大蒜素在体外脑出血细胞模型中的抗炎和抗氧化应激作用
2024-09-29 00:00:00桂如林陆聪张昊张醒醒佘捷吴卫疆
江苏大学学报(医学版) 2024年5期

[摘要]" 目的: 探讨体外脑出血模型中大蒜素的作用。方法: 选择新生SD大鼠提取原代星形胶质细胞(astrocyte,ASC);分别采用氧化血红素和炎症因子(IL-1α、TNF-α、C1q)处理原代ASC,构建对应的模拟脑出血后氧化应激损伤和炎症反应的细胞模型,并用不同浓度大蒜素进行干预。采用CCK-8法检测细胞活力,CFDA探针检测细胞内活性氧聚集水平,Tunel染色法观察凋亡程度,流式细胞仪检测细胞凋亡率,蛋白免疫印迹检测凋亡相关蛋白、ASC表型相关蛋白及血红素加氧酶-1(heme oxygenase-1,HO-1)、核因子E2相关因子2(nuclear factor erythroid 2-related factor 2,Nrf2)等表达变化。结果: 大蒜素处理可增强ASC对血红素毒性的抵抗作用,减少血红素引起的活性氧蓄积,减少细胞凋亡,降低促凋亡蛋白表达,并且可抑制A1表型蛋白C3表达,上调A2表型蛋白S100和转化生长因子表达。此外,大蒜素干预能够上调Nrf2和HO-1表达。结论: 大蒜素在体外脑出血模型中可减轻氧化应激损伤和炎症反应,发挥神经保护作用。
[关键词]" 脑出血;星形胶质细胞;大蒜素;炎症;凋亡;氧化应激
[中图分类号]" R743.34" [文献标志码]" A" [文章编号]" 1671-7783(2024)05-0414-09
DOI: 10.13312/j.issn.1671-7783.y230139
[引用格式]桂如林,陆聪,张昊,等. 大蒜素在体外脑出血细胞模型中的抗炎和抗氧化应激作用[J]. 江苏大学学报(医学版), 2024, 34(5): 414-422.
[基金项目]国家自然科学基金青年基金资助项目 (81901250)
[作者简介]桂如林(1997—),女,硕士研究生;吴卫疆(通讯作者),博士,讲师,E-mail: jiang7607@163.com
Anti-inflammatory and anti-oxidative stress effects of allicin in intracerebral hemorrhage cell model
GUI Rulin, LU Cong, ZHANG Hao, ZHANG Xingxing, SHE Jie, WU Weijiang
(School of Medicine, Jiangsu University, Zhenjiang Jiangsu 212013, China)
[Abstract]" Objective: To investigate the effect of allicin in vitro intracerebral hemorrhage model. Methods: Primary astrocyte (ASC) was extracted from neonatal SD rats. Primary ASC was treated with oxidized heme and inflammatory factors (IL-1α, TNF-α and C1q), respectively, and the corresponding cell model of oxidative stress injury and inflammatory response after intracerebral hemorrhage was constructed, and various concentrations of allicin were chosen for further experiments. The cell viability was evaluated by CCK-8, the intracellular reactive oxygen species accumulation level was measured by CFDA probe, the degree of apoptosis was observed by Tunel staining, the apoptosis rate was detected by flow cytometry, and the changes of apoptosis-related proteins, ASC phenotype-related proteins and HO-1, Nrf2, and so on were detected by Western blotting. Results: Allicin treatment could enhance the resistance of ASC to hemin toxicity, reduce the accumulation of reactive oxygen species caused by hemin, cell apoptosis and the expression of pro-apoptotic proteins, and also inhibit the expression of A1 phenotype protein C3 and up-regulate the expression of A2 phenotype protein S100 and TGF. In addition, allicin intervention could increase the expression of NRF2 and HO-1, and promote the nuclear translocation of Nrf2. Conclusion: Allicin could reduce oxidative stress and inflammatory response in an in vitro cerebral hemorrhage model, and exert neuroprotective effects.
[Key words]" intracerebral hemorrhage; astrocytes; allicin; inflammation; apoptosis; oxidative stress
脑出血是指由非外伤因素导致脑实质出血,又称自发性出血[1]。脑出血发病迅猛,可在较短时间内造成严重脑损伤,具有较高的发病率和死亡率[2-4]。脑出血引起的大脑损伤分为原发性损伤和继发性损伤两类[3];继发性神经损伤包括毒性反应、炎症反应和氧化应激损伤。脑出血发生后,从坏死和受损组织中释放出渗出性血液成分如补体、免疫球蛋白、凝血因子等以及血红蛋白的降解产物[5-6]。
星形胶质细胞(astrocyte,ASC)作为中枢神经系统中最主要的胶质细胞,广泛分布于脑组织神经细胞间和血管外间隙中,在调节神经元存活、递质代谢、信号传递以及维持血脑屏障等方面发挥重要作用[7]。当中枢神经系统受到创伤、缺血和发生神经退变时,为应对各种损伤,ASC从静息态变为反应性,这已成为中枢神经系统病变的病理标志[8-9]。反应性ASC包含多种亚群,在继发性神经损伤和组织修复中具有双重作用,基于基因表达的差异,可分为具有神经毒性的A1型和具有神经保护作用的A2型两种表型,A1型ASC表达C3标志蛋白,A2型ASC表达S100a10、转化生长因子-β(transforming growth factor-β,TGF-β)标志蛋白[10]。
大蒜素是大蒜中存在的主要生物活性化合物之一,属于一种有机含硫化合物,通过与蛋白质中的谷胱甘肽和巯基发生氧化还原反应从而发挥生物活性[11-12]。大蒜素及其产生的一系列具有生物活性的有机硫化合物在抗氧化、抗凋亡和免疫调节方面发挥作用,其多效性可能与其产生的蛋白质翻译后修饰有关[13-14]。此外,大蒜素通过与多种信号通路相互作用,减轻炎症反应所致的神经损伤并改善认知功能[12,15]。
大蒜素在缺血性脑卒中、蛛网膜下腔出血等神经性病变中表现出神经保护作用[15-16],但其在脑出血中的作用尚不清楚。因此,本研究拟通过建立模拟脑出血后继发性炎症反应和氧化应激损伤的细胞模型,观察大蒜素的作用并探讨其可能的作用机制。
1" 材料与方法
1.1" 材料
出生1~3 d的SD大鼠,雌、雄各5只,购自江苏大学实验动物中心,许可证编号:SCXK 2018-0012。
高糖型DMEM和青霉素链霉素购自美国Hyclone公司;大蒜素和血红素购于上海麦克林生化科技公司;CCK-8细胞计数试剂盒、一步法Tunel染色试剂盒、活性氧试剂盒和Annexin V-FITC细胞凋亡检测试剂盒购自上海碧云天生物公司;RIPA裂解缓冲液、PMSF、Cocktail和ECL发光液购自武汉Servicebio公司;DAPI购自美国Sigma公司;鼠抗Tubulin抗体、HRP标记的羊抗鼠、HRP标记的羊抗兔、488标记的羊抗兔二抗和Cy3标记的羊抗兔二抗购自武汉博士德生物公司;兔单抗半胱氨酸蛋白酶Caspase3、Cleaved-caspase3、蛋白激酶B(protein kinase B,PKB/Akt)、磷酸化PKB/Akt(phosphorylated PKB/Akt,p-PKB/p-Akt)、丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)、磷酸化MAPK(phosphorylated MAPK,p-MAPK)、核因子-κB(nuclear factor-kappaB,NF-κB)、NF-κB抑制蛋白(inhibitor of NF-κB,IκB)、磷酸化IκB(phosphorylated IκB,p-IκB)抗体购自美国CST公司;兔单抗血红素加氧酶-1(heme oxygenase-1,HO-1)、核因子E2相关因子2(nuclear factor erythroid 2-related factor 2,Nrf2)抗体购自美国Proteintech公司;补体3(complement 3,C3)、BCL-2相关X蛋白(BCL-2-associated X protein,Bax)抗体购自英国Abcam公司;兔单抗S100抗体购于美国Thermo公司;兔单抗TGF-β抗体购自美国Affinity Biosciences公司。
1.2" 方法
1.2.1" 原代ASC提取" 将新生的SD大鼠浸泡于75%乙醇中进行处死并消毒,随后置于超净工作台,剪开头皮并剥去颅骨,取出全脑。在预冷PBS中去除小脑,并将两侧大脑半球分离,在手术放大镜下剥除脑膜、血管和海马体。剩余的大脑皮质用0.25%胰蛋白酶消化10 min,并通过轻柔地吹打解离。于25 ℃行1 000×g离心7 min收集细胞沉淀,重悬于含90% DMEM、10%胎牛血清和100 U/mL青霉素链霉素的培养基中。将细胞置于含5% CO2的37 ℃细胞培养箱中,每3 d更换1次培养基。细胞融合后通过振荡摇落法清除上层小胶质细胞,采用免疫荧光染色验证ASC纯度。
1.2.2" ASC活力测定
1.2.2.1" 测定大蒜素对ASC活力的影响" 将ASC以5×103密度接种于96孔板,分为0、10、50、100、250、500、750、1 000 μmol/L大蒜素组,常规培养24 h,分别更换含有0、10、50、100、250、500、750、1 000 μmol/L大蒜素(用10%二甲基亚砜溶解)的无血清DMEM孵育24 h;将10 μL CCK-8溶液分别加至对应孔中,于37 ℃、5% CO2孵育1.5 h;采用酶标仪检测450 nm处光密度(D)值,选择最适大蒜素浓度进行后续实验。
1.2.2.2" 测定大蒜素抑制血红素导致的细胞毒性" 选择200 μmol/L大蒜素作为最适浓度,以不同浓度血红素处理ASC代表不同毒性强度。将ASC以5×103密度接种于两块96孔板,常规培养24 h;一块96孔板分别更换为含0、5、10、20、30、50 μmol/L血红素(用10 mmol/L NaOH溶解)的无血清DMEM孵育12 h;另一块96孔板先用含200 μmol/L大蒜素的常规DMEM孵育24 h,再分别更换为含0、5、10、20、30、50 μmol/L血红素的无血清DMEM孵育12 h;将10 μL CCK-8溶液分别加至对应孔中,于37 ℃、5% CO2孵育1.5 h;采用酶标仪检测450 nm处细胞D值。
1.2.3" 血红素诱导建立氧化应激细胞模型及大蒜素干预
1.2.3.1" 细胞分组" 选择血红素对ASC的半数致死量(25 μmol/L)进行实验。ASC分组如下:对照组,细胞常规培养24 h,更换DMEM孵育24 h,更换无血清DMEM孵育12 h;血红素组,细胞常规培养24 h,更换DMEM孵育24 h,更换含25 μmol/L血红素的无血清DMEM孵育12 h;溶剂对照组,细胞常规培养24 h,更换含10 mmol/L NaOH的DMEM孵育24 h,更换无血清DMEM孵育12 h;血红素+不同浓度大蒜素组,细胞常规培养24 h,分别更换含50、100、200 μmol/L大蒜素的无血清DMEM孵育24 h,更换含25 μmol/L血红素的DMEM孵育12 h。
1.2.3.2" Tunel染色法检测细胞凋亡" 将“1.2.3.1”分组处理ASC接种于无菌盖玻片,用0.3% Triton X-100于冰上处理细胞2 min;随后将细胞置于50 μL Tunel反应缓冲液中,37 ℃孵育1 h;用 DAPI(1∶5 000稀释)复染细胞核。采用正置荧光显微镜采集图像(德国Carl Zeiss公司)。
1.2.3.3" Annexin V-FITC检测细胞凋亡" 将“1.2.3.1”分组处理ASC以1×105密度接种至6孔板,根据Annexin V-FITC细胞凋亡试剂盒说明进行处理,20~25 ℃避光孵育10~20 min;于30 min内采用流式细胞仪(美国Beckman Coulter公司)进行检测。
1.2.3.4" 细胞活性氧测定" 将“1.2.3.1”分组处理ASC以5×105密度接种至6孔板,将细胞与10 μmol/L DCFH-DA避光孵育30 min;PBS洗涤2次;采用流式细胞仪或荧光显微镜(德国Carl Zeiss公司)检测荧光。
1.2.3.5" 蛋白质印迹测定凋亡相关蛋白表达水平" 将“1.2.3.1”分组处理ASC以5×105密度接种于6孔板,弃上清液并用添加PMSF和Cocktail的RIPA裂解缓冲液制备全细胞裂解物。配制10%凝胶,行60 V电泳,当Marker位置到达浓缩胶底部分界处时,将电压调为110 V继续电泳40 min;在冰浴中以300 mA恒流转膜60 min,将蛋白转至PVDF膜;5%脱脂牛奶于室温封闭2 h;4 ℃一抗孵育过夜,兔抗Bax抗体、Caspase3抗体、Cleaved-caspase3抗体、Akt抗体、p-Akt抗体、MAPK抗体、p-MAPK抗体以及内参β-微管蛋白,稀释比均为1∶1 000;TBST室温洗膜3次,每次5 min;二抗室温孵育60 min,稀释比为1∶5 000;TBST洗涤,在膜上滴加ECL溶液,采用Tanon图像系统(上海天能公司)进行显影。
1.2.4" 检测大蒜素干预对HO-1和Nrf2相对表达的影响
1.2.4.1" 细胞分组" ASC分组如下:对照组,ASC常规培养24 h,更换为常规DMEM孵育24 h;溶剂对照组,ASC常规培养24 h,更换为含二甲基亚砜的DMEM孵育24 h;不同浓度大蒜素组,ASC常规培养24 h,分别更换为含50、100、200 μmol/L大蒜素的DMEM孵育24 h。
1.2.4.2" 蛋白质印迹测定HO-1和Nrf2蛋白相对表达" 取“1.2.4.1”分组处理细胞,以5×105密度接种于6孔板,后续制备全细胞裂解物与蛋白质免疫印迹步骤同“1.2.3.5”,一抗兔抗HO-1、Nrf2及内参β-微管蛋白稀释比均为1∶1 000,二抗稀释比为1∶5 000。
1.2.5" 炎症因子诱导建立炎症反应细胞模型及大蒜素干预
1.2.5.1" 细胞分组" 将ASC进行如下分组:对照组,ASC常规培养24 h,更换DMEM孵育24 h,再次更换DMEM孵育24 h;炎症因子组,ASC常规培养24 h,更换DMEM孵育24 h,更换为含3 ng/mL IL-1α、30 ng/mL TNF-α、400 ng/mL C1q的DMEM孵育24 h;炎症因子+不同浓度大蒜素组,ASC常规培养24 h,分别更换为含50、100、200 μmol/L大蒜素的DMEM孵育24 h,更换为含3 ng/mL IL-1α、30 ng/mL TNF-α、400 ng/mL C1q的DMEM孵育24 h。
1.2.5.2" 蛋白质印迹测定ASC表型转化相关蛋白表达水平" 取“1.2.5.1”分组处理细胞,以5×105密度接种于6孔板,后续制备全细胞裂解物与蛋白质免疫印迹步骤同“1.2.3.5”,内参β-微管蛋白和兔抗C3抗体、S100a10抗体、兔抗TGF-β抗体、IκB抗体,稀释比均为1∶1 000,二抗稀释比为1∶5 000。
1.3" 统计分析
实验至少独立重复3次,数据以均数±标准差(x±s)表示。数据统计分析及统计表绘制采用GraphPad Prism 6.0。两组间比较采用独立样本t检验,多组间比较采用单因素方差分析,进一步两两比较采用Tukey多重比较法。Plt;0.05为差异有统计学意义。
2" 结果
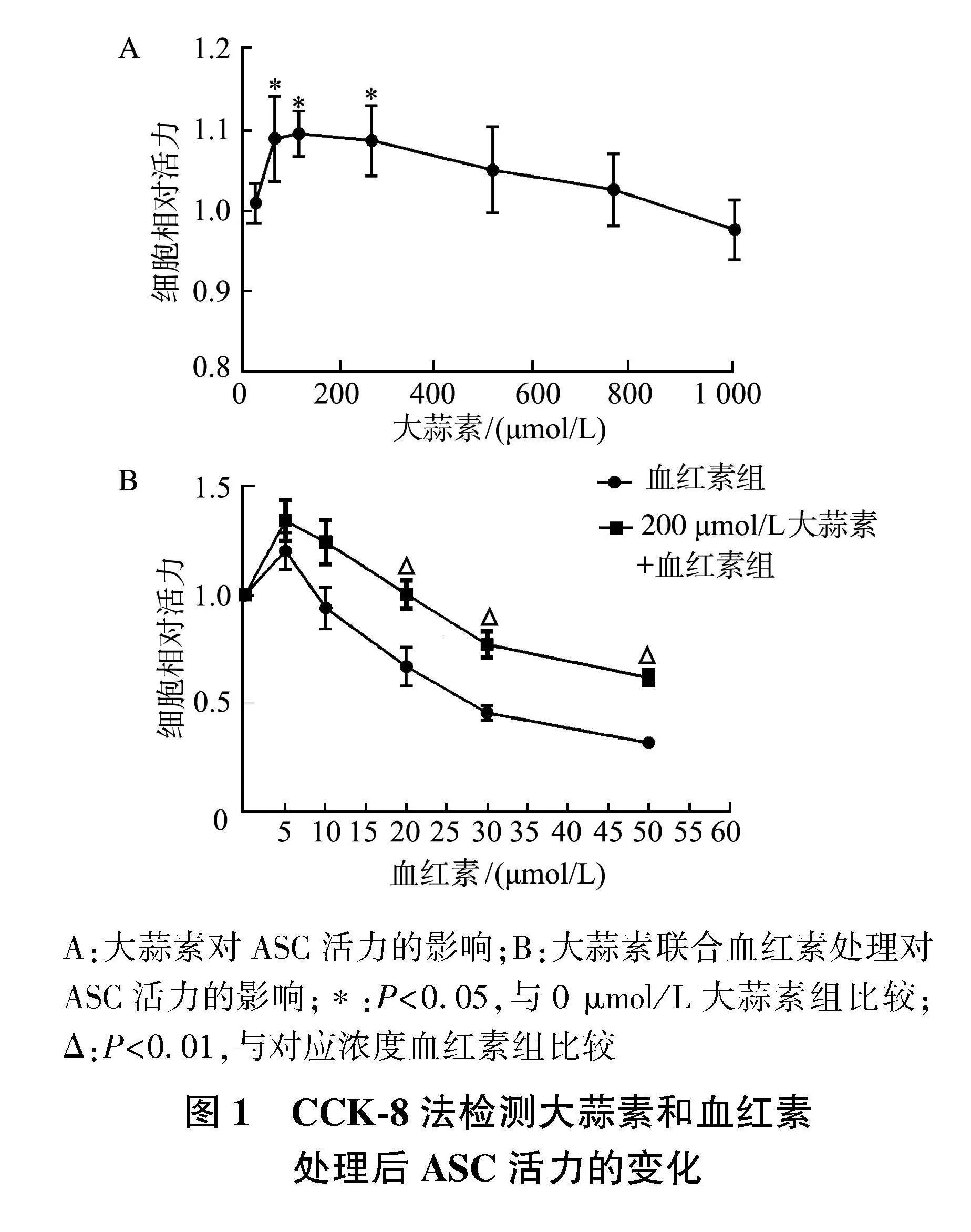
2.1" 大蒜素可抑制血红素对ASC的毒性
用不同浓度大蒜素处理细胞24 h,评估其对ASC的安全浓度。如图1A所示,与0 μmol/L大蒜素相比,50、100、250 μmol/L大蒜素组细胞活力显著上升(Plt;0.05),当大蒜素浓度高于250 μmol/L时,细胞活力呈下降趋势,但差异无统计学意义。因此,选择50、100、200 μmol/L大蒜素用于后续实验对比剂量区别;选择200 μmol/L大蒜素作为最适浓度。
如图1B显示,分别与对应浓度血红素组相比,200 μmol/L大蒜素+20、30、50 μmol/L血红素组细胞活力显著上升(Plt;0.01)。此外,血红素对ASC的半数致死量为25 μmol/L,因此选用该浓度用于后续实验。
A:大蒜素对ASC活力的影响;B:大蒜素联合血红素处理对ASC活力的影响;*:Plt;0.05,与0 μmol/L大蒜素组比较;Δ:Plt;0.01,与对应浓度血红素组比较
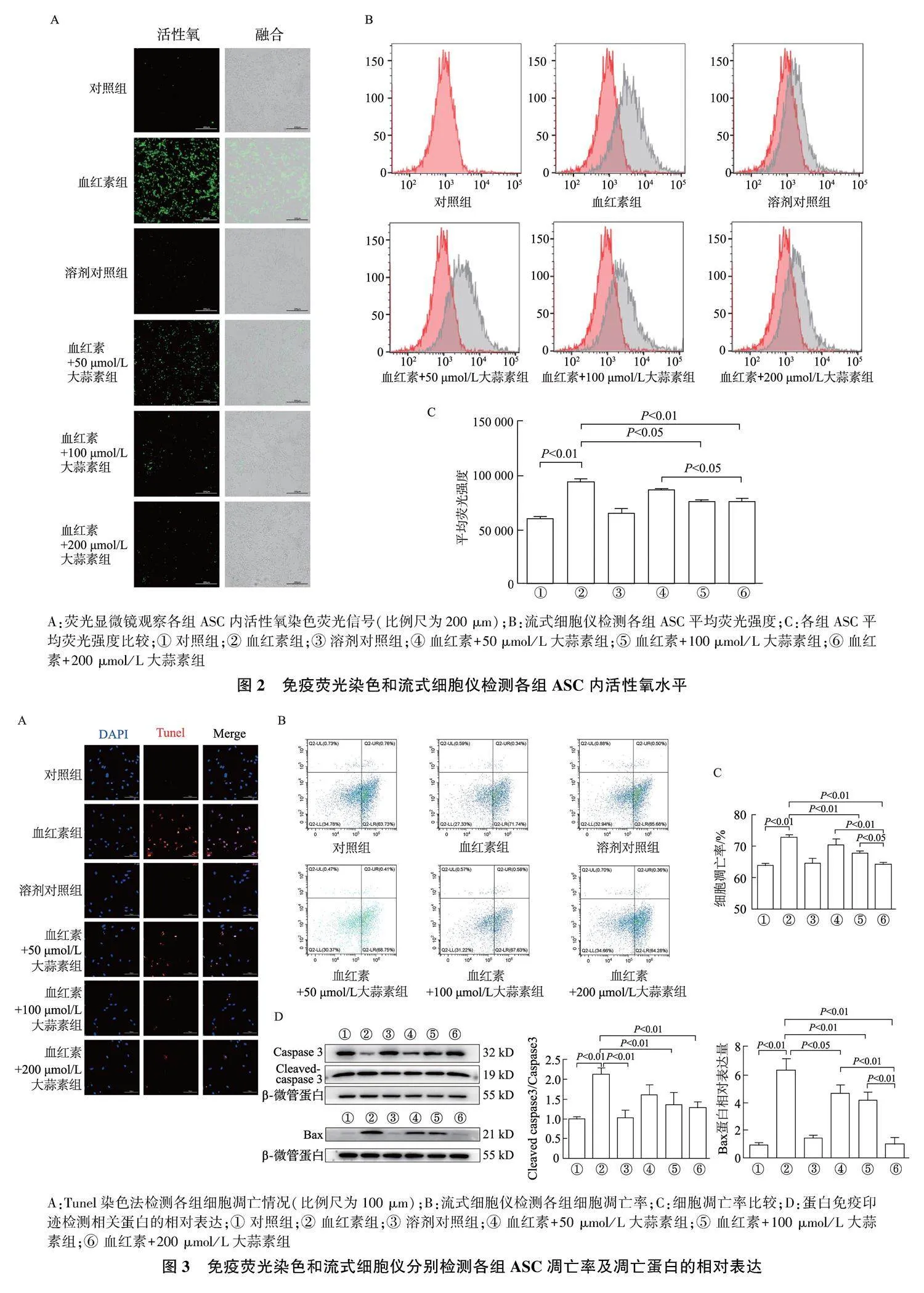
2.2" 大蒜素处理可抑制血红素引起的活性氧蓄积
免疫荧光染色结果显示,与对照组和溶剂对照组相比,血红素组细胞内绿色荧光信号增强,血红素+50、100、200 μmol/L大蒜素组细胞内绿色荧光信号减弱,见图2A。流式细胞仪分析结果显示,与对照组相比,血红素组平均荧光强度显著升高(Plt;0.01);与血红素组相比,血红素+100、200 μmol/L大蒜素组平均荧光强度显著降低(Plt;0.05或Plt;0.01),血红素+200 μmol/L大蒜素组平均荧光强度较血红素+50 μmol/L大蒜素组显著降低(Plt;0.05),见图2B、2C。
2.3" 大蒜素处理可抑制血红素引起的细胞凋亡
Tunel染色结果显示,与对照组相比,血红素组中代表凋亡细胞的红色荧光信号较强,大蒜素处理可以减弱细胞内红色荧光信号,见图3A。
流式细胞术结果显示,与对照组相比,血红素组细胞凋亡率显著升高(Plt;0.01);与血红素组相比,血红素+100、200 μmol/L大蒜素组细胞凋亡率均显著降低(P均lt;0.01),其中,血红素+200 μmol/L大蒜素组细胞凋亡率显著低于血红素+50 μmol/L大蒜素组和血红素+100 μmol/L大蒜素组(Plt;0.01或Plt;0.05)。见图3B、3C。
蛋白免疫印迹结果显示,与对照组相比较,血红素组中Cleaved caspase-3/caspase-3比值显著增加(Plt;0.01);与血红素组相比较,血红素+100、200 μmol/L大蒜素组中Cleaved caspase-3/caspase-3比值均显著下降(P均lt;0.01);与对照组相比,血红素组中促凋亡蛋白Bax表达量显著增加(Plt;0.01),与血红素组相比,血红素+50、100、200 μmol/L大蒜素组中Bax蛋白表达均显著减低(Plt;0.05或Plt;0.01),血红素+200 μmol/L大蒜素组中Bax蛋白表达显著低于血红素+50 μmol/L大蒜素组和血红素+100 μmol/L大蒜素组(P均lt;0.01)。见图3D。
2.4" 大蒜素预处理可上调Nrf2/HO-1表达
蛋白免疫印迹结果显示,与对照组相比,100、200 μmol/L大蒜素组中HO-1表达量显著增加(P均lt;0.05);且200 μmol/L大蒜素组中HO-1表达量明显高于50 μmol/L大蒜素组(Plt;0.05),见图4A。与对照组相比,100、200 μmol/L大蒜素组中Nrf2表达量显著增加(P均lt;0.01),100、200 μmol/L大蒜素组中Nrf2表达量均显著高于50 μmol/大蒜素组(P均lt;0.05),见图4B。
A:荧光显微镜观察各组ASC内活性氧染色荧光信号(比例尺为200 μm);B:流式细胞仪检测各组ASC平均荧光强度;C:各组ASC平均荧光强度比较;① 对照组;② 血红素组;③ 溶剂对照组;④ 血红素+50 μmol/L大蒜素组;⑤ 血红素+100 μmol/L大蒜素组;⑥ 血红素+200 μmol/L大蒜素组
A:Tunel染色法检测各组细胞凋亡情况(比例尺为100 μm);B:流式细胞仪检测各组细胞凋亡率;C:细胞凋亡率比较;D:蛋白免疫印迹检测相关蛋白的相对表达;① 对照组;② 血红素组;③ 溶剂对照组;④ 血红素+50 μmol/L大蒜素组;⑤ 血红素+100 μmol/L大蒜素组;⑥ 血红素+200 μmol/L大蒜素组
A:各组ASC中HO-1蛋白相对表达水平比较;B:各组ASC中Nrf2蛋白相对表达水平比较;① 对照组;② 溶剂对照组;③ 50 μmol/L大蒜素组;④ 100 μmol/L大蒜素组;⑤ 200 μmol/L大蒜素组
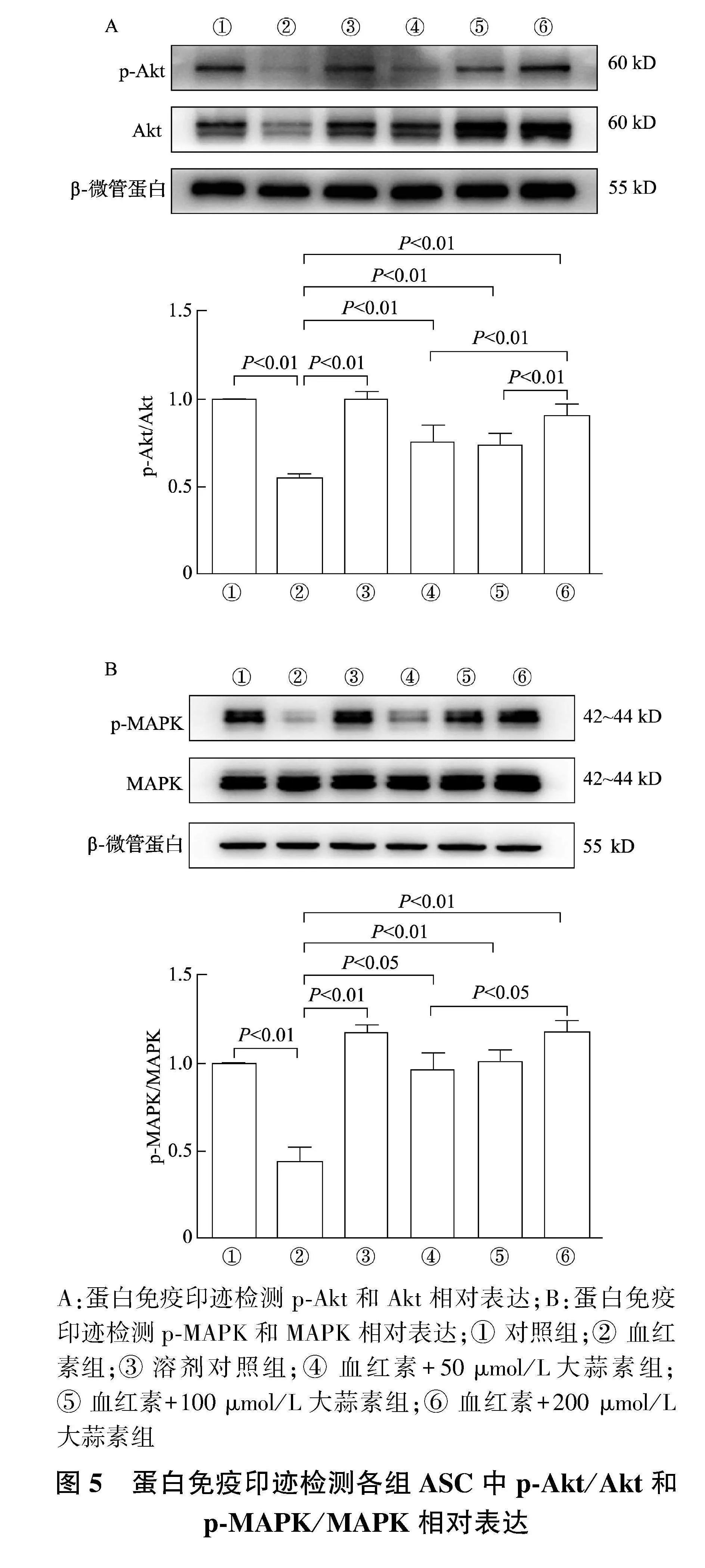
2.5" 大蒜素预处理可上调p-Akt/Akt和p-MAPK/MAPK比值
如图5A、5B所示,与对照组相比,血红素组p-Akt/Akt比值显著减低(Plt;0.01);与血红素组相比,血红素+50、100、200 μmol/L大蒜素组p-Akt/Akt比值显著上调(P均lt;0.01),其中,血红素+200 μmol/L大蒜素组比值显著高于血红素+50、100 μmol/L大蒜素组(P均lt;0.01)。
如图5C、5D所示,与对照组相比,血红素组p-MAPK/MAPK比值显著降低(Plt;0.01);与血红素组相比,血红素+50、100、200 μmol/L大蒜素组p-MAPK/MAPK比值均显著升高(Plt;0.05或Plt;0.01),其中,血红素+200 μmol/L大蒜素组显著高于血红素+50 μmol/L大蒜素组(Plt;0.05)。
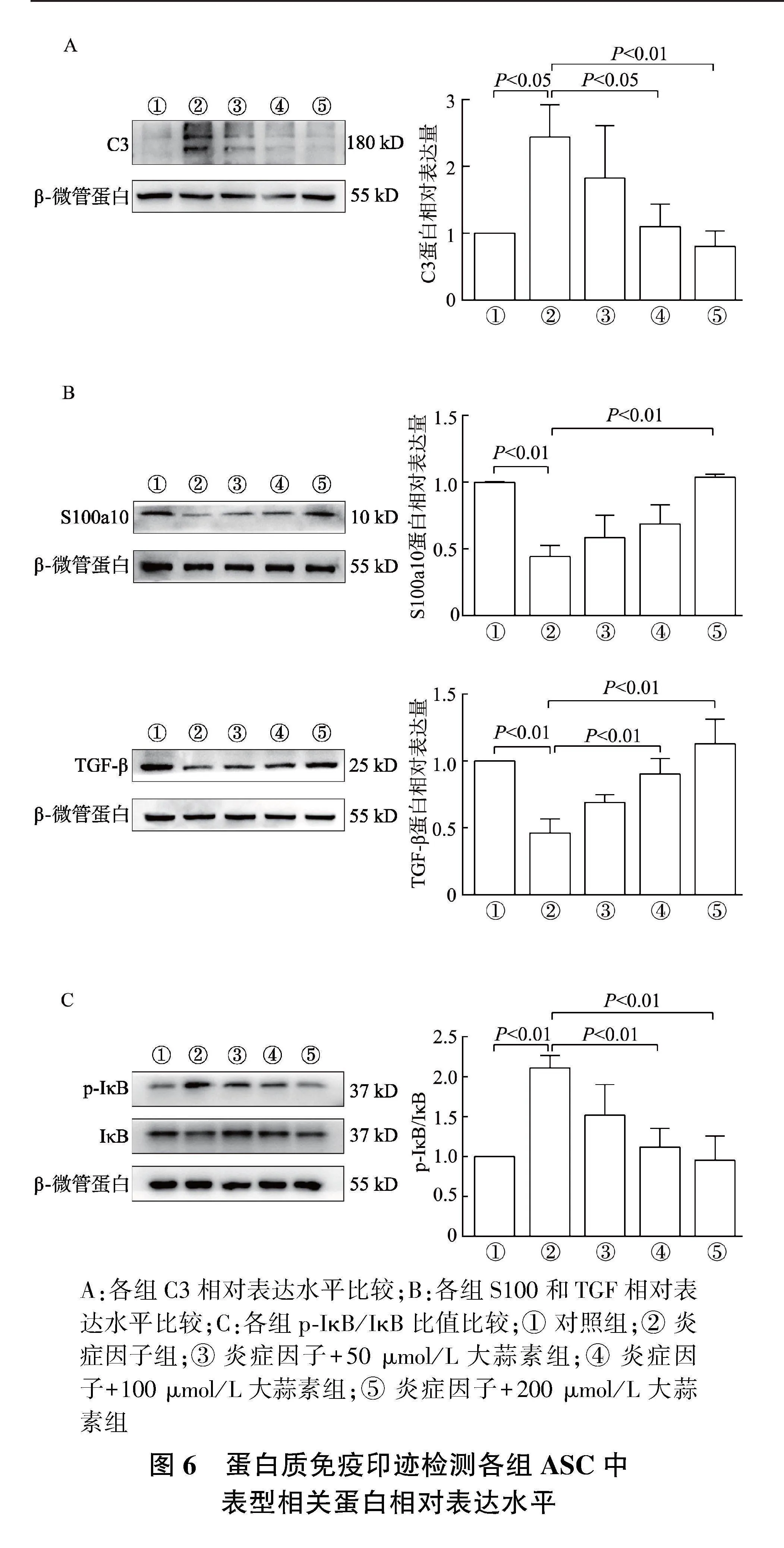
2.6" 大蒜素预处理下调ASC A1表型蛋白表达,上调A2表型蛋白表达
如图6A所示,与对照组相比,炎症因子组C3相对表达明显增加(Plt;0.05),与炎症因子组相比,炎症因子+100、200 μmol/L大蒜素组C3相对表达显著下降(Plt;0.05或Plt;0.01)。
如图6B所示,与对照组相比,炎症因子组中TGF-β相对表达显著下降(Plt;0.01);与炎症因子组相比,炎症因子+100、200 μmol/L大蒜素组中TGF-β相对表达显著增加(Plt;0.01);与对照组相比,炎症因子组中S100a10相对表达明显下降(Plt;0.01),与炎症因子组相比,炎症因子+200 μmol/L大蒜素组中S100a10相对表达明显上升(Plt;0.01)。
如图6C所示,与对照组相比,炎症因子组p-IκB表达量增多,p-IκB/IκB比值明显升高(Plt;0.01);与炎症因子组相比,炎症因子+100、200 μmol/L大蒜素组p-IκB表达量减少,p-IκB/IκB比值明显下降(P均lt;0.01)。
A:蛋白免疫印迹检测p-Akt和Akt相对表达;B:蛋白免疫印迹检测p-MAPK和MAPK相对表达;① 对照组;② 血红素组;③ 溶剂对照组;④ 血红素+50 μmol/L大蒜素组;⑤ 血红素+100 μmol/L大蒜素组;⑥ 血红素+200 μmol/L大蒜素组
A:各组C3相对表达水平比较;B:各组S100和TGF相对表达水平比较;C:各组p-IκB/IκB比值比较;① 对照组;② 炎症因子组;③ 炎症因子+50 μmol/L大蒜素组;④ 炎症因子+100 μmol/L大蒜素组;⑤ 炎症因子+200 μmol/L大蒜素组
3" 讨论
考虑到脑出血后血红蛋白引起的继发性神经损伤是病情恶化的关键因素,参考本课题组之前的研究[17],本实验采用血红素诱导的方法构建脑出血后继发性毒性损伤模型,选用炎症因子IL-1、TNF-α、C1q作为诱导剂,构建稳定的A1型ASC模型。相比采用凝血酶在体外模拟脑出血的方法,此法简单、快捷、重复性好,消除了凝血酶浓度差异所致的不稳定因素对实验结果产生的影响[18-19]。本研究结果显示,血红素显著增加了ASC内的活性氧含量和细胞凋亡,大蒜素可增强ASC对血红素毒性的抵抗,减少血红素引起的活性氧蓄积,减少细胞凋亡;炎症因子可激活ASC形成A1表型,而大蒜素抑制其引起的A1极化。对比不同浓度大蒜素对ASC保护作用的差异,结果表明,在ASC活性氧水平、HO-1和Nrf2蛋白相对表达水平分析中,200 μmol/L大蒜素处理组指标显著优于50 μmol/L大蒜素组;在ASC凋亡率、Bax、p-Akt/Akt相对表达水平分析中,200 μmol/L大蒜素组指标显著优于100 μmol/L和50 μmol/L大蒜素组;在p-MAPK/MAPK相对表达水平分析中,200 μmol/L大蒜素组显著优于50 μmol/L大蒜素组。由此说明,随着大蒜素浓度升高,作用效果越明显,在50 μmol/L和200 μmol/L处理浓度的对比中,呈一定的浓度依赖性。
HO-1是Nrf2下游基因,可将有毒的血红素降解为胆红素,其表达增加是机体对氧化损伤的一种适应性保护反应[20]。研究表明,激活Nrf2/HO-1通路有助于减弱血肿周围的损伤级联反应,并减轻脑出血后的脑损伤和神经功能缺损[21-22]。本实验结果显示,大蒜素处理可致ASC中Nrf2和HO-1蛋白相对表达量增加,这可能是大蒜素帮助ASC抵御氧化应激损伤的机制,且200 μmol/L大蒜素组指标优于50 μmol/L大蒜素组,呈一定的浓度依赖性。
PI3K/Akt信号通路活化可促进或抑制神经细胞内多种蛋白磷酸化,调控神经细胞存活、增殖、分化、自噬及调控神经细胞突触可塑性等,与神经系统疾病的发生密切相关[23]。针对药物作用机制的研究发现,TREM2和CCR4能够通过促进Akt磷酸化发挥对脑出血后损伤的保护效应[24-25]。MAPK途径在细胞凋亡、炎症和应激中起重要作用;MAPK的磷酸化激活是一种防御机制,与促凋亡蛋白的下调和抗凋亡蛋白的上调密切相关,有助于减轻神经元损伤所产生的有害影响,在一些神经疾病模型中具有神经保护作用[26-27]。本研究结果显示,大蒜素处理可促进Akt磷酸化,且200 μmol/L大蒜素组优于100、50 μmol/L大蒜素组;此外,大蒜素处理可促进MAPK磷酸化,且200 μmol/L大蒜素组显著优于50 μmol/L大蒜素组,这与大蒜素抗细胞凋亡作用相一致。由此推测,大蒜素在血红素神经毒性模型中对ASC的保护作用可能与其促进Akt和MAPK磷酸化有关。
A1型ASC介导的免疫级联反应在脑出血诱导的脑损伤和修复进展中起关键作用[28-29]。A1型ASC上调补体成分C3表达,其作用包括分泌神经毒素,抑制突触形成和诱导神经元快速死亡等[30]。A1型ASC的活化过程与NF-κB通路密切相关[31-32]。A2型ASC则上调一些神经营养因子如S100a10、TGF-β表达,参与神经元存活、生长与分化[33-34]。将抗炎药物注入神经损伤部位后,可抑制该部位A1型反应性ASC的活化,从而使神经功能得到改善与恢复[35-36]。尽早干预A1型反应性ASC转化的关键机制,如抑制I-κB磷酸化,有助于抑制A1表型的产生[37]。本研究进一步探究了I-κB磷酸化情况,结果表明大蒜素处理可下调I-κB磷酸化水平。由此表明,大蒜素可能通过抑制I-κB磷酸化的方式,从而逆转由炎症因子诱导的ASC A1极化现象。
综上所述,本研究表明大蒜素在模拟脑出血环境的氧化应激损伤和炎症反应细胞模型中具有良好的神经保护作用,其保护作用与剂量有关,呈一定的浓度依赖性。然而,本研究尚未探讨时间效应对大蒜素保护作用的影响。此外,大蒜素在脑出血体内模型中的具体保护作用机制有待后续进一步探讨。
[参考文献]
[1]" 中华医学会神经外科学分会, 中国医师协会急诊医师分会, 国家卫生和计划生育委员会脑卒中筛查与防治工程委员会. 自发性脑出血诊断治疗中国多学科专家共识[J]. 中华神经外科杂志, 2015, 31(12): 1189-1194.
[2]" Xiao M, Li Q, Feng H, et al. Neural vascular mechanism for the cerebral blood flow autoregulation after hemorrhagic stroke[J]. Neural Plast, 2017, 2017: 5819514.
[3]" Fang Y, Tian Y, Huang Q, et al. Deficiency of TREK-1 potassium channel exacerbates blood-brain barrier damage and neuroinflammation after intracerebral hemorrhage in mice[J]. J Neuroinflammation, 2019, 16(1): 96.
[4]" Urday S, Kimberly WT, Beslow LA, et al. Targeting secondary injury in intracerebral haemorrhage—perihaematomal oedema[J]. Nat Rev Neurol, 2015, 11(2): 111-122.
[5]" Clark JF, Loftspring M, Wurster WL, et al. Bilirubin oxidation products, oxidative stress, and intracerebral hemorrhage[J]. Acta Neurochir Suppl, 2008, 105: 7-12.
[6]" Keep RF, Hua Y, Xi G. Intracerebral haemorrhage: mechanisms of injury and therapeutic targets[J]. Lancet Neurol, 2012, 11(8): 720-731.
[7]" Sofroniew MV, Vinters HV. Astrocytes: biology and pathology[J]. Acta Neuropathol, 2010, 119(1): 7-35.
[8]" Escartin C, Guillemaud O, Carrillo-de Sauvage MA. Questions and (some) answers on reactive astrocytes[J]. Glia, 2019, 67(12): 2221-2247.
[9]nbsp; Anderson MA, Ao Y, Sofroniew MV. Heterogeneity of reactive astrocytes[J]. Neurosci Lett, 2014, 565: 23-29.
[10]" Zamanian JL, Xu L, Foo LC, et al. Genomic analysis of reactive astrogliosis[J]. J Neurosci, 2012, 32(18): 6391-6410.
[11]" Subramanian MS, Nandagopal Ms G, Amin Nordin S, et al. Prevailing knowledge on the bioavailability and biological activities of sulphur compounds from alliums: A potential drug candidate[J]. Molecules, 2020, 25(18): 4111.
[12]" Nadeem M, Kazmi I, Ullah I, et al. Allicin, an antioxidant and neuroprotective agent, ameliorates cognitive impairment[J]. Antioxidants (Basel), 2021, 11(1): 87.
[13]" Mocayar Marón FJ, Camargo AB, Manucha W. Allicin pharmacology: Common molecular mechanisms against neuroinflammation and cardiovascular diseases[J]. Life Sci, 2020, 249: 117513.
[14]" Gruhlke MCH, Antelmann H, Bernhardt J, et al. The human allicin-proteome: S-thioallylation of proteins by the garlic defence substance allicin and its biological effects[J]. Free Radic Biol Med, 2019, 131: 144-153.
[15]" Zhuang F, Shi X, Qiao S, et al. Allicin promotes functional recovery in ischemic stroke via glutathione peroxidase-1 activation of Src-Akt-Erk[J]. Cell Death Discov, 2023, 9(1): 335.
[16]" Shao J, Wu Q, Lv SY, et al. Allicin attenuates early brain injury after experimental subarachnoid hemorrhage in rats[J]. J Clin Neurosci, 2019, 63: 202-208.
[17]" 陆聪, 桂如林, 吕广钊, 等. Connexin43敲降对血红素诱导的星形胶质细胞凋亡和铁死亡的影响[J]. 江苏大学学报(医学版), 2023, 33(1): 29-36.
[18]" Yang M, Gao F, Liu H, et al. Hyperosmotic induction of aquaporin expression in rat astrocytes through a different MAPK pathway[J]. J Cell Biochem, 2013, 114(1): 111-119.
[19]" Yamamoto N, Yoneda K, Asai K, et al. Alterations in the expression of the AQP family in cultured rat astrocytes during hypoxia and reoxygenation[J]. Brain Res Mol Brain Res, 2001, 90(1): 26-38.
[20]" Naito Y, Takagi T, Higashimura Y. Heme oxygenase-1 and anti-inflammatory M2 macrophages[J]. Arch Biochem Biophys, 2014, 564: 83-88.
[21]" Wang D, Peng X, Yang A, et al. Edaravone promotes nerve function recovery after acute cerebral infarction in rats via targeting Keap1-Nrf2/ARE[J]. Panminerva Med, 2021, 63(3): 384-385.
[22]" Chen X, Liang H, Xi Z, et al. BM-MSC transplantation alleviates intracerebral hemorrhage-induced brain injury, promotes astrocytes vimentin expression, and enhances astrocytes antioxidation via the Cx43/Nrf2/HO-1 Axis[J]. Front Cell Dev Biol, 2020, 8: 302.
[23]" Chen S, Peng J, Sherchan P, et al. TREM2 activation attenuates neuroinflammation and neuronal apoptosis via PI3K/Akt pathway after intracerebral hemorrhage in mice[J]. J Neuroinflammation, 2020, 17(1): 168.
[24]" Deng S, Jin P, Sherchan P, et al. Recombinant CCL17-dependent CCR4 activation alleviates neuroinflammation and neuronal apoptosis through the PI3K/AKT/Foxo1 signaling pathway after ICH in mice[J]. J Neuroinflammation, 2021, 18(1): 62.
[25]" Sun Y, Liu WZ, Liu T, et al. Signaling pathway of MAPK/ERK in cell proliferation, differentiation, migration, senescence and apoptosis[J]. J Recept Signal Transduct Res, 2015, 35(6): 600-604.
[26]" Jia L, Liu J, Song Z, et al. Berberine suppresses amyloid-beta-induced inflammatory response in microglia by inhibiting nuclear factor-kappaB and mitogen-activated protein kinase signalling pathways[J]. J Pharm Pharmacol, 2012, 64(10): 1510-1521.
[27]" Lin X, Zhang N. Berberine: Pathways to protect neurons[J]. Phytother Res, 2018, 32(8): 1501-1510.
[28]" Hara M, Kobayakawa K, Ohkawa Y, et al. Interaction of reactive astrocytes with type Ⅰ collagen induces astrocytic scar formation through the integrin-N-cadherin pathway after spinal cord injury[J]. Nat Med, 2017, 23(7): 818-828.
[29]" Lan X, Han X, Li Q, et al. Modulators of microglialactivation and polarization after intracerebral haemorrhage[J]. Nat Rev Neurol, 2017, 13(7): 420-433.
[30]" Chistyakov D, Gavrish GE, Goriainov SV, et al. Oxylipin profiles as functional characteristics of acute inflammatory responses in astrocytes pre-treated with IL-4, IL-10, or LPS[J]. Int J Mol Sci, 2020, 21(5): 1780.
[31]" Liddelow SA, Guttenplan KA, Clarke LE, et al. Neurotoxic reactive astrocytes are induced by activated microglia[J]. Nature, 2017, 541(7638): 481-487.
[32]" Lian H, Yang L, Cole A, et al. NFκB-activated astroglial release of complement C3 compromises neuronal morphology and function associated with Alzheimer′s disease[J]. Neuron, 2015, 85(1): 101-115.
[33]" Kowiański P, Lietzau G, Czuba E, et al. BDNF: A key factor with multipotent impact on brain signaling and synaptic plasticity[J]. Cell Mol Neurobiol, 2018, 38(3): 579-593.
[34]" Xu X, Zhang A, Zhu Y, et al. MFG-E8 reverses microglial-induced neurotoxic astrocyte (A1) via NF-κB and PI3K-Akt pathways[J]. J Cell Physiol, 2018, 234(1): 904-914.
[35]" Vismara I, Papa S, Veneruso V, et al. Selective modulation of A1 astrocytes by drug-loaded nano-structured gel in spinal cord injury[J]. ACS Nano, 2020, 14(1): 360-371.
[36]" Liu W, Wang Y, Gong F, et al. Exosomes derived from bone mesenchymal stem cells repair traumatic spinal cord injury by suppressing the activation of A1 neurotoxic reactive astrocytes[J]. J Neurotrauma, 2019, 36(3): 469-484.
[37]" Wang L, Pei S, Han L, et al. Mesenchymal stem cell-derived exosomes reduce A1 astrocytes via downregulation of phosphorylated NFκB P65 subunit in spinal cord injury[J]. Cell Physiol Biochem, 2018, 50(4): 1535-1559.
[收稿日期]" 2023-05-22" [编辑]" 刘星星
猜你喜欢
中国典型病例大全(2022年11期)2022-05-13 21:44:11
世界科学技术-中医药现代化(2020年2期)2020-07-25 02:05:56
老年医学与保健(2017年6期)2017-02-06 05:30:02
中国现代医生(2016年27期)2016-12-21 16:05:30
中国医药导报(2016年25期)2016-11-30 07:20:06
中国实用医药(2016年7期)2016-03-17 14:04:02
科技视界(2016年5期)2016-02-22 19:03:28
西南军医(2016年6期)2016-01-23 02:21:19
中国卫生标准管理(2015年5期)2016-01-14 05:17:05
西南军医(2015年2期)2015-01-22 09:09:37