群龄级结构呈倒“J”型,为增长型种群。在原生林和次生林中,油丹种群结构均呈现出倒“J”形,然而两种不同生境中各个年龄段的植株数量存在差异,尤其是幼苗,次生林的幼苗数量超过原生林幼苗数量的5倍。在相同的调查方法下,次生林的幼苗平均数量高于原生林。在典型抽样调查方法下,原生林幼苗平均值为4.63,次生林幼苗平均值为6.19,说明次生林幼苗数量大于原生林;通过实测法调查,原生林幼苗平均值为0.19,次生林幼苗平均值为0.51,也得出相同的结论,说明次生林幼苗数量更多(表2)。
2.2 油丹种群的龄级结构动态
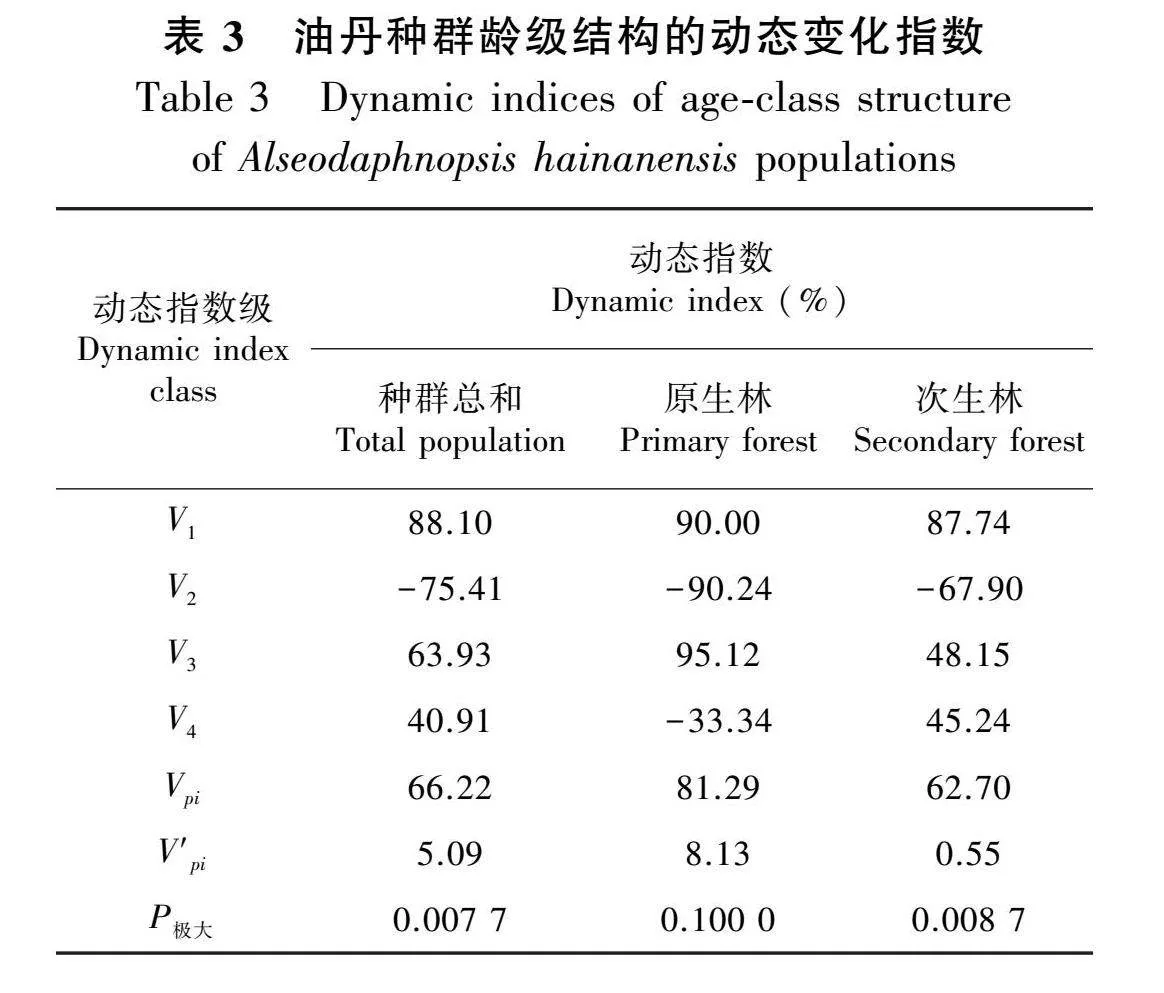
由表3可知,原生林油丹种群中,V1、V3大于0,表明Ⅰ-Ⅱ、Ⅲ-Ⅳ龄级的结构动态关系具有增长趋势;V2、V4小于0,说明Ⅱ-Ⅲ、Ⅳ-Ⅴ龄级的结构动态关系趋于衰退。次生林油丹种群中,V2小于0,说明Ⅱ-Ⅲ龄级的结构动态关系趋于衰退,其余龄级均为增长关系。在没有外部干扰因素的前提下,原生林和次生林中油丹种群的总体龄级结构动态指数Vpi均显示出正值,这表明油丹种群目前正处于增长状态,该结论与种群龄级结构的分析结论基本一致。即使在考虑到外部因素可能造成的影响后,原生林和次生林中油丹种群的总体龄级结构动态指数V′pi仍然保持正值,这一数据反映出该种群维持着增长趋势;考虑外部干扰时次生林的数值急剧下降且小于原生林。种群总和的随机干扰风险极大值P极大为0.007 7,原生林的随机干扰风险极大值P极大为0.100 0,次生林的随机干扰风险极大值P极大为0.008 7,说明不同生境中的油丹种群对外界环境干扰均具有一定的敏感性,种群抗干扰能力差。次生林的抗干扰概率最小,这说明次生林油丹种群的抗干扰能力较好。
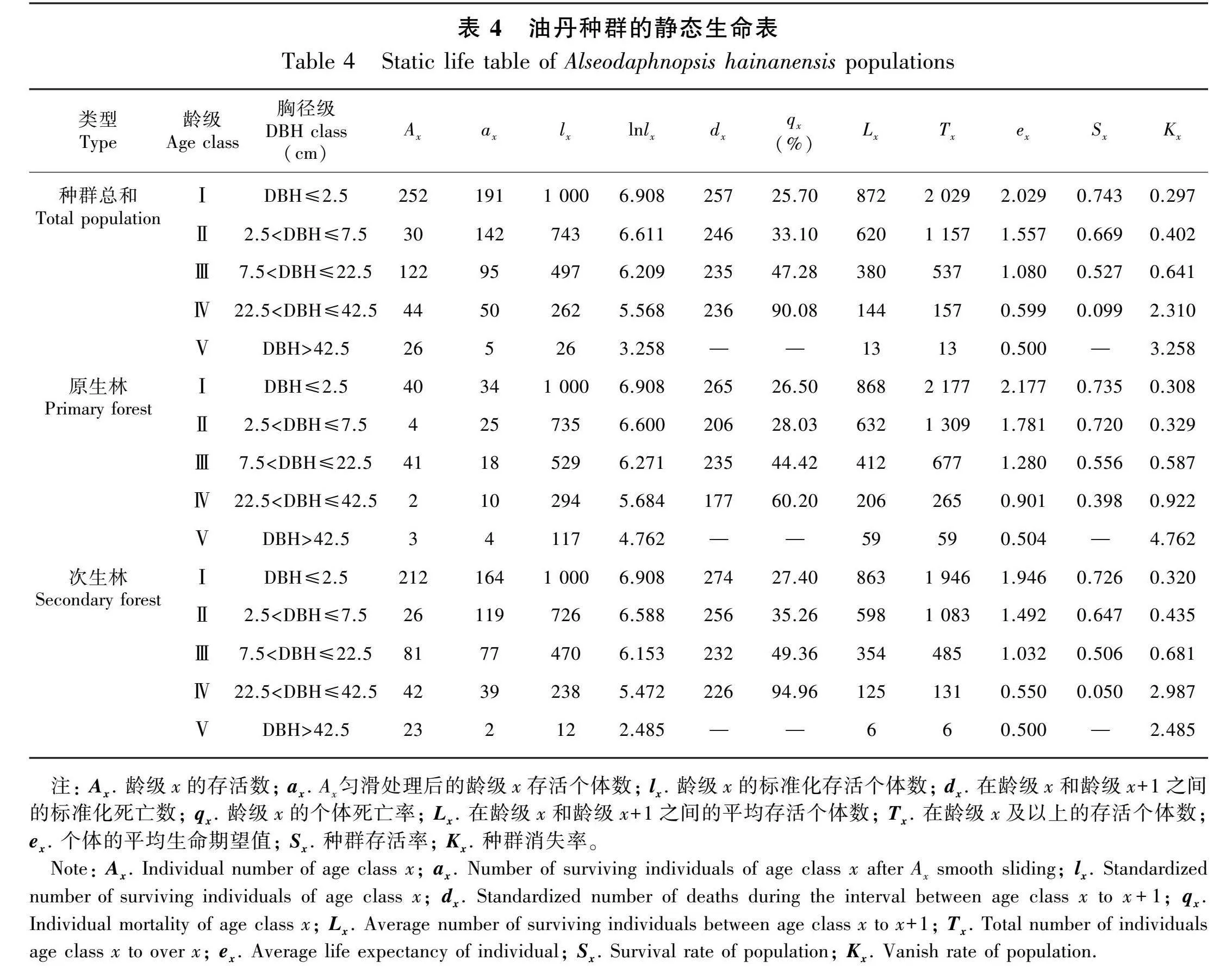
2.3 种群静态生命表
从静态生命表(表4)可以看出,原生林和次生林中油丹种群的标准化存活个体数(lx)随龄级的增长而降低,这与种群逐渐接近生理寿命有关,此趋势基本符合种群的生物学特征。原生林和次生林中油丹种群的生命期望值(ex)随龄级的增长逐渐减小,其中原生林和次生林中的油丹种群都是在第Ⅰ龄级和第Ⅱ龄级阶段ex较高,说明在幼苗和幼树阶段,个体对环境的适应能力较强。原生林和次生林中油丹种群的死亡率(qx)也表现为相似的趋势,随着龄级的增大,qx逐渐增大,次生林的油丹种群qx在第Ⅳ龄级较大,为94.96%,此龄级的消失率(Kx)也达到最大值,推测该龄级的个体可能遭到了砍伐或破坏。
2.4 种群的存活曲线
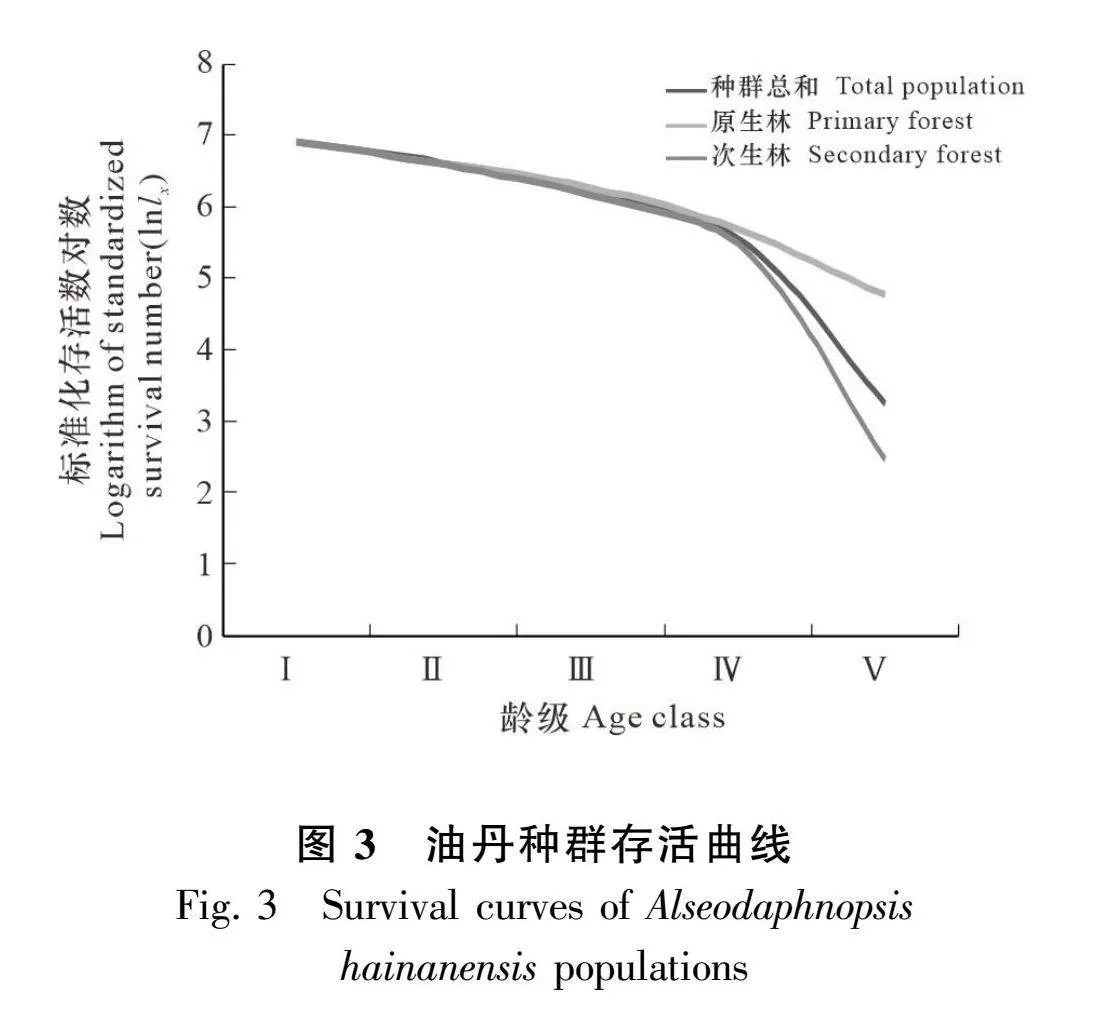
由图3可知,在原生林和次生林中,油丹种群的存活曲线表现出一致性和差异性。一致性方面,两种生境中的种群在最初的3个龄级(Ⅰ-Ⅲ)中,其标准化存活数对数(lnlx)的下降趋势大致保持一致。然而,在第四和第五龄级(Ⅳ-Ⅴ)期间,两种生境中的lnlx下降速度出现明显差异,即原生林的下降速度相对较慢,而次生林的下降速度则较为迅速。
原生林、次生林、种群总和的幂函数,检验方程和指数函数检验方程见表5,上述方程中,指数函数的R2值和F值均超过了幂函数的相应值,这表明油丹种群的存活曲线与Deevey-Ⅱ型的吻合度更高。
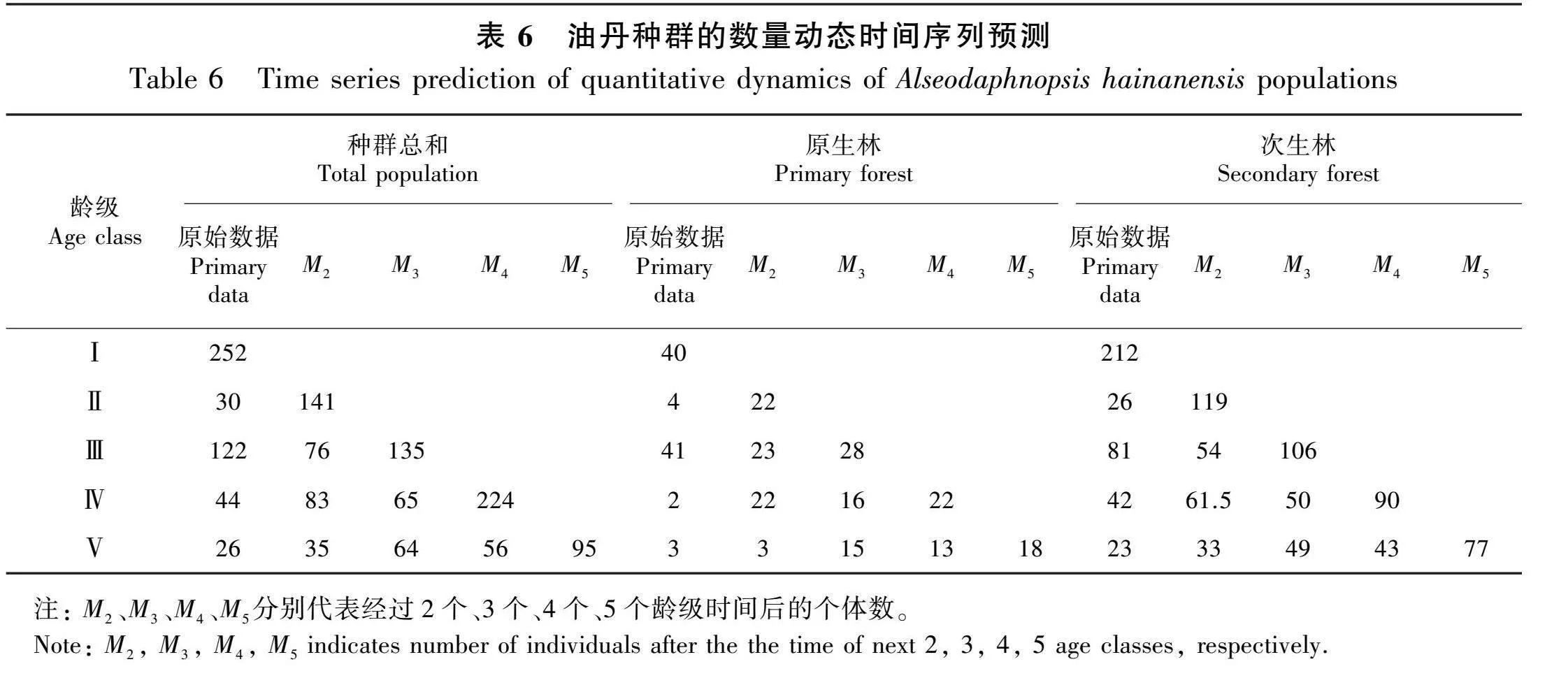
2.5 种群的时间序列预测分析
由表6可知,油丹种群总和中Ⅲ龄级的个体数经历2个龄级时间后出现下滑,其他各龄级在经历不同的龄级时间后个体数均有不同幅度的增加。在经历2~3个龄级时间后,原生林种群中Ⅲ龄级的个体数出现下滑,由原来的41株下降到23株和28株。次生林Ⅲ龄级的个体数在经历2个龄级后也出现下滑,由原来的81株减少到了54株。总体而言,随着时间的推移,油丹种群中的低龄级个体能够发育为成年个体,补充和替代中老龄级个体。
3 讨论
3.1 油丹种群的龄级结构
研究表明,原生林和次生林中的油丹种群龄级结构基本呈倒“J”型,油丹种群在特定的龄级遇到了生长瓶颈,但总体龄级结构动态指数均保持正值,这反映出该种群整体上呈现出增长的态势。本研究共记录到油丹474株,其中Ⅰ龄级的幼苗有252株,占比53.16%,可见幼苗数量储备充足;两种生境中油丹幼苗数量均较多,但在油丹幼苗转变为幼树的过程中,幼苗大量死亡,只有少部分幼苗能成功过渡到幼树阶段。野外调查过程中发现,霸王岭、尖峰岭、五指山、吊罗山、黎母山的油丹种群都面临一样的困境,幼苗经常性更新失败。罗文等(2023)研究了尖峰岭两种生境中坡垒的种群结构,发现种群的龄级结构基本呈倒“J”型,但在由幼苗转变为小树的过程中同样存在增补限制问题,并且这种增补限制未随研究地点而发生改变,这与本研究结果具有一致性。此外,油丹次生林比原生林保存有更多成年个体,幼苗数量也更多,这一现象的成因可能与油丹自身的生物学特性以及生境的环境差异有关。一方面,陶建平和臧润国(2004)研究发现油丹属于阳性树种,幼苗生长对光照要求苛刻,往往幼苗数量多,但死亡率也高,这与本研究结果具有一致性。另一方面,油丹种群次生林的郁闭度比原生林低,次生林郁闭度平均值为0.62,原生林郁闭度平均值为0.69,次生林林下光照强度更高。综上所述,推测次生林的光照条件更适合油丹幼苗生长。赵家豪等(2022)研究发现原始林林下的高郁闭现状无法得到改善,不利于南方铁杉生长;王燕玲等(2023)研究发现林下光照不足是阻碍红鳞蒲桃幼苗种群天然更新的因素之一,可见适当的光照有利于阳性树种幼苗的生长。
3.2 油丹种群的动态特征
本研究表明,原生林和次生林中油丹种群的存活曲线趋近于Deevey-Ⅱ型,在整个生长阶段各龄级的死亡率较为稳定,与极小种群植物喜树(张兴旺等,2023)、极小种群植物海南风吹楠(蒋迎红等,2017)、珍稀濒危植物珙桐(徐云飞等,2020)的存活曲线类型相似。
赵阳等(2018)和项小燕等(2016)提出限制种群更新的关键因子主要是有限的资源和生长空间。在森林更新过程中,幼苗和幼树阶段是最为敏感和重要的阶段(张婕等,2014)。野外观察发现,油丹的种子比较大,落地后很难直接入土,长时间停留在地面,容易发霉及被动物取食。在重力的作用下,种子垂直落下,仅分布在母树附近,这就导致幼苗期竞争激烈,淘汰了大量幼龄个体,导致整体种群较小。研究结果表明,次生林的幼苗数量相对原生林充足,尽管次生林存在一定数量的油丹种群,但只有极少数的幼苗可以转化为幼树,导致维持油丹种群稳定性的基础不牢固。长期低下的幼苗至幼树转化率可能会导致油丹种群的增长趋势逆转,转变为衰退型种群。前人研究表明,大量的低龄幼苗是各龄级个体数得以增加的基础,低龄幼苗比例越高种群的增长潜力越大(张晓鹏等,2022)。因此需要采取合适的措施,增加油丹幼苗的成活率,保证种群稳定增长。
从时间序列预测结果可看出,不同龄级个体数的增长速率在原生林和次生林中存在差异。在经历5个龄级时间之后,原生林和次生林中的油丹个体数量都逐步增加,这说明原生林和次生林中的油丹种群均具备较好的恢复潜能,只要给予适当的时间,都能恢复至稳定状态。与本研究类似,罗文等(2023)对海南尖峰岭的坡垒种群进行了预测,发现在保持尖峰岭现有坡垒种群和生境不被破坏的前提下,原始林和次生林的坡垒种群数量在经历2~5个龄级的时间后均将具备较好的恢复潜能。张晓鹏等(2022)对辽东山区的红松幼苗种群进行研究,发现随着龄级的增加,该种群能够实现天然更新,但存在更新障碍。
3.3 油丹种群的濒危原因及保护建议
大多数濒危植物的濒危原因可通过内外两个因素共同解释。内在因素包括遗传力、生殖力、生活力、适应力的衰竭,这些原因威胁植物生长繁衍致使其稀有濒危;外在因素包括自然因素和人为因素(吴小巧等,2005;Xu amp; Zang,2023)。本研究主要从内在因素探讨油丹的濒危原因。Zhang等(2024)在海南岛收集了蕉木的35个样本,研究发现栖息地破碎化不仅会减少种群规模,还会增加空间距离,导致基因流和遗传多样性的减少。油丹在海南的分布也呈现出破碎化格局,结合上述观察及前人的研究,推测油丹种群遗传多样性低,影响了其适应环境变化的能力,加剧了灭绝的风险,这有待后续研究验证。结合上述分析,油丹濒危的主要原因如下:(1)种子繁殖成功率低;(2)竞争能力弱,幼龄个体死亡率高;(3)种群分布范围小,可能导致遗传多样性低。鉴于本研究揭示的油丹种群存活状况,保护工作应聚焦于以下几个领域:(1)完善对现有油丹种群的保护管理工作,加强现有种群的就地保护和天然更新监测。对于油丹成年个体分布集中的区域,应加强监管,禁止砍伐树木,使油丹群落生境得到有效保护;对于幼苗丰富的区域可以采取适当的上层林冠修枝措施,降低郁闭度,促进其自然更新和再生。(2)加大幼苗繁育研究及迁地栽培技术,利用培育的幼苗开展野外回归研究,从而促进油丹种群恢复,扩大油丹种群数量及分布面积。(3)开展种群遗传多样性研究,深入理解其致濒机理,研究油丹遗传组成分布、揭示其响应环境变化的遗传机制,为制定相应的保护措施提供科学依据。
4 结论
本研究发现,原生林和次生林中的油丹种群结构均呈倒“J”型,各龄级均有个体分布;种群均属于增长型,抗干扰能力较差;存活曲线接近于Deevey-Ⅱ型,在整个生长阶段,各龄级的死亡率基本一致;在经历2~5个龄级时间后,两种生境中的油丹种群均能够实现自然更新,但低龄个体在更新过程中遭遇的障碍可能会使种群未来面临衰退的风险。建议加强就地保护、人工扩繁和迁地保护等研究,同时采取适度的人为措施,提高幼苗到幼树的转化率,促进油丹种群数量增长。
参考文献:
CHANG HT, LIU L, TU PF, 2000. Studies on the chemical constituents of Alseodaphne hainanensis [J]. Chinese Traditional and Herbal Drugs, 31(10): 7-9. "[常海涛, 刘潋, 屠鹏飞, 2000. 油丹化学成分的研究 [J]. 中草药, 31(10): 7-9.]
CHEN XD, 1998. A study on the method of quantitative analysis for plant population and community structural dynamics [J]. Acta Ecologica Sinica, 18(2): 104-107. "[陈晓德, 1988. 植物种群与群落结构动态量化分析方法研究 [J]. 生态学报, 18(2): 104-107.]
CHEN YK, YANG XB, LI DH, et al., 2011. Interspecific associations among dominant populations in the communities with Alseodaphne hainanensis in Bawangling, Hainan Island [J]. Journal of Tropical and Subtropical Botany, 19(3): 237-244. "[陈玉凯, 杨小波, 李东海, 等, 2011. 海南霸王岭油丹所在群落的优势种群种间联结性研究 [J]. 热带亚热带植物学报, 19(3): 237-244.]
FAN HL, HONG W, WU CZ, et al., 2004. Population structure and dynamics of dominant species in Castanopsis fargesii secondary forest in Datian County, Fujian Province [J]. Journal of Plant Resources and Environment, 13(3): 44-49. "[范海兰, 洪伟, 吴承祯, 等, 2004. 福建大田栲树次生林优势种群结构与动态 [J]. 植物资源与环境学报, 13(3): 44-49.]
FROST I, RYDIN H, 2000. Spatial pattern and size distribution of the animal-dispersed tree Quercus robur in two spruce-dominated forests [J]. Ecoscience, 7(1): 38-44.
GONG ZT, ZHANG GL, QI ZP, 2004. Introduction to the soil system of Hainan Island [M]. Beijing: Science Press. "[龚子同, 张甘霖, 漆智平, 2004. 海南岛土系概论 [M]. 北京:科学出版社.]
GOU ZH, HUANG CT, LIN L, et al., 2017. Effects of four hormones on seed germination of Alseodaphne hainanensis [J]. Tropical Forestry, 45(1): 4-6. "[苟志辉, 黄川腾, 林玲, 等, 2017. 4种激素对油丹种子发芽的影响 [J]. 热带林业, 45(1): 4-6.]
HE JN, NING CW, ZHANG WT, et al., 2024. Altitudinal patterns of population structure dynamics of Picea schrenkiana in the northern slope of the eastern Tianshan Mountains [J]. Acta Ecologica Sinica, 44(12): 5389-5398. "[何佳宁, 宁才文, 张闻涛, 等, 2024. 东天山北坡雪岭云杉种群结构动态的海拔格局 [J]. 生态学报, 44(12): 5389-5398.
HETT JM, LOUCKS O L, 1976. Age structure models of balsam fir and eastern hemlock [J]. The Journal of Ecology, 64(3): 1029-1044.
HU QH, CHEN L, LAI Y, et al., 2023. Population structure and dynamic characteristics of endangered plant Amentotaxus argotaenia in Guanshan Mountain of Jiangxi Province [J]. Acta Agriculturae Universitatis Jiangxiensis, 45(6): 1418-1430. "[胡千惠, 陈琳, 赖媛, 等, 2023. 江西官山濒危植物穗花杉种群结构与动态 [J]. 江西农业大学学报, 45(6): 1418-1430.]
HU M, ZENG SQ, LONG SS, et al., 2017. Structure and dynamic characteristics of Cyclobalanopsis glauca secondary forests [J]. Journal of Central South University of Forestry amp; Technology, 37(11): 110-114. "[胡满, 曾思齐, 龙时胜, 等, 2017. 青冈栎次生林种群结构及动态特征 [J]. 中南林业科技大学学报, 37(11): 110-114.]
HUANG GH, LIANG KN, ZHOU ZZ, 2011. Research and preservation of "Madhuca hainanensis and Alseodaphne hainanensis [J]. Guangdong Forestry Science and Technology, 27(5): 67-71. "[黄桂华, 梁坤南, 周再知, 2011. 子京和油丹的研究与保护 [J]. 广东林业科技, 27(5): 67-71.]
JIANG H, 1992. Population ecology of Picea asperata [M]. Beijing: China Forestry Publishing House. "[江洪, 1992. 云杉种群生态学 [M]. 北京: 中国林业出版社.]
JIANG YH, XIANG WH, HE YH, et al., 2017. Population quantitative characteristics and dynamics of Horsfieldia hainanensis, a rare and extremely small population plant [J]. Journal of Central South University of Forestry amp; Technology, 37(8): 66-71. "[蒋迎红, 项文化, 何应会, 等, 2017. 极小种群海南风吹楠种群的数量特征及动态 [J]. 中南林业科技大学学报, 37(8): 66-71.]
LI ZC, CHEN YF, HONG XJ, et al., 2015. Age structure and point pattern analysis of Dacrydium pectinatum in Bawangling, Hainan Island [J]. Chinese Journal of Ecology, 34(6): 1507-1515. "[李肇晨, 陈永富, 洪小江, 等, 2015. 海南霸王岭陆均松种群年龄结构与点格局分布特征 [J]. 生态学杂志, 34(6): 1507-1515.]
LIU SJ, ZHOU GS, FANG SB, 2015. Climatic suitability regionalization of rubber plantation in China [J]. Scientia Agricultura Sinica, 48(12): 2335-2345. "[刘少军, 周广胜, 房世波, 2015. 中国橡胶树种植气候适宜性区划 [J]. 中国农业科学, 48(12): 2335-2345.]
LIU ZH, 1990. A primary study on structure and dynamics of Castanopsis fargesii population on Jinyun Mountain, Sichuan Province [J]. Journal of Plant Ecology and Geobotany, 14(2): 120-128. "[刘智慧, 1990. 四川省缙云山栲树种群结构和动态的初步研究 [J]. 植物生态学与地植物学学报, 14(2): 120-128.]
LUO W, XU H, LI YP, et al., 2023. Population structure and quantitative dynamics of a wild plant with extremely small populations Hopea hainanensis [J]. Forest Research, 36(2): 169-177. "[罗文, 许涵, 李艳朋, 等, 2023. 极小种群野生植物坡垒的种群结构与数量动态研究 [J]. 林业科学研究, 36(2): 169-177.]
OMELKO A, UKHVATKINA O, ZHMERENETSKY A, et al., 2018. From young to adult trees: How spatial patterns of plants with different life strategies change during age development in an old-growth Korean pine-broadleaved forest "[J]. Forest Ecology and Management, 411: 46-66.
SHEN SK, MA HY, WANG YH, et al., 2008. The structure and dynamics of natural population of the endangered plant Euryodendron excelsum H. T. Chang [J].Acta Ecologica Sinica, 28(5): 2404-2412. "[申仕康, 马海英, 王跃华, 等, 2008. 濒危植物猪血木自然种群结构及动态 [J]. 生态学报, 28(5): 2404-2412.]
TAN YB, ZHAN CA, XIAO ZX, et al., 2010. Population structure and dynamic characteristics of Machilus chinensis in Nan’ao Island, Guangdong Province [J]. Chinese Journal Ecology, 29(10): 1901-1906. "[谭一波, 詹潮安, 肖泽鑫, 等, 2010. 广东南澳岛中华楠种群结构及动态特征 [J]. 生态学杂志, 29(10): 1901-1906.]
TAO JP, ZANG RG, 2004. Studies on the dynamics of seedling bank in gap of tropical montane rain forest in Bawangling Hainan Island [J]. Scientia Silvae Sinicae, 40(3): 33-38. "[陶建平, 臧润国, 2004. 海南霸王岭热带山地雨林林隙幼苗库动态规律研究 [J]. 林业科学, 40(3): 33-38.]
WANG YL, ZHAO LJ, ZHU LQ, et al., 2023. Seedling quantitative characteristics and dynamics of Syzygium hancei populations in Guangxi, China [J]. Chinese Journal of Plant Ecology, 47(9): 1278-1286. "[王燕玲, 招礼军, 朱栗琼, 等, 2023. 广西天然红鳞蒲桃种群幼苗数量特征及动态分析 [J]. 植物生态学报, 47(9): 1278-1286.]
WANG YT, HUANG ZH, WANG J, et al., 2021. The population structure and dynamic characteristics of Phellodendron amurense in Yanshan Mountains "[J]. Acta Ecologica Sinica, 41( 7): 2826-2834. [王泳腾, 黄治昊, 王俊, 等, 2021. 燕山山脉黄檗种群结构与动态特征 [J]. 生态学报, 41(7): 2826-2834.]
WU XQ, 2005. Research on the protection of woody rare and endangered plants and their guarantee mechanism in Jiangsu Province [D]. Nanjing: Nanjing Forestry University. "[吴小巧, 2005. 江苏省木本珍稀濒危植物保护及其保障机制研究 [D]. 南京: 南京林业大学.]
XIANG XY, WU GL, DUAN RY, et al., 2016. Studies on population structure and dynamics of Pinus dabeshanensis [J]. Resources and Environment in the Yangtze Basin, 25(1): 55-62. "[项小燕, 吴甘霖, 段仁燕, 等, 2016. 大别山五针松种群结构及动态研究 [J]. 长江流域资源与环境, 25(1): 55-62.]
XIE TT, SU PX, ZHOU ZJ, et al., 2014. Structure and dynamic characteristics of Calligonum mongolicum population in the desert-oasis ecotone [J]. Acta Ecologica Sinica, 34(15): 4272-4279. "[解婷婷, 苏培玺, 周紫鹃, 等, 2014. 荒漠绿洲过渡带沙拐枣种群结构及动态特征 [J]. 生态学报, 34(15): 4272-4279.]
XU Y, ZANG RG, 2023. Conservation of rare and endangered plant species in China [J]. Iscience, 26(2): 106008.
XU YF, LIU QS, XU WJ, et al., 2020. Differences in population structure and dynamic characteristics of Davidia involucrata Baill. between high and low latitude regions [J]. Bulletin of Botanical Research, 40(6): 855-866. "[徐云飞, 刘沁松, 徐文娟, 等, 2020. 天然珙桐种群结构与动态特征在高低纬度地区的差异 [J]. 植物研究, 40(6): 855-866.]
ZHANG L, ZHANG HL, CHEN YK, et al., 2024. Assessing genetic diversity in critically endangered Chieniodendron hainanense populations within fragmented habitats in Hainan [J]. Scientific Reports, 14(1): 6988.
ZHANG J, SHANGGUAN TL, DUAN YH, et al., 2014. Age structure and dynamics of Quercus wutaishanica population in Lingkong Mountain of Shanxi Province, China "[J]. Chinese Journal of Applied Ecology, 25(11): 3125-3130. "[张婕, 上官铁梁, 段毅豪, 等, 2014. 灵空山辽东栎种群年龄结构与动态 [J]. 应用生态学报, 25(11): 3125-3130.]
ZHANG WH, 1998. Population ecology of Adenophora lobophylla [M]. Harbin: Northeast Forestry University Publishing House: 107-114. "[张文辉, 1998. 裂叶沙参种群生态学研究 [M]. 哈尔滨: 东北林业大学出版社: 107-114.]
ZHANG XP, YU LZ, YANG XY, et al., 2022.Population structure and dynamics of Pinus koraiensis seedlings regenerated from seeds in a montane region of eastern Liaoning Province, China [J]. Chinese Journal of Applied Ecology, 33( 2): 289-296. "[张晓鹏, 于立忠, 杨晓燕, 等, 2022. 辽东山区天然更新红松幼苗种群结构与动态 [J]. 应用生态学报, 33(2): 289-296.]
ZHANG XW, XIE YP,WU XM, et al., 2023. Population structure and dynamic characteristics of wild plant species with extremely small populations of Camptotheca acuminata in Mingxi, Fujian Province, China [J]. Ecology and Environmental Sciences, 32(6): 1037-1044. "[张兴旺, 谢艳萍, 吴晓敏, 等, 2023. 福建省明溪县极小种群野生植物喜树种群结构与动态特征 [J]. 生态环境学报, 32(6): 1037-1044.]
ZHANG Y, YI XM, WANG YX, et al., 2015. Impact of tree harvesting on the population structure and dynamics of Pinus koraiensis (Pinaceae) [J]. Acta Ecologica Sinica, 35(1): 38-45. "[张悦, 易雪梅, 王远遐, 等, 2015. 采伐对红松种群结构与动态的影响 [J]. 生态学报, 35(1): 38-45.]
ZHAO JH, YE YQ, SUN XD, et al., 2022. Population dynamics and spatial distribution of the rate and endangered plant Tsuga chinensis var. tchekiangensis in Wuyishan, Jiangxi Province [J]. Acta Ecologica Sinica, 42(10): 4032-4040. "[赵家豪, 叶钰倩, 孙晓丹, 等, 2022. 江西武夷山珍稀濒危植物南方铁杉种群动态与空间分布 [J]. 生态学报, 42(10): 4032-4040.]
ZHAO Y, QI R, JIAO J, et al., 2018. The population structure and dynamic characteristics of Picea purpurea at the Gahai-Zecha area [J]. Acta Ecologica Sinica, 38(20): 7447-7457. "[赵阳, 齐瑞, 焦健, 等, 2018. 尕海-则岔地区紫果云杉种群结构与动态特征 [J]. 生态学报, 38(20): 7447-7457.]
(责任编辑 周翠鸣)
猜你喜欢
安徽农业科学(2021年15期)2021-09-03 03:03:03
湖北林业科技(2021年1期)2021-07-25 16:22:37
热带作物学报(2020年9期)2020-10-29 07:35:39
热带作物学报(2019年11期)2019-12-19 02:13:14
安徽农学通报(2019年11期)2019-07-12 07:53:51
植物保护(2019年3期)2019-06-27 02:07:05
绿色科技(2018年11期)2018-03-04 08:43:00
绿色科技(2016年23期)2017-03-15 15:41:56
科教导刊·电子版(2016年25期)2016-11-16 10:24:35
安徽农业科学(2016年4期)2016-10-21 16:47:53
