
河南省纤毛鹅观草种质资源表型多样性分析及综合评价
2024-12-28 00:00:00张鹤山吴新江郭胜华田宏熊军波陆姣云刘洋
南方农业学报 2024年10期




摘要:【目的】分析纤毛鹅观草种质资源的表型多样性,筛选作为饲草利用的优良种质,为纤毛鹅观草种质挖掘和新品种培育提供优异资源和理论参考。【方法】以来自河南地区不同生态区域的15份野生纤毛鹅观草种质为材料,对其16个表型性状进行变异分析、相关分析、聚类分析和主成分分析,并以各主成分特征值贡献率为权重建立综合得分(D)评价数学模型,对纤毛鹅观草种质进行综合评价排序。【结果】16个表型性状在不同纤毛鹅观草种质间存在极显著差异(Plt;0.01,下同),旗叶宽度的变异系数最大(21.88%),其次为种子产量(17.88%)和旗叶长度(17.81%),种子宽度的变异最小,仅为5.67%;7个营养相关性状的平均变异系数为15.48%,其余9个繁殖相关性状的平均变异系数为10.82%。种子产量与小穗宽度、芒长度和千粒重显著相关(Plt;0.05,下同),而干草产量与旗叶宽度、叶鞘长度、花序长度、小穗数、小穗宽度和种子长度显著正相关,与小穗长度和芒长度极显著正相关。15份纤毛鹅观草种质资源在欧式距离10处可分为4个类群,第I类群包括5份种质,主要性状特征是植株较高,穗秆和花序较长,旗叶长宽中等;第II类群包括2份种质,主要特征是植株较矮,繁殖性状较为突出;第III类群包括3份种质,主要特征是旗叶较大,第一节间较长,干草产量较高,但其种子较小,种子产量低;第IV类群包括5份种质,主要特征是植株矮小。旗叶宽度、叶鞘长度、芒长度、小穗数和小穗长度是引起纤毛鹅观草不同居群表型性状差异的主要因子。【结论】河南南阳的4个种质资源(编号为RC13、RC10、RC15、RC12)综合性状表现较好,这些种质株高更高,旗叶更宽,饲草产量和种子产量相对更高,可用作纤毛鹅观草育种亲本材料或直接生产利用。
关键词:纤毛鹅观草;表型性状;变异;综合评价
中图分类号:S543.202.4文献标志码:A文章编号:2095-1191(2024)10-3003-10
Phenotypic diversity and comprehensive evaluation of Roegne⁃ria ciliaris(Trin.)Nevski germplasm resources in Henan
ZHANG He-shan1,WU Xin-jiang1,GUO Sheng-hua2,TIAN Hong1,XIONG Jun-bo1,LU Jiao-yun1,LIU Yang1*
(1Institute of Animal Husbandry and Veterinary Medicine,Hubei Academy of Agricultural Sciences/Key Laboratory ofAnimal Embryo Engineering and Molecular Breeding of Hubei,Wuhan,Hubei 430064,China;2Forest Seed andSeedling Management Station of Hubei Forestry Bureau,Wuhan,Hubei 430075,China)
Abstract:【Objective】To analyze the phenotypic diversity of Roegneria ciliaris(Trin.)Nevski germplasm resources,and to screen the excellent germplasms for forage utilization,which could provide excellent resources and theoretical reference for R.ciliaris germplasms exploration and new variety breeding.【Method】The 16 phenotypic traits of 15 wild R.ciliaris populations collected from different ecological regions in Henan were subjected to variation analysis,correla-tion analysis,cluster analysis and principal component analysis,and the comprehensive score(D)evaluation mathemati-cal model was established with the contribution rate of each principal component characteristic value as the weight to com-prehensively evaluate and rank the R.ciliaris germplasm resources.【Result】There were extremely significant differences in 16 phenotypic traits among different R.ciliaris germplasms(Plt;0.01,the same below).The variation coefficient of flag leaf width was the largest(21.88%),followed by seed yield(17.88%)and flag leaf length(17.81%).The variation of seed width was the smallest(only 5.67%).The average variation coefficients of 7 nutrition-related traits was 15.48%,and the average variation coefficient of the other 9 reproduction-related traits was 10.82%.Correlation analysis showed that seed yield was significantly affected by spikelet width,awn length and 1000-seed weight(Plt;0.05,the same below),while hay yield was significantly and positively correlated with flag leaf width,leafsheath length,inflorescence length,spikelet number,spikelet width and seed length,extremely significantly correlated with spikelet length and awn length.Through cluster analysis,the 15 R.ciliaris germplasm resources could be classified into 4 taxa at Euclidean distance 10.The I taxon included 5 germplasms.The main trait characteristics were taller plants,longer spike rod and inflorescence,and medium length and width of flag leaves.The II taxon included 2 germplasms,and the trait characteristics were more prominent reproduction-related traits and shorter plants.The III taxon included 3 germplasms,and the main trait characteristics were larger flag leaves,longer internode,higher hay yield and lower seed yield.The IV taxon included 5 germplasms,and the main characteristics were short plants.Principal component analysis showed that flag leaf width,leafsheath length,awn length,spikelet number and spikelet length were the main factors causing differences in phenotypic traits among different populations of R.ciliaris.【Conclusion】The 4 germplasm resources originated from Nanyang,Henan(numbers as RC13,RC10,RC15,RC12)perform better in terms of overall traits,with higher plant,wider flag leaf,and higher forage yield and seed yield,which are expected to be used as parental materials for R.ciliaris breeding or be applied directly in produc-tion.
Key words:Roegneria ciliaris(Trin.)Nevski;phenotypic traits;variation;comprehensive evaluation
Foundation items:China Agriculture Research System(CARS-34-48);Hubei Agricultural Science and Technology Innovation Project(2021-620-000-001-21)
0引言
【研究意义】纤毛鹅观草[Roegneria ciliaris(Trin.)Nevski]是禾本科小麦族鹅观草属的多年生草本植物,适应性强,对温度和干旱耐受性极强,其营养体生长茂盛,再生能力较强,粗蛋白含量高,马、牛、羊、兔、鹅等喜食,是各类家畜夏秋放牧、冬春补饲的优良牧草,广泛分布于我国东北、华北、华东、华中及西北等区域(陈默君和贾慎修,2000;孔令娜等,2009)。纤毛鹅观草在我国利用较早,1990年我国审定登记了第一个饲用型纤毛鹅观草新品种赣饲1号(王加亭和侯湃,2022),每公顷干草产量16.7 t(催忠刚等,2014),为当地草食畜牧业生产和草地生态建设发挥了重要作用。此外,纤毛鹅观草亦具有麦类作物抗病和抗逆的优良基因,常被用作麦类作物育种改良的材料(万永芳等,1997;杨欣明等1999)。因此,开展纤毛鹅观草种质评价与挖掘研究,对筛选优异纤毛鹅观草优异种质资源和新品种培育具有重要意义。【前人研究进展】表型性状是植物为适应不同时空环境而形成的特异性表现(Grasseinetal.,2010;Lajoie and Vellend,2018),是植物内在组织生长分化的外在表现,在一定程度上能反映出物种基因的差异,体现物种的多样性(宫珂等,2019)。通过植物表型性状研究其野生居群的形态多样性是可行的(徐炳声,1998),已在水稻(Oryza sativa)(陈越等,2019)、燕麦(Avenanuda)(史京京等,2019)及鹰嘴豆(Cicer arietinum)(于海天等,2021)等物种内得到应用。鹅观草属植物因其较强的抗逆性在世界各地有广泛分布,在不同生态区域形成了显著差异的表型特征。研究发现,鹅观草(R.kamoji)不同居群间具有显著的性状分化,表现出丰富的性状多样性(孙宗华等,2017),并且这些性状分化与生长环境显著相关(肖海峻等,2007b)。肖海峻等(2007a)对来自四川、云南、新疆、甘肃、内蒙古、宁夏及山西等地的鹅观草种质15个表型性状进行分析,结果发现鹅观草株高、穗长、旗叶长、旗叶宽、小穗数、小花数和穗下第一节间长等表型性状具有丰富的多样性,且采集于西南地区(四川、云南等)的鹅观草表型多样性比西北地区(新疆、甘肃、内蒙古等)更丰富。高飞等(2006)研究也发现,表型性状在不同中华鹅观草(R.sinica)居群间或居群内具有差异,表明鹅观草属植物在长期适应自然环境的过程中形成了多样的表型性状,提高了物种适应环境的能力,为优异种质的筛选打下基础。根据表型性状的差异评价筛选优良种质资源具有可行性(李景欣,2005)。郜战宁等(2023)通过7个表型性状评价了143份大麦种质资源并筛选出多个优异种质资源。尽管纤毛鹅观草研究开展较早,但多集中在染色体(卢宝荣等,1988;蔡联柄和冯海生,1998)、抗病性(翁益群和刘大钧,1989)和抗除草剂(吕爱枝等,2006)等方面,鲜见有关纤毛鹅观草表型性状多样性评价及种质筛选方面的研究报道。自20世纪90年代赣饲1号纤毛鹅观草登记以来,未有纤毛鹅观草新品种审定登记。【本研究切入点】长江流域及其以北地区属于北亚热带过渡带,夏季炎热,冬季冷凉,大多数暖季型牧草冬季枯黄,甚至不能越冬,而冷季型牧草在夏季发生“夏枯”现象,即夏季生长受到抑制甚至死亡,而纤毛鹅观草具有很强的耐热和耐寒性,是一种优良的牧饲兼用型草种。当前,鹅观草属植物的研究多集中于鹅观草物种,且种质资源多采集于我国西南、西北及北方地区,华中地区的鹅观草属植物相关研究鲜见报道。【拟解决的关键问题】对采集于河南地区的纤毛鹅观草种质资源开展表型性状多样性研究,并进行主成分分析及综合评价,以期筛选出具有潜在饲用价值的优良种质材料,为纤毛鹅观草种质挖掘和新品种培育提供优异资源和数据支撑。
1材料与方法
1.1试验材料
试验材料为采集于河南地区的野生纤毛鹅观草种质资源,具体信息见表1。
1.2试验地概况
试验地位于湖北省武汉市江夏区国家牧草种质资源圃。地理坐标30°16′05″N,114°08′48″E;海拔23 m。土壤为丘陵黄土,pH 6.8,肥力中等。年均降水量1200 mm,年均气温16.7℃,无霜期260~270 d。
1.3试验方法
1.3.1试验设计设2个试验区块,一个试验区块用于开花期前相关性状及草产量测定,另一试验区块用于开花期后相关性状及种子产量测定。每个区块内试验小区采用随机区组设计,小区面积15m2(3 m×5 m),3次重复,共15份材料,45个小区。试验材料于9月播种,条播,行距30 cm,播种深度2~3 cm,每个材料按种子用价统一播种量,每个小区种植10行;试验小区以复合肥(N∶P2O5∶K2O=15∶15∶15)和过磷酸钙作底肥,用量分别为150和75kg/ha。试验期间不再施肥和灌溉。
1.3.2指标测定及方法开花期测定株高(植株从地面至最高点的绝对高度)、旗叶长度(植株旗叶从基部至叶尖的长度)、旗叶宽度(旗叶中部最宽处的长度)、叶鞘长度(植株旗叶叶鞘的长度)、第一节间长(植株自上往下第一个节间的长度)、小穗数(每个花序上的小穗数量)和干草产量(参照《草品种区域试验技术规程禾本科牧草》标准执行)。蜡熟期测定花序长度(整个花序的绝对长度)、小穗长度(花序中部小穗不包括芒长度的小穗绝对长度)、小穗宽度(花序中部小穗最宽处的长度);成熟期测定穗秆长度(花序基部至第一个茎节的长度)、芒长度(小穗中部第一小花外稃芒的绝对长度)、种子长度(不含芒长度的成熟种子绝对长度)、种子宽度(成熟种子背腹面最宽处长度)、千粒重(1000粒正常成熟种子的重量)和种子产量(成熟后收获每个小区种子,自然晒干,称重)。除干草产量、千粒重和种子产量外,其余性状每重复测定10株,取平均值。
1.4统计分析
利用Excel 2019和SPSS 21.0进行变异分析和F测验,使用Pearson相关系数进行性状间相关分析;利用SPSS 21.0对表型性状进行聚类分析、主成分分析和成分得分系数计算。按以下公式对原始数据进行标准化处理:
Dij=(Xij-X)/δ
式中,Dij为第i性状第j材料的标准值,X为第i性状的均值,δ为第i性状的标准差。然后以各性状成分得分系数为权重计算各材料每个主成分得分值。
2结果与分析
2.1纤毛鹅观草表型性状变异分析结果
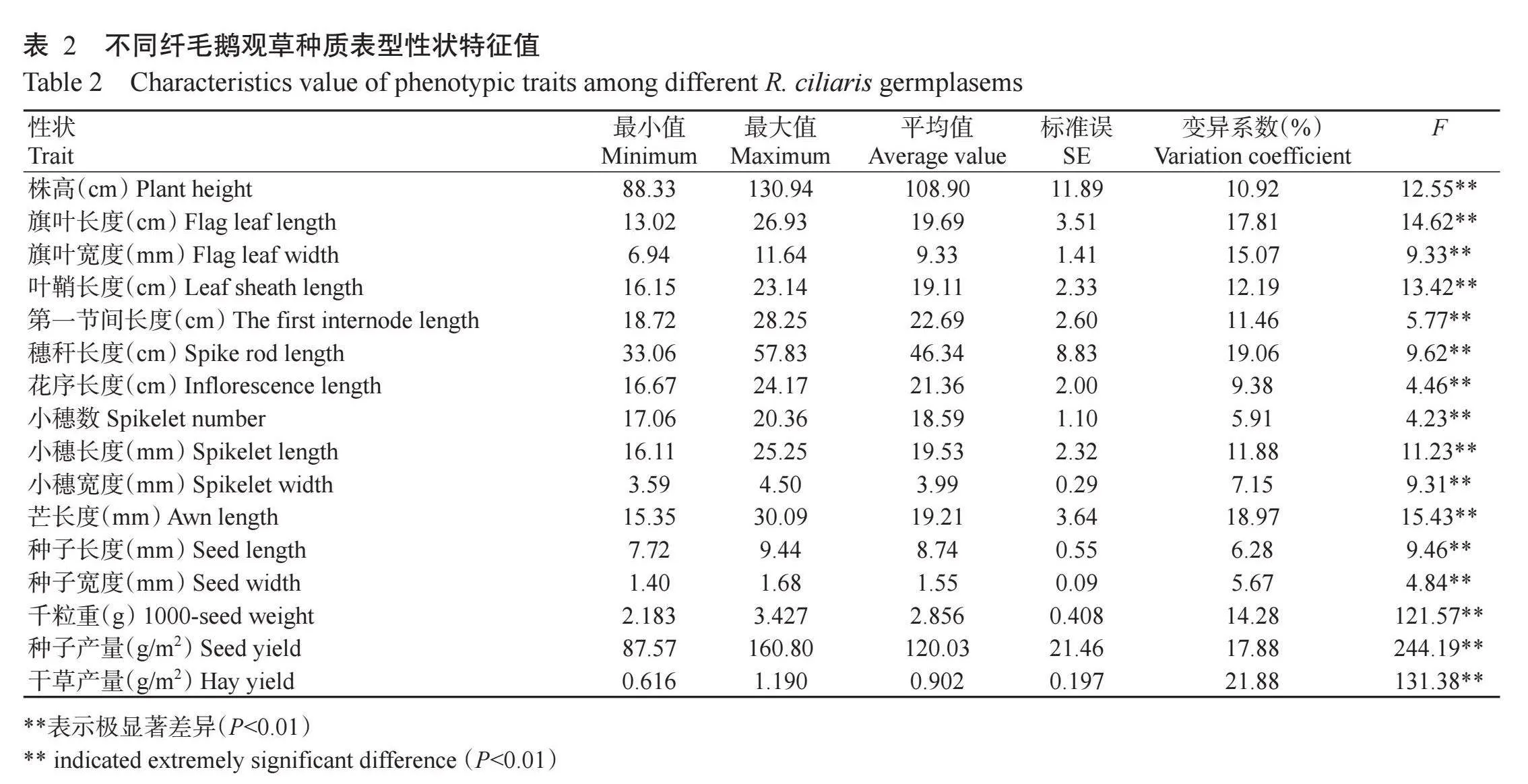
15份纤毛鹅观草种质资源表型性状特征值及变异情况如表2所示。由不同材料间F测验值可知,16个表型性状在不同纤毛鹅观草间存在极显著差异(Plt;0.01,下同),变化幅度最大的性状是旗叶长度,最大值为最小值的2.07倍,其次为芒长度和干草产量,最大值分别为最小值的1.96倍和1.93倍;变化幅度最小的性状是小穗数,最大值仅为最小值的1.19倍。干草产量的变异系数最大,为21.88%,种子宽度的变异最小,仅为5.67%。此外,7个营养相关性状(株高、旗叶长度、旗叶宽度、叶鞘长度、第一节间长度、穗秆长度和干草产量)的变异系数均大于10.00%,平均值为15.48%,其余9个繁殖相关性状的平均变异系数为10.82%,其中花序长度、小穗数、小穗宽度、种子长度和种子宽度5个性状的变异系数低于10.00%,说明表型性状变异分化在不同纤毛鹅观草间有差异,营养相关性状比繁殖相关性状变异程度更大。
2.2纤毛鹅观草表型性状相关分析结果
纤毛鹅观草表型性状间相关系数如表3所示。各性状间具有不同程度的正向或负向相关性。株高与旗叶宽度、叶鞘长度、穗秆长度、花序长度、种子长度和千粒重极显著正相关,与第一节间长度和小穗宽度显著正相关(Plt;0.05,下同),说明植株越高,营养相关性状指标越大,花序越长,种子粒也越大;旗叶长度与叶鞘长度和第一节间长度极显著正相关,与穗秆长度、花序长度、种子长度和种子宽度显著正相关。旗叶宽度与叶鞘长度、第一节间长度、花序长度、小穗宽度、种子长度及种子千粒重极显著正相关,与穗秆长度、小穗长度、种子宽度和干草产量显著正相关。叶鞘长度与第一节间长度、花序长度、种子长度、种子宽度极显著正相关,与穗秆长度、种子千粒重和干草产量显著正相关。第一节间长度与花序长度、种子长度和种子宽度极显著正相关,与穗秆长度和种子千粒重显著正相关。穗秆长度与花序长度和千粒重显著正相关,而与小穗数显著负相关,说明穗秆越长,花序越长,种子越大,但小穗数较少。花序长度不仅受营养性状如株高、旗叶长度、旗叶宽度、叶鞘长度、第一节间长度的影响,并且也与穗秆长度、种子长度、种子千粒重和干草产量显著正相关。小穗数与穗秆长度显著负相关,与芒长度和干草产量显著正相关。小穗长度与小穗宽度、芒长度、干草产量极显著正相关,与种子长度和种子千粒重显著正相关。种子产量与小穗宽度、芒长度和千粒重显著正相关,而干草产量与旗叶宽度、叶鞘长度、花序长度、小穗数、小穗宽度及种子长度显著正相关,与小穗长度和芒长度极显著正相关。
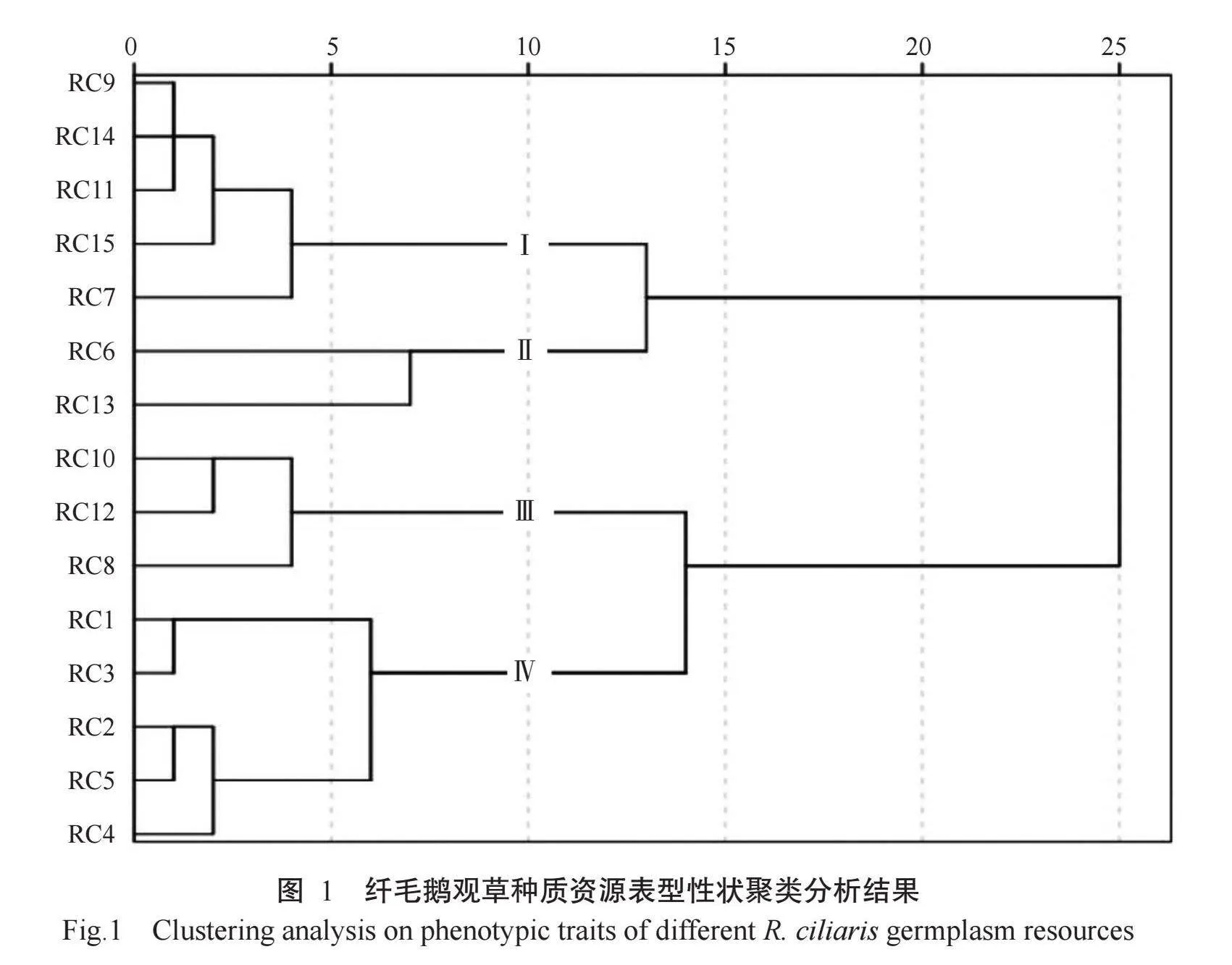
2.3纤毛鹅观草种质资源聚类分析结果
基于16个表型性状的纤毛鹅观草聚类分析结果如图1所示。在欧式距离10处将15份纤毛鹅观草种质聚类为4个类群,第Ⅰ类群包括5份种质,其主要性状特征是植株较高,平均株高120.46 cm,穗秆和花序较长,平均值分别为55.54和22.30 cm,旗叶长宽中等;该类群平均种子产量较高(131.60 g/m2),但干草产量偏低,平均产量仅为0.828 g/m2,小穗较少,芒短;第Ⅱ类群包括2份种质(RC6和RC13),该类群植株较矮(平均株高为109.22 cm),但繁殖相关性状较为突出,具有较多的小穗数,较长的小穗长度、小穗宽度和芒长度,种子较大,其千粒重较重(平均值为3.33 g),种子产量较大(平均值为153 g/m2),干草产量也较高,比所有材料平均干草产量增产27.6%,属于种子产量和饲草产量较优良的类群;第Ⅲ类群包括3份种质,主要特征是旗叶较大,平均旗叶长度和旗叶宽度分别为21.36和10.27 mm,叶鞘长(平均值为21.09 cm),第一节间较长,干草产量较高,属于营养相关性状较突出的群体,但其种子较小,千粒重较轻,种子产量低(平均值为90.63 g/m2);第Ⅳ类群包括5份种质,该类群植株矮小,平均株高仅95.84 cm,比所有材料平均株高低11.2%;旗叶长宽、第一节间长度、花序长度、小穗长度、小穗宽度、千粒重、干草产量等性状均为4个类群中最低,但种子产量较高,平均为112.72 g/m2,属于潜在种子高产型。
2.4纤毛鹅观草表型性状主成分分析结果
纤毛鹅观草16个表型性状的主成分分析结果如表4所示。纤毛鹅观草前4个特征值的累计贡献率为85.22%,表明前4个主成分能表达所有表型性状的大部分生物学信息。第一主成分(PC1)的特征值为6.990,贡献率为43.69%,占累计贡献率的51.3%,其中载荷较高的性状有旗叶宽度(0.893)、叶鞘长度(0.887)、千粒重(0.845)、种子长度(0.837)和株高(0.813),表明PC1反映旗叶和种子千粒重特征。第二主成分(PC2)的特征值为3.506,贡献率为21.91%,载荷较高的性状有芒长度(0.899)和小穗长度(0.762),表明PC2反映纤毛鹅观草的小穗特征。第三主成分(PC3)的特征值为1.865,贡献率为11.66%,主要反映小穗数(0.591)和种子产量(-0.643)特征。第四主成分(PC4)的特征值为1.274,贡献率为7.96%,载荷较大的性状有种子宽度(0.274)和种子产量(0.442),说明PC4反映种子的产量因子。公因子方差分析结果显示,所有性状的公因子方差均大于0.800,说明前4个主成分对所有性状有很好的解释能力,能反映所有性状特征。
2.5纤毛鹅观草种质资源综合评价
为进一步对纤毛鹅观草种质资源进行综合评价,根据主成分分析结果,以前4个主成分得分为依据,以各主成分特征值贡献率为权重,建立综合得分(D)评价模型:D=(PC1×43.69+PC2×21.91+PC3×11.66+PC4×7.96)/85.22,结果如表5所示。综合性状最好的材料为RC13,D为1.07,其次为材料RC10和RC15,D分别为0.80和0.60;材料RC8和RC5的综合性状最差,D为-0.89和-0.70。15份纤毛鹅观草种质的D排序为RC13gt;RC10gt;RC15gt;RC12gt;RC6gt;RC7gt;RC4gt;RC14gt;RC11gt;RC9gt;RC3gt;RC1gt;RC2gt;RC5gt;RC8。
3讨论
植物性状变异是在进化过程中为适应自然环境而引起的基因和表型性状差异化,是造成植物特异性的重要因素之一,也是培育新品种的基础(王晓鸣等,2022)。本研究结果显示,不同纤毛鹅观草种质在株高、旗叶长宽、第一节间长度、小穗长度等性状上分化明显,变异系数达10.00%以上,说明来自不同生态区域的15个纤毛鹅观草种质间表型性状存在明显差异。前人研究表明,采集于河南郑州区域的纤毛鹅观草种质株高、顶节长、旗叶长、穗长和小穗数等指标均值分别为122.3 cm、46.80 cm、25.63 cm、28.49 cm和23.4个(卢宝荣等,1988),明显高于采集于陕西省武功地区的纤毛鹅观草株高、旗叶长度、穗长和小穗数(魏秀华等,2009)。这种表型性状的差异代表了物种对自然环境的适应性(王娅丽和李毅,2008;宫珂等,2019),也为纤毛鹅观草特异性种质资源的筛选提供物质基础。肖海峻等(2007a)研究也发现,长期生长在不同生态环境的鹅观草会分化出适应自然环境的表型特征,且这种特征在不同地区的居群间具有显著的差异性。此外,本研究中纤毛鹅观草营养相关表型性状(如株高、旗叶长度、旗叶宽度、第一节间长度等)分化程度较繁殖相关性状(如花序长度、小穗长度、小穗宽度、种子长度和种子宽度)更大,在海南粗榧(Cephalotaxusmannii)(向志强等,1999)和鹅观草(肖海峻等,2007b)的研究中也得到类似结论,说明营养器官更易受到环境的影响,有助于纤毛鹅观草优良饲用价值种质材料筛选及新品种选育。
纤毛鹅观草的饲用价值主要体现在饲草产量和种子生产两个方面,通过表型性状可间接评价种质资源的饲用价值。本研究发现,干草产量和种子产量与其他表型性状具有不同程度的相关性,旗叶宽度、叶鞘长度、花序长度、小穗数、小穗长度和小穗宽度、芒长度和种子长度是反映饲草产量的主要指标,而种子产量与芒长度和千粒重显著相关,可根据育种目标进行相应选择。本研聚类分析结果也显示,旗叶较大、叶鞘较长、干草产量较高的纤毛鹅观草种质划分为同一个类群(类群Ⅲ),为优良育种种质资源筛选提供参考依据。
利用主成分分析对种质资源进行综合评价是筛选优异种质的有效方法,已在花生(殷冬梅等,2011)、樱桃番茄(张静等,2014)、小豆(周桂梅等,2023)、蚕豆(王琳琳等,2023)等作物中得到应用。本研究结果显示,旗叶宽度、叶鞘长度、芒长度、小穗数和小穗长度是引起纤毛鹅观草不同居群表型性状差异的主要因子。肖海峻等(2007b)研究也发现,旗叶宽度和小穗数是引起鹅观草居群变异的主要因子。肖海峻等(2007a)研究发现,株高和穗长度对鹅观草的变异影响更大,而本研究中旗叶宽度和叶鞘长度是引起纤毛鹅观草变异的主要性状,这可能是由于不同物种或不同表型性状指标引起的。本研究主成分综合评价结果显示,采自河南省南阳市的种质资源(编号为RC13、RC10、RC15和RC12)综合性状表现较好,这些种质株高较高,旗叶较宽,第一节间更长,饲草产量和种子产量相对更高,有望作为纤毛鹅观草育种的亲本材料或优良资源加以利用。
鹅观草属是小麦族最大的一个属,包括130个种(Baum etal.,1991),种间形态变异复杂,在一些种的鉴定上存在分歧(蔡联炳,1997),因此研究者常结合表型性状和核型分析进行种水平的鉴定。卢宝荣等(1988)对鹅观草、纤毛鹅观草和竖立鹅观草(R.japonensis)3个物种的形态和核型进行研究,结果发现纤毛鹅观草和竖立鹅观草在形态上虽有差异,但染色体核型基本一致,因此认为二者应为同一种。在纤毛鹅观草种内,不同生态区域的纤毛鹅观草资源在染色体核型上也有差异。魏秀华等(2005)发现不同来源的纤毛鹅观草染色体数目均为28条,但核型公式存在变异,中部和近中部着丝点染色体数目在不同材料间存在差异。孙义凯和董玉琛(1992)研究也得出东北地区的纤毛鹅观草染色体核型为2n=4x=28=22 m(2SAT)+6sm(2SAT),而蔡联炳等(1998)研究得出核型2n=4x=28=20m+8sm(4SAT),因此不同来源的纤毛鹅观草核型有差异,使得准确鉴定纤毛鹅观草种内不同来源种质的差异性成为可能。此外,董晗(2012)通过分子标记技术研究了黄淮海地区36个纤毛鹅观草居群的遗传多样性,并划分了不同的生态类群。本研究根据直观的表型性状对不同来源的纤毛鹅观草进行多样性分析,并发现干草产量和种子产量与部分表型性状显著相关,为优异纤毛鹅观草种质资源的筛选提供参考。然而,为更准确地了解不同纤毛鹅观草间的内在差异,将表型性状、核型分析和分子标记综合应用于纤毛鹅观草种内材料的鉴定评价,是一种有效的方法。
4结论
纤毛鹅观草表型性状在不同居群间存在显著变异分化,旗叶宽度、叶鞘长度、芒长度、小穗数和小穗长度是引起不同居群间性状差异的主要因子。干草产量性状与旗叶特征、小穗特征相关,种子产量受小穗长度的影响,可据此表型性状筛选具有潜在饲用价值的纤毛鹅观草资源。在15个纤毛鹅观草种质中,来自河南省南阳市的4个种质资源具有更好的综合性状表现,可作为纤毛鹅观草育种亲本材料。
参考文献(References):
蔡联炳,冯海生.1998.鹅观草属五个类群的核型与进化[J].广西植物,18(1):35-40.[Cai L B,Feng H S.1998.Karyotypes and evolution of five taxa of RoegneriaC.koch[J].Guihaia,18(1):35-40.]doi:http://ir.nwipb.ac.cn/handle/363003/1066.
蔡联炳.1997.中国鹅观草属的分类研究[J].植物分类学报,35(2):148-177.[Cai L B.1997.A taxonomical study on the genus RoegneriaC.Koch from China[J].Acta Phyto-taxonomica Sinica,35(2):148-177.]doi:10.1007/BF02 009745.
陈默君,贾慎修.2000.中国饲用植物[M].北京:中国农业出版社.[Chen M J,Jia S X.2000.Forage plants in China[M].Beijing:China Agriculture Press.]
陈越,丁明亮,张敦宇,付坚,钟巧芳,肖素勤,柯学,程在全.2019.云南水稻种质资源农艺性状表型多样性分析及综合评价[J].南方农业学报,50(9):1922-1930.[Chen Y,Ding M L,Zhang D Y,Fu J,Zhong Q F,Xiao S Q,Ke X,Cheng Z Q.2019.Phenotypic diversity analysis and com-prehensive evaluation of agronomic characters of rice germplasm resources in Yunnan Province[J].Journal of Southern Agriculture,50(9):1922-1930.]doi:10.3969/j.issn.2095-1191.2019.09.05.
催忠刚,张海琴,黎琦,杨财容,黄娟,罗小梅,周永红.2014.3种鹅观草在成都平原的生产性能[J].草业科学,31(4):732-736.[Cui Z G,Zhang H Q,Li Q,Yang C R,Huang J,Luo X M,Zhou Y H.2014.Grass production of three Roegneriakamoji varieties in Chengdu plain[J].Pratacul-tural Science,31(4):732-736.doi:10.11829\j\issn.1001-0629.2013-0341.]
董晗.2012.不同居群纤毛鹅观草的遗传多样性研究[D].济南:山东师范大学.[Dong H.2012.The genetic diversity analysis of different Roegneria ciliaris populations[D].Jinan:Shangdong Normal University.]
高飞,柴守诚,高爱农.2006.基于形态学的中华鹅观草遗传多样性分析[J].麦类作物学报,26(3):12-17.[Gao F,Chai S C,Gao A N.2006.Genetic diversity of Roegneria sinica populations based on morphological analysis[J].Journal of Triticeae Crops,26(3):12-17.]doi:10.3969/j.issn.1009-1041.2006.03.003.
郜战宁,杨永乾,王树杰,冯辉,薛正刚.2023.143份大麦种质资源的综合评价[J].作物杂志,(5):59-65.[Gao Z N,Yang Y Q,Wang S J,Feng H,Xue Z G.2023.Comprehen-sive evaluation of 143 barley germplasm resources[J].Crops,(5):59-65.]Doi:10.16035/j.issn.1001-7283.2023.05.009.
宫珂,靳瑰丽,李陈建,岳永寰,王慧宁,韩万强.2019.天山北坡野生无芒雀麦的表型性状[J].生态学杂志,38(9):2615-2621.[Gong K,Jin G L,Li C J,Yue Y H,Wang H N,Han W Q.2019.Phenotypic traits of Bromus inermis on the northern slope of Tianshan Mountains[J].Chinese Jour-nal of Ecology,38(9):2615-2621.]doi:10.13292/j.1000-4890.201909.039.
孔令娜,陈卫平,冯金侠.2009.纤毛鹅观草的研究与利用[J].生物学通报,44(11):4-6.[Kong L N,Chen W P,Feng J X.2009.Study and utilization of Roegneria ciliaris[J].Bulletin of Biology,44(11):4-6.]doi:10.3969/j.issn.0006-3193.2009.11.002.
李景欣.2005.内蒙古冰草种质资源遗传多样性研究[D].呼和浩特:内蒙古农业大学.[Li J X.2005.Genetic diver-sity of germplasm resources of Agropyron cristatum in Inner Mongolia[D].Huhehot:Inner Mongolia Agricultural University.]
卢宝荣,颜济,杨俊良.1988.鹅观草属三个种的形态变异与核型的研究[J].云南植物研究,10(2):139-146.[Lu B R,Yan J,Yang J L.1988.The studies of morphological variations and karyotype analysis on the three Roegneria Species[J].Acta Botanica Yunnanica,10(2):139-146.]
吕爱枝,赵和,王天宇,王海波.2006.小麦近缘野生种抗除草剂Dalapon变异体的筛选[J].麦类作物学报,26(1):121-125.[LüA Z,Zhao H,Wang TY,Wang H B.2006.Selec-tion of herbicide resistant mutants in wild relatives of wheat[J].Journal of Triticeae Crops,26(1):121-125.]doi:10.3969/j.issn.1009-1041.2006.01.027.
史京京,薛盈文,郭伟,于崧,陆旺,于立河,胡慧影,郭灿,侯楠.2019.黑龙江西部地区引进的饲用燕麦种质资源遗传多样性分析及综合评价[J].南方农业学报,50(3):515-523.[Shi J J,Xue Y W,Guo W,Yu S,Lu W,Yu L H,Hu H Y,Guo C,Hou N.2019.Genetic diversity analysis and comprehensive evaluation of introduced Avena sativa L.germplasm resources in the western area of Heilongjiang[J].Journal of Southern Agriculture,50(3):515-523.]doi:10.3969/j.issn.2095-1191.2019.03.11.
孙义凯,董玉琛.1992.东北地区小麦族11种植物的核型报道[J].植物分类学报,30(4):342-345.[Sun Y K,Dong Y C.1992.Karyotypes of eleven species of triticeae in northeastern China[J].Acta Phytotaxonomica Sinica,30(4):342-345.]
孙宗华,张玥,邓梦秋,姜子小,焦振飞,周永红,张海琴.2017.7个鹅观草居群的形态学和细胞遗传学研究[J].植物遗传资源学报,18(6):1032-1038.[Sun Z H,Zhang Y,Deng M Q,Jiang Z X,Jiao Z F,Zhou Y H,Zhang H Q.2017.Cytogenetic and morphological studies on seven populations of RoegneriakamojiOhwi(Triticeae:Poaceae)[J].Jounal of Plant Genetic Resources,18(6):1032-1038.]doi:10.13430/j.cnki.jpgr.2017.06.005.
万永芳,颜济,杨俊良,刘法圈.1997.小麦近缘野生植物的赤霉病抗性研究[J].植物病理学报,27(2):107-111.[Wan Y F,Yan J,Yang J L,Liu F Q.1997.Study on wild rela-tivesof wheat for resistance to head scab[J].Acta Phyto-pathologica Sinica,27(2):107-111.]doi:10.13926/j.cnki.apps.1997.02.003.
王加亭,侯湃.2022.中国审定登记草品种集[M].北京:中国农业出版社.[Wang J T,Hou P.2022.China approved and registered grass varieties collection[M].Beijing:China Agriculture Press.]
王琳琳,钟洋敏,缪叶旻子,马瑞芳,刘庭付.2023.基于主成分和聚类分析的鲜食蚕豆农艺与品质性状综合评价[J].江苏农业学报,39(3):788-797.[Wang L L,Zhong Y M,Miao Y M Z,Ma R F,Liu T F.2023.Comprehensive evaluation of vegetable fababean in agronomic and qualitytraits by principal component analysis and cluster analysis[J].Jiangsu Journal of Agricultural Sciences,39(3):788-797.]doi:10.3969/j.issn.1000-4440.2023.03.020.
王晓鸣,邱丽娟,景蕊莲,任贵兴,李英慧,李春辉,秦培友,谷勇哲,李龙.2022.作物种质资源表型性状鉴定评价:现状与趋势[J].植物遗传资源学报,23(1):12-20.[Wang X O,Qiu L J,Jing R L,Ren G X,Li Y H,Li C H,Qin PY,Gu Y Z,Li L.2022.Evaluation on phenotypic traits of crop germplasm:Status and development[J].Journal of Plant Genetic Resources,23(1):12-20.]doi:10.13430/j.cnki.jpgr.20210802001.
王娅丽,李毅.2008.祁连山青海云杉天然群体的种实性状表型多样性[J].植物生态学报,32(2):355-362.[Wang Y L,Li Y.2008.Study on phenotypic diversity of cone andseed in natural populations of Picea crassifolia in Qilian Mountain,China[J].Chinese Journal of Plant Ecology,32(2):355-362.]doi:10.3773/j.issn.1005-264x.2008.02.012.
魏秀华,李付军,邱玉宾,李保华,李霞.2009.小麦族鹅观草属三个物种的关系研究[J].广西植物,29(1):28-31.[Wei X H,Li F J,Qiu Y B,Li B H,Li X.2009.Relationstudies on Roegneria amurensis,R.ciliaris and R.japonen-sis(Poaceae:Triticeae)[J].Guihaia,29(1):28-31.]doi:10.3969/j.issn.1000-3142.2009.01.005.
魏秀华,周永红,杨瑞武,丁春邦,张利,张海琴.2005.鹅观草属三个物种及其居群间核型变异研究[J].草业学报,14(2):57-62.[Wei X H,Zhou Y H,Yang R W,Ding C B,Zhang L,Zhang H Q.2005.Study on karyotypic variations of three Roegneria species and their accessions[J].Acta Prataculturae Sinica,14(2):57-62.]doi:10.3321/j.issn:1004-5759.2005.02.011.
翁益群,刘大钧.1989.鹅观草(RoegneriaC.Koch)与普通小麦(Triticumaestivum L.)属间杂种F1的形态、赤霉病抗性和细胞遗传学研究[J].中国农业科学,22(5):1-7.[Weng Y Q,Liu D J.1989.Morphology,scab resistance and cytogenetics of intergeneric hybrid of Triticum aesti-vum L.withRoegneriaC.koch(Agropyron)species[J].Scientia Agricultura Sinica,22(5):1-7.]
向志强,付永川,刘玉成,杜道林.1999.不同种群中海南粗榧(Cephalotaxusmannii)形态变异研究[J].广西植物,19(2):131-135.[Xiang Z Q,Fu Y C,Liu Y L,Du D L.1999.Study on the morphological variations of Cephalo-taxus mannii in different populations[J].Guihaia,19(2):131-135.]doi:10.3969/j.issn.1000-3142.1999.02.006.
肖海峻,徐柱,翟利剑,李玉冰,谢继红,孟利前.2007a.鹅观草居群表型多样性研究[J].西北植物学报,27(11):2222-2227.[Xiao H J,Xu Z,Zhai L J,Li Y B,Xie J H,MengL Q.2007a.Phenotypic diversity of RoegneriakamojiOhwi population[J].Acta Botanica Boreali-Occidentalia Sinica,27(11):2222-2227.]doi:10.3321/j.issn:1000-4025.2007.11.013.
肖海峻,徐柱,李临杭,马玉宝.2007b.鹅观草表型性状变异与生境间的相关性[J].中国草地学报,29(5):22-30.[Xiao H J,Xu Z,Li L H,Ma Y B.2007b.Correlation analysis between the variation of phenotypic characters and the habitat of RoegneriakamojiOhwi[J].Chinese Journal of Grassland,29(5):22-30.]doi:10.3969/j.issn.1673-5021.2007.05.004.
徐炳声.1998.中国植物分类学中的物种问题[J].植物分类学报,36(5):470-480.[Xu B S.1998.The species prob-lem in plant taxonomy in China[J].Acta Phytotaxonomica Sinica,36(5):470-480.]
殷冬梅,张幸果,王允,崔党群.2011.花生主要品质性状的主成分分析与综合评价[J].植物遗传资源学报,12(4):507-512,518.[Yin D M,Zhang X G,Wang Y,Cui D Q.2011.Principal component analysis and comprehensive evaluation on quality traits of peanut parents[J].Journal of Plant Genetic Resources,12(4):507-512.]doi:10.13430/j.cnki.jpgr.2011.04.007.
杨欣明,李立会,李秀全,董玉琛.1999.向普通小麦导入纤毛鹅观草抗黄矮病基因的研究I.F1和BC1的产生及其细胞遗传学[J].遗传学报,26(4):370-376.[Yang X M,Li L H,Li X Q,Dong Y C.1999.Introduction of genes resistant barly yellow dwarf virus from Roegneria ciliaris to com-mon wheat I.Production and cytogenetics of F1 and BC1 progenies[J].Acta Genetica Sinica,26(4):370-376.]doi:10.1038/sj.cr.7290029.
于海天,杨峰,吕梅媛,胡朝芹,杨新,郑爱清,王玉宝,何玉华,王丽萍.2021.伊朗鹰嘴豆种质资源农艺性状遗传多样性分析及综合评价[J].南方农业学报,52(3):769-778.[Yu H T,Yang F,LüM Y,Hu C Q,Yang X,Zheng A Q,Wang Y B,He Y H,Wang L P.2021.Genetic diversity and comprehensive evaluation of agronomic traits of chick-pea resources from Iran[J].Journal of Southern Agricul-ture,52(3):769-778.]doi:10.3969/j.issn.2095-1191.2021.03.025.
张静,常培培,梁燕,梁静,赵建涛,邹志荣,黄薪历,胡晓辉.2014.樱桃番茄主要品质性状的主成分分析与综合评价[J].北方园艺,(21):1-7.[Zhang J,Chang P P,Liang Y,Liang J,Zhao J T,Zou Z R,Huang X L,Hu X H.2014.Principal component analysis and comprehensive evalua-tion on quality traits of cherry tomato[J].Northern Horti-culture,(21):1-7.]
周桂梅,陈琳,刘振兴,陈健,亚秀秀,刘志军.2023.小豆种质资源农艺性状鉴定与综合评价[J].江苏农业科学,51(21):44-49.[Zhou G M,Chen L,Liu Z X,Chen J,Ya X X,Liu Z J.2023.Identification and comprehensive evalua-tion of agronomic traits of adzuki bean germplasmresources[J].Jiangsu Agricultural Sciences,51(21):44-49.]doi:10.15889/j.issn.1002-1302.2023.21.007.
Baum B R,Yen C,Yang J L.1991.Roegneria:Its generic li-mits and justification for its recognition[J].Candadian Journal of Botany,69(2):282-294.doi:10.1139/b91-038.
Grassein F,Till-Bottraud I,Lavorel S.2010.Plant resource-use strategies:The importance of phenotypic plasticity in response to a productivity gradient for two subalpine spe-cies[J].Annals of Botany,106(4):637-645.doi:10.1093/aob/mcq 154.
Lajoie G,Vellend M.2018.Characterizing the contribution of plasticity and genetic differentiation to community-level trait responses to environmental change[J].Ecology and Evolution,8(8):3895-3907.doi:10.1002/ece3.3947.
(责任编辑陈燕)
猜你喜欢
趣味(数学)(2020年4期)2020-07-27 01:44:16
支部建设(2020年15期)2020-07-08 12:34:32
江苏农业科学(2016年12期)2017-04-05 15:20:57
江苏农业科学(2016年8期)2017-02-15 19:33:33
江苏农业科学(2016年8期)2017-02-15 19:32:36
科学与财富(2016年26期)2016-12-01 21:32:23
医学信息(2016年29期)2016-11-28 09:43:04
大学教育(2016年11期)2016-11-16 20:33:18
企业导报(2016年9期)2016-05-26 20:52:48
百科知识(2015年18期)2015-09-10 07:22:44
