
北京市紫椴种群年龄结构及动态研究
2017-03-15 15:41许梅沐先运
绿色科技 2016年23期
关键词:北京市
许梅+沐先运


摘要:在玉渡山和雾灵山共选择了12个样方,探讨了北京市紫椴种群的年龄结构及未来发展规律,结果表明:该种群年龄结构和种群结构动态量化结果均为增长型;种群存活曲线趋于DeeveyⅡ型,未来一定时间内,在没有外部干扰的情况下,种群将保持相对稳定的种群数量并逐渐扩大。
关键词:北京市;紫椴;径级;静态生命表;存活曲线;时间序列预测
中图分类号:Q948.1
文献标识码:A 文章编号:1674-9944(2016)23-0001-05
1 引 言
种群结构是种群最基本的特征,不仅能反映种群动态及其发展趋势,在一定程度上也反映了种群与环境间的相关关系以及它们在群落中的地位。植物种群数量动态是植物个体生存能力与外界环境相互作用的结果[1]。研究不同生境条件下植物种群年龄结构、生命表以及存活曲线,不仅可以反映种群现实状况,还可以展现植物种群与环境抗争的关系,尤其对于濒危植物的保护和利用研究具有重要意义[2]。
紫椴(Tilia amurensis)是椴树科椴树属的落叶阔叶乔木,是东北植被区阔叶红松混交林群落的主要伴生种及人工天然复合群落的优良混交组分,为国家Ⅱ级保护植物[3]。作为东北地区重要的造林树种和蜜源植物,其生态价值和经济价值都十分显著。由于长期以来的过量采伐和利用,野生紫椴种群的数量和质量逐年下降,蓄积大幅减少[4]。因此,开展紫椴种群年龄结构及其动态特征的研究,对于紫椴种群的保护意义重大。
近年来,越来越多的学者致力于对紫椴的研究,涉及育种造林[5]、无性繁殖技术[6]、生长环境[7~9]、种子解剖[10]、种子休眠[11]、种子雨和种子库[12,13]、遗传变异[14,15]、形态解剖[16~18]、种群空间格局[4,19]等各方面。而对于紫椴种群年龄结构的研究,除在聂绍荃[20]的紫椴种群生态学研究中有涉及外,尚未见有新的报道。本研究通过静态生命表、存活曲线以及时间序列预测模型等分析种群数量动态、发展趋势及其在群落中的地位,探索其生态濒危机制,为紫椴的深入研究奠定基础和提供科学依据,以期尽早提出对北京市紫椴种群适应性保护和恢复对策。
2 研究区概况
本研究取样范围在40°33′15″~40°34′20″ N, 115°51′39″~117°24′33″ E,海拔932~1240 m,研究区位于北京市玉渡山、雾灵山地区。玉渡山为国家级生态保护区,地处北京延庆县城西北,南沿后河、古城河河谷,西起海拔2241 m的北京市第二高峰海坨主峰,东至龙庆峡,境内海拔580~1589 m。该地区植物生长茂盛,种类繁多,乔木主要由松、桦、山杨、辽东栎、椴树和北京丁香为主的杂木林组成;灌木层主要由山榆、二色胡枝子、平榛、虎榛、绣线菊、鼠李、山桃、荆条、山杏等组成。雾灵山是燕山山脉主峰,海拔2118 m,年平均气温7.6 ℃,年降水量1000 mm,位于華北、东北、蒙古三大植物区系交汇之处,植物垂直带谱明显,植被类型多样,许多古老孑遗植物保存良好,森林覆盖率93%,有高等植物1870种,其中中国珍稀濒危保护植物10种。
3 研究方法
3.1 样方调查
2015年7~8月,在全面了解紫椴自然分布的基础上,据对北京市紫椴分布基状况的了解和实地踏查,在玉渡山和雾灵山共选择了12个20 m×20 m的样方进行调查,记录各样方的群落类型、海拔、坡度、坡向、郁闭度等生境因子以及GPS坐标(表1)。并对每个样方内的乔木进行每木调查,记录胸径大于等于4 cm的全部乔木的种名、树高、胸径、冠幅等,并记录所有胸径小于4 cm的紫椴幼苗的树高、基径和冠幅。
3.2 数据处理与分析
3.2.1 种群年龄结构分析方法
种群年龄结构是种群的重要特征,是分析探索种群动态的有效方法[21]。紫椴作为国家Ⅱ级保护物种,不宜采取用生长锥获取年轮数据的方法,但由于紫椴相关解析木资料缺乏,故采用大小结构分析法,用立木级结构代替种群年龄结构来分析种群动态。许多学者对种群大小结构立木级的划分提出了不同的标准,本文根据北京市紫椴种群生长状况,并参考聂绍荃[20]和董点[22]等对紫椴种群年龄阶段的划分方法,根据胸径(DBH)大小将紫椴划分为4个年龄阶段:幼树阶段(0 cm
此外,将紫椴依胸径大小分级,每级间隔3 cm,即0~3 cm为第1径级,3~6 cm为第2径级,6~9 cm为第3径级……,把树木径级从小到大的顺序看作是时间顺序关系,第1径级对应第1龄级,第2径级对应第2龄级,如此一一对应,统计各龄级株数,编制紫椴种群静态生命表,进而分析其动态变化。
3.2.2 种群结构动态量化
采用陈晓德[23]的量化方法定量描述北京市紫椴种群的动态。
3.2.3 种群静态生命表及存活曲线
以紫椴种群年龄结构为基础编制静态生命表,具体方法见文献[24,25]。存活曲线反映种群的动态特征,依据静态生命表中不同年龄的存活数绘制出种群的存活曲线,以龄级为横坐标,以标准化存活数(lx)和标准化死亡量(dx)为纵坐标,绘制北京市紫椴种群的存活曲线;以龄级为横坐标,死亡率(qx)为纵坐标,绘制种群的死亡率曲线。
3.2.4 时间序列预测
采用时间序列分析的一次移动平均法[21,26]对北京市紫椴种群龄级结构进行模拟和预测,其模型为:Mt=n[]1∑t[]k=t-n+1Xk,式中,n为需要预测的未来时间年限,t为龄级,是近期n个观测值在t时刻的平均值,称为第n周期的移动平均,即表示未来n年时t龄级的种群大小,Xt表示当前t龄级的种群大小。本文t分别取种群经历2、4、6和8个龄级的时间后,对种群未来发展趋势进行预测。
4 结果与分析
4.1 年龄结构及其类型
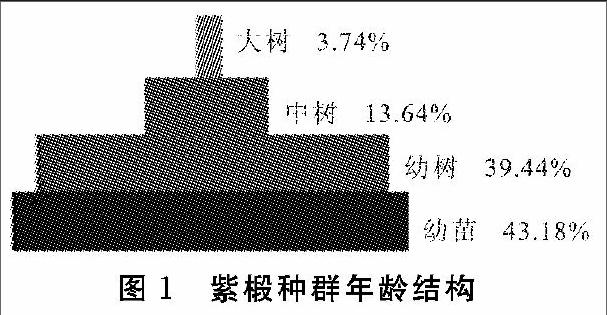
本研究中共记录到紫椴个体总数为614株,平均胸径为9.91 cm,最大胸径为35.3 cm。根据不同的年龄阶段,按照幼树、小树、中树、大树划分,绘制北京市紫椴种群的年龄结构图(图1)。可以看出:幼树所占的比例最大,为总体的44.79%,小树的数量次之。种群年龄结构呈正金字塔形,说明北京市紫椴种群为增长型。幼年个体与中老年个体之间的比例为83∶17,根据M.Alexander的划分方法,北京市紫椴种群具有高生产力。
按照划分的12个大小级绘制北京市紫椴种群的年龄结构图(图2),可知种群年龄结构整体呈左偏态分布,低中龄级个体较多,高龄级个体偏少。其中第2龄级个体数最多,占总数的19.54%;第1龄级个体数较少,表明紫椴幼苗缺乏;第3龄级个体少于其相邻龄级个体的数量,揭示第3龄级个体损失较大;第5龄级比上一级数量显著减少,说明环境筛选作用以及其他因素对第5龄级的紫椴的生存造成一定程度的压力;同理,第9龄级的紫椴个体也遭受了较大的损失。总的来说,该种群主体处于幼树和小树阶段,种群的维持依赖于中幼龄个体,种群发展较稳定。在对种群的抚育和管理中,一方面要着力提高幼苗的成活率,另一方面要减少老龄个体的死亡率,保证种群向增长型发展。
种群结构动态量化结果显示,各年龄级间个体数量变化动态指数Vn分别为:-36.67%、9.17%、-4.39%、36.84%、31.94%、38.78%、20.00%、62.50%、22.22%、57.14%、66.67%,其中V1小于0,表明林下幼苗缺少,种群天然更新较差;V3小于0,说明种群发展过程中存在一定的波动。整个种群年龄的动态指数为Vpi=0.149,在考虑外部干扰的情况下为Vpi'=0.012>0,表明种群在不考虑外部干扰时呈增长型;而在考虑外部干扰时仍有一定的增长性,但增长较为缓慢,这揭示出外部干扰对北京市紫椴种群的发展具有一定影响。
4.2 静态生命表分析
对于长寿命多年生植物种群,应用“空间差异代替时间变化”的方法来研究其种群动态具有实际意义,年龄结构和静态生命表以及种群动态预测都是基于现有调查数据,能够反映种群的基本属性,结果符合种群的自然动态规律[24,27]。静态生命表常是由“空间推导时间”构成,分析中常产生死亡率为负的情况。对此许多学者[25,27,28]采用匀滑技术对数据进行处理,以解决静态生命表中产生死亡率为负的问题,本文亦采用相同的方法,对北京市紫椴种群第1龄级、第4龄级的数据进行匀滑处理后(ax′),绘制静态生命表如表2。
由种群静态生命表可以看出:随着年龄的增加,北京市紫椴种群个体存活量lx逐渐减少;死亡量dx呈不规则的波动状态,这表明各个龄级的紫椴面临着不同的生存威胁,导致种群发展过程不稳定;死亡率qx和消失率Kx曲线的变化是一致的,总体上都表现上升趋势,但曲线皆呈波动状态:第6和第8龄级分别占据一个峰值,而第4、7、9龄级占据了三个谷值,这与死亡量曲线所显示的变化规律是相符的;另外,该种群的区间寿命Lx和总寿命Tx都逐渐缩短,且曲线都是内凹型;平均期望寿命ex总体也呈现逐渐减少的趨势,只是在第8龄级出现波动,其曲线可用y=0.0166x2-0.5419x+4.7938(R2=0.9742 R2=0.9771)描述。
4.3 生存曲线分析
以北京市紫椴种群静态生命表中标准化存活量lx为纵坐标,以龄级为横坐标绘制种群存活曲线(图3)。按Deevey的划分,种群存活曲线一般有3种基本类型:Ⅰ型是凸曲线,属于该型的种群绝大多数都是能活到该物种年龄,早期死亡率较低,但当活到一定生理年龄时,短期内几乎全部死亡;Ⅱ型是直线,也称对角线型,属于该型的种群各年龄的死亡率基本相同;Ⅲ型是凹曲线,早期死亡率高,一旦活到某一年龄,死亡率就较低。
由图3可以看出:北京市紫椴种群生长前期个体数较高,随年龄增加而下降,并趋于平稳。运用SPSS统计软件建立两种数学模型,进行北京市紫椴种群存活曲线类型的检验,即指数方程式Nx=N0e-bx用以描述DeeveyⅡ型存活曲线,幂函数式用以描述DeeveyⅢ型存活曲线。经建立其相应模型得到:Nx=2933.9e-0.417x(F=100.890;R2=0.9098);Nx=3384.7x-1.712(F=20.757;R2=0.6749)。通过对比F检验值和相关指数R2值可得,北京市紫椴种群存活曲线更趋于DeeveyⅡ型。该存活曲线形状趋于折线,存在明显的拐点,分别为5、7、8、9龄级处,表明紫椴个体在这几个龄级时存活力发生明显变化,这点不仅体现在存活曲线上,也体现在死亡率曲线和消失率曲线(图4)的变化中,可根据死亡率曲线和消失率曲线来分析各龄级紫椴个体死亡率的详细变化过程。
北京市紫椴种群的死亡率曲线和消失率曲线变化趋势一致,皆为波动的曲线。高峰值出现于第6龄级和第8龄级,表明这两个阶段种群中个体通过环境筛选的压力增大,种群损失较大,死亡率增大;而在第7和第9龄级分别存在低谷值,说明该种群中的紫椴个体在这几个阶段时对环境的适应性和耐受性都达到一个较高的水平,生存力增强,这也可能是由于在上一生长阶段强烈的环境压力的选择下存活下来的个体,其抗逆性和耐受性都得到了锻炼和加强,因此具有较高的生存力。除此之处,从图4中可以看出,9龄级以上的紫椴死亡率急剧上升至最高值且有继续增长的趋势,说明胸径大于27 cm的紫椴极容易死亡,这是由于老龄个体本身营养生长滞缓,繁殖能力消退,抗逆性减弱,加上环境筛选的压力,导致了高死亡率。因此,在对该紫椴种群进行管理时,要特别关注这些老龄紫椴个体。
4.4 种群数量动态预测
以北京市紫椴种群各龄级株数为原始数据,将各径级数据换算成株/hm2,按照一次平均移动法计算出各龄级在未来2个龄级、4个龄级、6个龄级及8个龄级后的株数,将结果绘成年龄与株数关系图(图5)。从同一时间上看,除M0和M2曲线在较小的径级有波动外,各曲线显示的紫椴株数随径级增加而整体表现为减少趋势。而随着时间的推移(从M0到M8)可以发现,除第2龄级和第4龄级外,其余各龄级的紫椴个体数量都随时间的延续而增加的。第2龄级的紫椴个体存活数在从M0到M2的时间内,有明显的减少;第4龄级的紫椴个体存活数在从M0到M4中的两个时间段内,都有一定数量的减少。从不同径级大小看,对大树(≥10径级)而言,紫椴个体存活数的增长率随着径级的增加明显减少,在M2、M4、M6曲线上均表现如此。
因此,时间序列预测结果表示北京市紫椴种群具有一定的增长和恢复能力,与种群动态量化结果一致。可以推断,如果能维持种群现有的环境状况,没有外来干扰,北京市紫椴种群将在一定时期内保持相对稳定的种群数量。而要使北京市的紫椴种群尽快恢复状大,则需加强对种群的管理,特别在幼苗期和老龄期加强抚育管理,对紫椴幼苗、胸径9~12 cm(4径级)、胸径27 cm(10径级)以上的紫椴个体予以特殊关注,弄清这些个体存活率降低的原因并采取相应的措施提高其存活率;同时,在本研究野外调查中发现,紫椴老龄个体容易感染病虫害,因此还要对胸径27 cm以上的老龄紫椴个体实施特殊的保护措施。
5 结论
北京市地区紫椴种群林下幼苗缺乏,种群天然更新较差。这可能是由于紫椴种子休眠的特性以及霉烂的影响,导致紫椴种子的自然萌发率低,更新幼苗少,且自然状态下见到的往往是以萌芽更新为主的紫椴林。本地区紫椴种群拥有较多的幼树和小树,種群年龄结构为增长型,但种群结构也并非理想状态,中树相对缺乏、大树数量明显不足,这也会影响种群的健康发展。该紫椴种群的存活曲线趋于Deevey Ⅱ型,个体在非成年时期具有较大的个体死亡量,而胸径为15~18 cm(6径级)和21~24 cm(8径级)的紫椴个体达到死亡率的两个峰值,这是由于紫椴自身生长规律的影响,使这两个生长阶段的紫椴个体抗逆性较低,通过环境筛选的压力增大,从而导致死亡率升高。再者,胸径大于27 cm(10径级)死亡率大大增加,这是由于紫椴老龄个体营养生长滞缓,抗逆性减弱,再加上这部分个体易感染病虫害所致。
种群数量特征是种群生物学特性与多种环境因素共同作用的结果[2],只要环境条件改善,目前的不利状态有可能改变。北京市紫椴种群具有一定的增长和恢复能力,种群在不考虑外部干扰时种群发展呈增长型;而在考虑外部干扰时仍有一定的增长性,但增长较为缓慢,这说明外部干扰对紫椴种群的发展影响较大。因此,如果能维持种群现有的环境状况,没有外来干扰,北京市紫椴种群将在一定时期内保持相对稳定的种群数量。
要使北京市的紫椴种群尽快恢复状大,需加强对种群的管理。要保证种群持续发展至少需要补充相应数量的幼苗,否则种群将停止增长甚至走向衰退,因此应采取措施来弥补由于种子休眠和霉烂而导致的紫椴幼苗库不足问题。对胸径为15~18 cm和21~24 cm的紫椴个体要予以特殊关注,加大对其生长生理等的科学研究,弄清这些个体存活率降低的原因,并采取相应的措施降低死亡率、提高生存力。此外,对胸径大于27 cm的紫椴老龄个体,需及时采取有效措施加大对病虫害的防治和治理,保护现有立木资源,平衡种群年龄结构,促进种群恢复和发展。
致谢:感谢导师张志翔教授对本次调查的指导,参与野外调查的北京林业大学自然保护区学院的蒲真、王潇、李俊同学,林学院许少聪、孟泽林同学,以及松山国家级自然保护区管理处和雾灵山自然保护区管理处对本次调查研究的大力支持!
参考文献:
[1]Crawley.M.J.Plant Ecology[M].London:Blackwell Scientific Publications,1986:97~185.
[2]Manuel,C.Molles,J.Ecology,Concept and Applications (2nd edn.)[M].New York:McGraw-Hill Companies,2002:186~254.
[3]国家林业局.国家重点保护野生植物名录(第一批)[Z].北京:农业部,1999.
[4]邱 知,金 川,于世勇.磨盘山次生林紫椴种群空间结构研究[J].林业资源管理,2012(4):62~65.
[5]邹学忠,阎忠林,刘清田,等.紫椴种子休眠原因及催芽处理方法的研究[J].辽宁林业科技,1991(5):6~10.
[6]乔 静,廉守喜,李贵莲,等.紫椴扦插繁殖技术的研究[J].防护林科技,2004(4):16~17.
[7]马建路.水曲柳、胡桃楸、黄波罗和紫椴适生立地条件的对比研究[J].林业资源管理,1996(4):52~56.
[8]赵 刚,邹学忠,吴月亮,等.紫椴适生立地条件研究[J].辽宁林业科技,2005(1):7~9.
[9]王洪峰,穆立蔷,王洪成,等.紫椴在自然和城市环境中生长状况比较研究[J].西北林学院学报,2012,27(5):101~105.
[10]杜凤国,苏春华,李云凤,等.紫椴和糠椴种子解剖构造的研究[J].吉林林学院学报,1994,10(2):99~102.
[11]王海南.紫椴种子休眠机理及休眠解除技术的研究[D].哈尔滨:东北林业大学,2012.
[12]陈香茗,赵秀海,夏富才,等.长白山紫椴种子雨的时空分布格局[J].东北林业大学学报,2011,39(1):7~10.
[13]钟德胜,马志伟,孙元发.哈尔滨地区紫椴的种子雨和种子库的观察[J].黑龙江生态职业学院学报,2008,21(5):44~45.
[14]刘赢男.紫椴天然种群遗传多样性和遗传分化的研究[D].哈尔滨:东北林业大学,2006.
[15]穆立蔷.紫椴种群地理变异与环境相关性研究[D].哈尔滨:东北林业大学,2006.
[16]马 瑶.不同地理分布区紫椴形态解剖结构研究[D].哈尔滨:东北林业大学,2007.
[17]周 浩.不同地理条件下紫椴亚显微结构比较研究[D].哈尔滨:东北林业大学,2008.
[18]刘立平.紫椴地上器官化学成分的初步研究[D].哈尔滨:东北林业大学,2009.
[19]张 健,郝占庆,宋 波,等.长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性[J].应用生态学报,2007,18(8):1681~1687.
[20]聂绍荃.紫椴种群生态学研究[M].哈尔滨:东北林业大学出版社.1992.
[21]刘贵峰,丁 易,臧润国,等.天目山云杉种群分布格局[J].应用生态学报,2011,22(1):9~13.
[22]董 点,林天喜,唐景毅,等.紫椴生物量分配格局及异速生长方程[J].北京林业大学学报,2014,36(4):54~63.
[23]陈晓德.植物种群与群落结构动态量化分析方法研究[J].生态学报,1998,18(2):214~217.
[24]丁国民,刘兴明,倪自银,等.甘肃祁连山青海云杉种群数量动态的初步研究[J].植物资源与环境学报,2005,14(4):36~41.
[25]江 洪.云杉种群生态学[M].北京:中国林业出版社,1992.
[26]李晓笑,王清春,崔国发,等.濒危植物梵净山冷杉野生种群结构及动态特征[J].西北植物学报,2011,31(7):1479~1486.
[27]闫桂琴,赵桂仿,胡正海,等.秦岭太白红杉种群结构与动态的研究[J].应用生态学报,2001,12(6):824~828.
[28]吴承祯,洪 伟,谢金寿,等.珍稀濒危植物长苞铁杉种群生命表分析[J].应用生态学报,2000,11(3):333~336.
猜你喜欢
北京教育·普教版(2014年9期)2014-10-08
北京教育·普教版(2009年9期)2009-11-16
北京教育·普教版(2009年7期)2009-09-16
北京教育·普教版(2008年10期)2008-10-23
北京教育·普教版(2008年9期)2008-10-07
北京教育·普教版(2008年8期)2008-09-20
北京教育·普教版(2008年5期)2008-06-03
北京教育·普教版(2008年4期)2008-05-14
北京教育·普教版(2008年1期)2008-05-14
北京教育·普教版(2008年3期)2008-03-06
