
鱼类颜色视觉的研究进展
2024-05-29 03:45:42郭建磊杨德发薛金刚宋明元徐鹏翔
安徽农业科学 2024年8期
关键词:行为
郭建磊 杨德发 薛金刚 宋明元 徐鹏翔
摘要 视觉器官对于大多数鱼类来说是一个非常重要的感觉器官,鱼眼不仅能感觉光的敏感和颜色,而且还能感知物体的形状、大小、运动等。鱼类的视觉功能与摄食、集群、生殖、防御等行为方面具有密切联系,并发挥重要作用。为了研究鱼类颜色视觉能力进展,以目前国际主要的数据库(Science Direct、JSTOR、Wiley Online Library、SpringerLink、ProQuest等)为数据源,采用文献计量分析法,研究了鱼类颜色视觉能力的不同研究方法的发展与现状,从而更深入地了解鱼类视觉特征,可为今后在渔船作业中改变灯光颜色以提高对鱼群诱集的效果、在工厂化养殖过程中控制养殖环境的颜色以及光强来提高养殖效益、在控制帮助鱼类洄游以保护洄游鱼种等提供参考依据。
关键词 鱼类颜色视觉;行为;电生理;视蛋白
中图分类号 S917.4文献标识码 A文章编号 0517-6611(2024)08-0010-05
doi:10.3969/j.issn.0517-6611.2024.08.003
Bibliometric Study on Fish Color Vision
GUO Jian-lei1,YANG De-fa2,XUE Jin-gang2 et al
(1.Marine Ranching Engineering Technology Research Center of Liaoning,Dalian Ocean University,Dalian,Liaoning 116023;2.Molinot Super Marine Supplies Co.,Ltd.,Dalian, Liaoning 116104)
Abstract The vision orgon is a very important sensory organ for fish. Fish eyes can not only sense light sensitivity and color, but also perceive the shape, size, and movement of objects. The visual function of fish has a close connection and important role in feeding, clustering, reproduction, defense and other behavioral aspects. To study the progress of color vision ability in fish, this paper investigates the development and status of different research methods on color vision ability in fish using the current major international databases (Science Direct, JSTOR, Wiley Online Library, SpringerLink, ProQuest, etc.) as data sources and uses bibliometric analysis to study the development and current status of different research methods on fish color vision ability. This study was conducted to understand the visual characteristics of fish in a more in-depth way, which can provide a reference for future methods such as changing the color of lights in fishing vessels to improve the effect of fish attraction, controlling the color and light intensity of the breeding environment to improve the breeding efficiency in the process of factory farming, and controlling the help of fish migration to protect migratory fish species.
Key words Fish color vision;Behavior;Electrophysiology;Opsin
視觉器官对于大多数鱼类来说是一个非常重要的感觉器官。鱼类的眼睛结构类似于陆生脊椎动物,比如鸟类和哺乳动物,但它却有着独特的球状晶状体。大多数鱼类具有颜色视觉,视网膜上的感光细胞有2种,可区分颜色的视锥细胞(cone)和感受明暗的视杆细胞(rod)。感光色素(photopigment or visual pigment)有4种,视紫红质位于视杆细胞上,其对弱光敏感,不能区分颜色。其余位于视锥细胞上,3种视锥细胞包含不同的视紫蓝质分子,绿视锥细胞为450~675 nm(绿光),蓝视锥细胞为455 nm(蓝光);红视锥细胞为625 nm(橙色光)。所有感光色素均具有视蛋白(opsin),不同的视蛋白具有不同的光谱吸收率[1],有些鱼对紫外线[2]和一些偏振光较为敏感[3]。
该研究以目前国际主要的数据库(ScienceDirect、JSTOR、Wiley Online Library、SpringerLink、ProQuest等,以及依托Google Scholar补充未被国际主要数据库收录的其他文献来源)为数据源,采用文献计量分析法,研究了鱼类颜色视觉能力的不同研究方法的发展与现状,为今后在渔船作业中改变灯光颜色以提高对鱼群诱集的效果、在工厂化养殖过程中控制养殖环境的颜色以及光强来提高养殖效益、在控制帮助鱼类洄游以保护洄游鱼种等提供参考依据。
1 研究现状
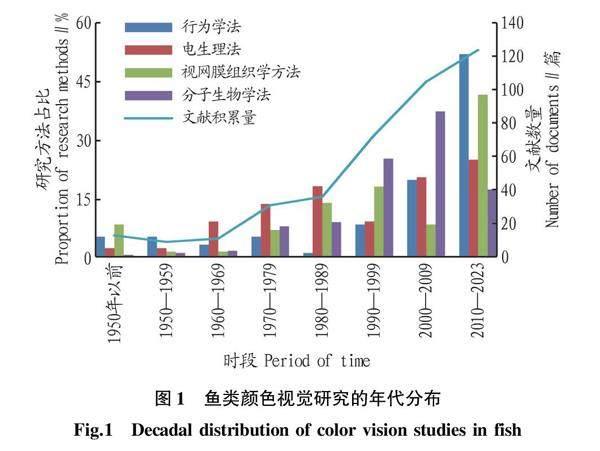
20世纪80—90年代是鱼类颜色视觉电生理研究的鼎盛时期;从20世纪90年代末至今,行为学方法、视网膜组织学研究法和分子生物学视蛋白研究法在同时发展,其各个方法的研究结果也基本相似,并共同证明了鱼类颜色视觉能力,以及在水产养殖业和捕捞业中的应用前景(图1)。然而,鱼类视觉的真正表现还需要结合行为反应观察来进一步阐述。行为反应的准确度,基本可以真实地反映鱼类的视觉能力,因为其结果与电生理学和解剖学反映的视觉系统相接近[4]。
因此,科研工作者分别从行为心理学(behavioristic psychology)、电生理学(electrophysiology)、视网膜组织学(retinal histology)、分子生物学(molecular biology)等方面开展研究。
2 行为学法
2.1 无条件反射法
利用鱼对视觉刺激的本能行为反应测量鱼的探查和区分能力。其中最有效的方法是,随着重复刺激次数的增加,行为表现没有衰减的刺激方法[4]。
2.2 条件驯化法
2.2.1
应答条件作用。这种方法被称作巴普洛夫(pavlovian)或经典条件作用法(classical conditioning methods)。通过对鱼类进行训练,建立条件反射机制,用来观察研究某一刺激与鱼类的感觉能力和行为的关系。
2.2.2
早期的行为研究。1884年,Graber[5]首次使用行为学方法,通过设置不同试验对照组,每组提供2种灯光环境供试验鱼自行选择,验证了泥鳅科须鳅属(Barbatula)和欧白鱼属(Alburnus)具有区分光亮和辨别颜色的能力。其试验鱼的偏好结果为:白>黑(明>暗)、红光>绿光>蓝光>紫外线光。在此基础上,1906年,Washburn等[6]用有饵料的红色医疗钳和无饵料的绿色医疗钳喂食黑斑须雅罗鱼(Semotilus atromaculatus)证明其可以区分颜色,并在行为学方法上提出神经学上的“关联记忆”现象。1919年,Reeves[7]使用Yerkes-Watson辨别箱(the Yerkes-Watson discrimination box)研究了鱼类颜色视觉。在1966年,Muntz等[8]用二选一迫使选择法(two-choice procedure)方法研究了金鱼(Carassius auratus)辨别颜色的能力。
20世纪50—60年代,日本开始发展有关鱼类颜色视觉在捕捞中的应用研究。黑木敏郎等[9-11]、草下孝也[12]分别分析了红、绿、蓝、黄、白等不同颜色的水上集鱼灯的波长分布。在此基础上,長峯嘉之[13]通过海上试验,验证了波长在460~620 nm的不同颜色集鱼灯对日本鯷(Engraulis japonicus)、竹荚鱼(Trachurus japonicus)、沙丁鱼(Sardina pilchardus)、秋刀鱼(Cololabis saira)的诱集效果。其结果为:绿白色、蓝色最佳;白、粉红色较好;黄、蓝白色一般;红色无效果。
从20世纪70年代末起,中国科研工作者开始围绕灯光捕捞、养殖、人工渔礁等方面展开鱼类颜色视觉行为研究。
在海水鱼类方面。1979年,俞文钊等[14]通过水槽行为试验,观察了蓝圆鲹(Decapterus maruadsi)和鲐鱼(Pneumatophorus japonicus)对等能光谱色的趋光反应。证明蓝圆鲹幼鱼、成鱼对蓝、绿色光的趋光率最高,对红光趋光率最低。鲐鱼对紫光和红光有最大的趋光率。罗会明等[15]通过观察水槽中鳗鲡(Anguilla japonica)幼苗对不同光色的趋光性,认为鳗鲡幼苗具有辨别光色的能力,并且随着颜色光亮度的变化而发生变化。1980年,何大仁等[16]通过水槽试验和海上观察认为,孔沙丁鱼(Sardinella perforate Cantor)和勃氏银汉鱼(Atherina bleekeri Günther)對强光区中较高波长的橙、绿光趋光率高。2007年,方金等[17]研究了养殖大黄鱼的趋光性发现大黄鱼对光源照度为300 lx的蓝光或200 lx的绿光有一定的正趋向性。2013年,林超等[18]对褐菖鲉(Sebasticus marmoratus)和日本黄姑鱼(Argyrosomus japonicus)的研究表明,在自然组的礁区聚集率高于有光组,礁区聚集率为橙光组<绿光组<红光组<蓝光组<白光组<自然组,褐菖鮋总体上表现出一定的负趋光性。配置不同光色的人工鱼礁模型对日本黄姑鱼的诱集效果与自然组无明显差异。2018年,黄六一等[19]研究了光照对花鲈(Lateolabrax japonicus)行为反应的影响,结果显示,红光、蓝光和黄光对花鲈有一定的吸引作用,而红光的吸引效果最为明显,红光的光强也存在一定的耐受限度,即红光对花鲈的吸引效果随着光强的增加而增大,到达饱和值后光强增加吸引度反而降低,研究结果可以为海洋牧场中花鲈的增殖放养控制鱼群提供技术参考。2019年,莫伟均等[20]研究花鳗鲡(Anguilla marmorata)幼鱼的趋光性,结果显示,花鳗鲡幼鱼对光的趋向性由其发育阶段决定,幼鱼尤爱红光、黄光和紫光,在花鳗鲡幼鱼洄游的鳗鱼道入口采用灯光诱鱼帮助其找到生长洄游通道。2020年,张志强等[21]在研究欧洲舌齿鲈(Dicentrarchus labrax)对养殖水池背景颜色的偏好中发现幼鱼在黑色等深色区域累计停留时间最长,表现出偏好性。
在淡水鱼类方面。王小平等[22]在玻璃水族箱里从趋光性、选择性、条件反射和呼吸频率等角度研究了鲢鱼(Hypophthalmichthys molitrix)、鲫鱼(Carassius auratus)、罗非鱼(Oreochromis spp.)、乌鳢(Ophiocephalus argus)、黄鳝(Monopterus albus)的颜色视觉,在对红、绿、蓝三色光的选择上,除了鲢鱼偏好红光外,其他试验鱼均偏好绿光。2001年,魏开建等[23-24]采用光梯度法研究了鳜鱼(Siniperca chuatsi)在发育过程中对6种光的趋光特性。鳜鱼在不同照度下的适宜光色不尽相同,随其生长,弱光区的适宜光色为短波段的蓝、绿光,而强光区的适宜光色由长波段的黄、橙、红光移至短波段的绿光。2008年,许传才等[25]研究了不同颜色光对鲤的诱集效果。鲤的最大平均趋集率分别为白光61.0%、红光45.0%、蓝光42.0%、绿光27.1%。2012年,肖炜等[26]证明奥利亚罗非鱼(Oreochromis aureus)对不同单色光具有选择性,在蓝光、绿光下的趋光率高于红光、黄光。2016年,巩建华等[27]研究了红鲫鱼(Carassius auratus)对黑、蓝、红、绿、橙、黄6种颜色光的趋向性选择反应和在不同的光强度下的趋光反应。红鲫鱼对6种颜色的选择有明显的差异为黑色>蓝色>红色>绿色>橙色>黄色,夜晚红鲫鱼对颜色选择的优先顺序与白天相同,有正趋光行为。2019年,李若琳等[28]对鲫鱼(Carassius auratus)幼鱼进行颜色标记并测量SMR(standard metabolic rate)和形态参数,用以测定颜色标记对鲫鱼标准的代谢效率是否有影响。孙春艳等[29] 研究锦鲫(Carassius auratus)和中华倒刺鲃(Spinibarbus sinensis)对颜色标记反应的差异,发现体外颜色标记对锦鲫和中华倒刺鲃的有氧运动能力均无影响,表明体外颜色标记法可较好地运用于鱼类群体行为学的相关研究。刘飞等[30-31]分别对红色草金鱼(Carassius auratus red var.)和红白鲫(Carassius auratus)进行了背景色选择的研究。红色草金鱼相对于白色背景来说更偏好于彩色(红、绿、蓝)背景,而在彩色背景选择中会更倾向于喜欢红色和绿色。而红白鲫在群体条件下更偏好于橙色和黄色,不喜欢红色。以上研究结论在开发高效诱捕灯、提高工厂化养殖效益等方面具有积极意义。
随着20世纪90年代末水产养殖业的蓬勃发展,一些关于不同颜色光照对鱼类摄食行为影响的研究也开展了起来。Tamazouzt等[32]研究水槽壁颜色和不同光照等级对欧亚鲈鱼(Perca fluviatilis)幼鱼生长和成活率的影响。White等[33]研究了紫外线光谱对孔雀鱼(Poecilia reticulata)摄食的影响。Monk等[34]验证了不同颜色的水槽壁对大西洋鳕鱼(Gadus morhua)幼鱼的生长、存活率和摄食行为的影响。Villamizar等[35]研究了不同光谱及光照对欧洲舌齿鲈(Dicentrarchus labrax)幼鱼生长、发育和成活率的影响。2013年,高橋明義等[36]阐述了特定波长光在鱼类养殖中的应用前景,并提出今后的课题是研发LED(light emitting diode)养殖用灯。2015年,Mesquita等[37]利用颜色驯化鲤鱼摄食。
2010年至今,随着油价上涨,如何选择既能满足生产需要,又能节省油耗的光源,成为灯光围网捕捞中的关键问题。LED具有能环保、安全可控、光谱稳定等优点,作为新一代的高效节能照明产品备受瞩目,科研工作者们相继开展了新型LED集鱼灯的研发[38-43]以及鱼类对其行为反应研究。陈清香等[44]通过水槽试验和海上试验证实,白光LED灯对蓝圆鲹的光诱效果显著优于蓝紫光LED灯;白光LED灯对竹筴鱼的光诱效果极显著优于蓝紫光LED。2019年,黄宇雯[45]研究了在黑暗、白光(荧光灯)以及由LED产生的红光、绿光和紫光环境条件下对黄金鲈(Perca flavescens)生长表现、性腺发育和应激状态的影响和雌雄个体的差异。
3 电生理方法
3.1 视网膜电图法(electroretinogram,ECG)
通过对鱼类视觉系统周边与中枢各级神经元的电活动进行记录,精确的分析鱼类视系统的光谱敏感性、适应性和色觉等问题,从而为行为学研究提供重要依据。
3.2 视顶盖诱发电位法(visually evoked tectum potential,VETP)
视顶盖是视神经传入纤维的基本收集器,由间脑和中脑某些核,以及从视网膜所获得的直接神经支配。因此,可以通过记录鱼类视觉中枢的电活动来分析神经细胞的活动,从而更深入了解鱼类的视觉特性。
在20世纪50年代末60年代初,微电极实验技术的发展和鱼类电生理研究新阶段紧密联系,其中相当大部分是对视觉系统各部分单独神经元特性的研究[46-49],其中包括借助光点和活动有形刺激对鱼感受野的研究。
自20世纪70年代以来,中国在这方面做了大量的工作。1977年,杨雄里等[50]对蓝圆鲹和鲐鱼的视网膜电图进行了研究,发现其b波对缺氧很敏感,并且视网膜电图具有典型的混合型视网膜特征。杨雄里等[50]、李震元等[51]分别测得了鲐鱼视顶盖诱发电位的适应特性和光谱敏感性。梁旭方等[52]用电生理方法测得鱖鱼的光谱敏感曲线峰值为绿光,同时鱖鱼对长波光有较高光谱敏感性,不同发育时期的幼鱼适宜颜色光与其光谱敏感曲线峰值不完全一致,说明鳜鱼成鱼无辨色能力,幼鱼可能具有一定的颜色视觉。王艺磊等[53]测得真鲷(Pagrosomus major)视网膜电图结果显示,真鲷存在2种以上的光感受系统,且具备变色能力。2020年,Maximova等[54]研究鱼类视网膜中具有持续活性的神经节细胞及其在视觉场景评价中的可能作用。鱼的视神经顶盖轴突末端记录到单个神经节细胞的细胞外突刺活动。持续响应的ON和OFF单元(USR)的接收野大小为4°~5°,与特征检测器的接收野大小相当。USR参与颜色辨别。神经节细胞同时被分离成ON和OFF通道,可为视顶盖神经元提供控制外界注意所需的视觉场景信息。
4 视网膜组织学方法
1940年,Caspersson[55-56]首次将显微镜和分光光度法结合,设计了第一台显微分光光谱仪,并创立了显微分光光度法(microspectrophotometry,MSP法)。
20世纪50—70年代,国外开始采用生化提取、MSP等方法对鱼类的视色素进行测定[57-60],分析鱼类颜色视觉能力。而国内对此方面的研究起步较晚。陈明等[61-62]对蓝圆鲹、鲐鱼和5种淡水鱼类的杆细胞视色素进行测定,并采用部分漂白的方法分析了它们的光谱差别,发现青鱼(Mylopharyngodon piceus)、草鱼(Ctenopharyngodon idellus)、鲢鱼(Hypophthalmichthys molitrix)和鳙鱼(Aristichthys nobilis)均属于网膜醛1色素,光谱吸收峰值分别为530、528、525和527 nm;鲐鱼为网膜酸2色素,光谱吸收峰值为500 nm;蓝圆鲹有2种视紫红,光谱吸收峰值分别为488、510 nm;罗非鱼含有网膜醛1和酸2的混合色素,光谱吸收峰值分别为500、522 nm。1995年,郑微云等[63]通过电镜切片组织学法证实了黑鲷(Sparus macrocephalus)视网膜中存在视杆、单锥和孪生双锥3种光感受细胞。并测得视杆色素为视紫红质VP4851,其吸收光谱峰值为485 nm,与黑鲷栖息水层中日光能量分布相对应,也与该水层中的生物光场相适应。2014年,李超等[64]运用组织学方法,对铠平鲉(Sebastes hubbsi)、花斑平鲉(Sebastes nigricans)、大泷六线鱼(Hexagrammos otakii)和斑头六线鱼(Hexagrammos agrammus)的性成熟个体视网膜光感受细胞和最小分辨角进行了比较研究。2016年,车景青等[65]对大菱鲆(Scophthalmus maximus)鱼苗视網膜组织结构与视觉特征的研究。大菱鲆变态发育前视网膜视敏度高、光敏度低,变态发育后感光系统退化视敏度降低、光敏度增强以适应底栖生活。2017年,牛亚兵[66]对玫瑰高原鳅(Triplophysa rosa)和贝氏高原鳅(Triplophysa bleekeri)视觉器官进行比较组织学研究,发现玫瑰高原鳅因长期生活在黑暗的环境中,为适应洞穴生活,其视网膜结构与贝氏高原鳅相比发生了退化。2019年,Luehrmann等[67]研究天竺鲷科鱼类的视觉色素多样性,发现其既能很好地适应昏暗的光线条件,又能保持复杂的色觉。de Busserolles等[68]研究夜间活动珊瑚礁鱼科尾斑棘鳞鱼(Sargocentron caudimaculatum)的视网膜结构和分析其视觉生态,发现它们为了适应夜间的生活方式,视网膜结构都以棒状为主,这些棒状的视网膜被排列成库,会增加眼睛的光敏度,帮助其在昏暗的环境下进行颜色判断。
5 分子生物学方法
视蛋白是具有7个跨膜结构域的G蛋白耦联受体超家族(G protein-coupled receptors,GPCR)中的一个庞大的家族。根据其序列相似性及产物λmax的不同,脊椎动物的视蛋白可分为5个亚家族:①RH1,又称为视杆蛋白(rhodopsin),其 λmax约500 nm,在视杆细胞中表达,介导暗视觉,无色觉功能;②RH2(视紫质样蛋白基因2,rhodopsin-like pigments),其λmax在 470~510 nm 范围内;③SWS(短波长敏感性视蛋白,short wavelength-ensitive pigments),其λmax在360~460 nm 范围内;④LWS(
长波长敏感性视蛋白,long wavelength-sensitive),其λmax在 510~560 nm 范围内;RH2 、SWS、LWS视蛋白均在视锥细胞里表达且介导明视觉[69]。在不同的物种和生活环境中视蛋白基因的长度和基因结构组成也是不同的,因此,可以根据不同视蛋白各自所占的比例来反映鱼类颜色视觉能力。
从20世纪90年代至今,国外对鱼类视蛋白的研究主要涉及视蛋白的种类、视蛋白的表达量、视蛋白与环境之间的关系等方面。Lythgoe等[70]对大堡礁笛鲷属(Lutjanus)的视色素生态学研究指出,被研究的笛鲷属双锥区视色素的吸收光谱与其栖息地水质的吸收光谱相近。单锥区有一个蓝光或紫外光敏感视色素。视杆细胞的视色素出现少许变异,其最大吸收峰的波长在 489~502 nm 范围内。此后,鱼类视觉系统与环境之间的关系的研究逐渐增加。Yokoyama[71]指出随着生物所处环境的不同,视杆细胞和视锥细胞的比例也不同;深海夜行性的动物为了尽量感受可用的弱光,视网膜上几乎全是视杆细胞。Yokoyama[72]的研究指出矛尾鱼(Latimeria chalumnae)的RH2变为478 nm,这可能和其生活的环境有关。非洲矛尾鱼生活在约200 m深的海洋中,在这个深度能到达的光线波长约480 nm。Yokoyama等[73]对金鳞鱼(Ostichthys japonicus)的RH1基因进化进行了研究,结果表明 RH1基因的λmax值与其栖息的水环境有密切的关系:随着水层深度的不同,其最大吸收峰值 λmax分别发生了变化。周慧[74]在研究四指马鲅(Eleutheronema tetradactylum)视网膜发育对不同光周期环境的适应性研究中,对RH1、LWS、RH2、SWS2视蛋白基因序列和氨基酸序列进行生物信息学分析,运用荧光定量PCR研究视蛋白在不同的光照周期下表达量的变化。Musilova等[75]和Torres-Dowdall等[76]都对视蛋白的opsin基因的表达进行了研究,深水鱼类蓝色敏感(SWS2A)、绿色敏感(RH2A)和红色敏感(LWS)opsin基因的表达存在较大的差异,浅水鱼类都以红色敏感(LWS)opsin基因为主。这种视网膜视蛋白基因的表达的变化导致视觉敏感性的差异,代表了对不同颜色光暗度环境的适应。
6 结语
无论溪流、湖泊、沼泽、河口、珊瑚礁、大洋还是深海,鱼类生存的各类型的环境皆有其特殊的环境因子,使得各种环境中光谱的组成有很大的差别,各类鱼种随着栖息地的环境变化发展出不同环境所需要的颜色视觉:
①在浅水域或是清澈表层水域的鱼种大多数拥有蓝、绿、红光及紫外光的颜色视觉。
②居住在能见度较差的河口区,则促使该地区的鱼种强化其在红光区的感受力使得视觉向长波长光谱区段偏移。
③深海的鱼种,则受到随海水的深度增加,水吸收了除了蓝光以外的其他光的物理特性的影响,使得视觉系统丧失了看见绿色及红色的能力,但却特化成可以感受低照度蓝光,以帮助其在微弱光照环境下生活。
综上所述,近年来国内对鱼类颜色视觉的研究发展很快,一些成果在生产中得到了推广应用,但与国外研究水平相比,还有很大差距。从试验方法和研究手段上,室内工作多,实际观察和调研较少,使得鱼類在实验室中对光的反应与自然条件下有一定差别。常用的光梯度法无法测定鱼类色觉以及视敏度等指标,因此许多实验技术尚待改进。从研究广度上看,国内研究的鱼种类还不够丰富,一些重要的经济鱼类尚未进行研究,光诱捕种类的范围有待提高。从研究深度上看,除了对某几种鱼如蓝圆鲹、鲐鱼等的研究较深入以外,对其他鱼类仅限于某一方面,而缺乏系统性的研究。鱼类对颜色光的反应是一个条件反射与非条件反射交错的过程,因此要把行为生理学、电生理学与组织学、分子生物学等联系起来综合研究全面分析,使结论更加精确。由于国内对该方面的研究还存在着不足,目前处于设想和摸索阶段,因此,未来还要开展大量研究工作,在引进国外现代化的研究手段的同时,还应大力推广在生产实践中的应用,例如不同颜色的LED灯在养殖业以及捕捞业中的应用,为我国的渔业生产提供强大的技术支持。
参考文献
[1] BOWMAKER J K,LOEW E R.Vision in fish[M]//MASLAND R H,ALBRIGHT T D,DALLOS P,et al.The senses:A comprehensive reference.Amsterdam:Elsevier,2008:53-76.
[2] DUNLAP W C,WILLIAMS D M,CHALKER B E,et al.Biochemical photoadaptation in vision:U.V.-absorbing pigments in fish eye tissues[J].Comparative biochemistry and physiology part B:Comparative biochemistry,1989,93(3):601-607.
[3] WATERMAN T H,FORWARD R B.Field evidence for polarized light sensitivity in the fish Zenarchopterus[J].Nature,1970,228:85-87.
[4] NORTHMORE D P M,YAGER D.Psychophysical methods for investigations of vision in fishes[M]//ALI M A.Vision in fishes.Boston,MA:Springer,1975:689-704.
[5] GRABER V.Grundlinien zur Erforschung des Helligkeits-und Farbensinnes der Tiere[M].Prag:F.Tempsky,1884.
[6] WASHBURN M F,BENTLEY I M.The establishment of an association involving color-discrimination in the creek chub,Semotilus atromaculatus[J].Journal of comparative neurology and psychology,1906,16(2):113-125.
[7] REEVES C D.Discrimination of light of different wave-lengths by fish[M].New York:H.Holt&Company,1919.
[8] MUNTZ W R,CRONLY-DILLON J R.Colour discrimination in goldfish[J].Animal behaviour,1966,14(2/3):351-355.
[9] 黒木敏郎,中馬三千雄.漁業用忌避灯光の研究 I.振廻し光束について[J].日水誌,1953,18:26-29.
[10] 黒木敏郎,中山博.集魚用螢光色灯の研究:第I報:各色光の波長分布並に白熱灯との比較[J].鹿児島大学水産学部紀要,1958,6:95-98.
[11] 黒木敏郎,中山博,上之清尚.集魚用螢光管色灯の研究-2-[J].北海道大学水産学部研究彙報,1964,14(4):215-235.
[12] 草下孝也.白熱灯及び螢光水銀灯の集魚効果と水中照度[J].日本水産学会誌,1959,25(1):17-21.
[13] 長峯嘉之.色けい光ランプの集魚効果について[J].照明学会雑誌,1967,51(9):528-530.
[14] 俞文钊,何大仁,郑玉水.兰圆鲹、鲐鱼对等能光谱色的趋光反应[J].厦门大学学报(自然科学版),1979,18(2):126-130.
[15] 罗会明,郑微云.鳗鲡幼鱼对颜色光的趋光反应[J].淡水渔业,1979(8):9-16.
[16] 何大仁,罗会明,郑美丽.孔沙丁鱼和勃氏银汉鱼趋光特性的研究:对弥散性白光的反应及对等照度光谱色的适应过程[J].厦门大学学报(自然科学版),1980,19(2):81-88.
[17] 方金,宋利明,蔡厚才,等.网箱养殖大黄鱼对颜色和光强的行为反应[J].上海水产大学学报,2007,16(3):269-274.
[18] 林超,桂福坤.不同光色下人工鱼礁模型对褐菖鲉和日本黄姑鱼诱集效果试验[J].渔业现代化,2013,40(2):66-70,75.
[19] 黄六一,徐基强,陈婧,等.光照对花鲈行为反应的影响研究[J].渔业信息与战略,2018,33(1):45-50.
[20] 莫伟均,何贞俊,杨聿,等.基于鳗鱼道诱鱼技术研究花鳗鲡幼鱼光色趋向性[J].应用生态学报,2019,30(6):2109-2115.
[21] 張志强,马宾,张磊,等.欧洲舌齿鲈幼鱼对养殖水池背景色的偏好研究[J].大连海洋大学学报,2020,35(6):901-907.
[22] 王小平,袁传宓.五种淡水鱼类的颜色视觉初探[J].淡水渔业,1991(1):14-16.
[23] 魏开建,张海明,张桂蓉.鳜鱼苗在光场中反应行为的初步研究[J].水利渔业,2001,21(1):4-6.
[24] 魏开建,张桂蓉,张海明.鳜鱼不同生长阶段中趋光特性的研究[J].华中农业大学学报,2001,20(2):164-168.
[25] 许传才,伊善辉,陈勇.不同颜色的光对鲤的诱集效果[J].大连水产学院学报,2008,23(1):20-23.
[26] 肖炜,李大宇,杨弘,等.奥利亚罗非鱼在光场中的行为反应研究[J].中国农学通报,2012,28(26):105-109.
[27] 巩建华,郭春阳,田喆,等.红鲫鱼对环境颜色及光强的趋向性研究[J].生物学杂志,2016,33(5):13-18.
[28] 李若琳,唐晴晴,王萌,等.颜色标记对鲫标准代谢率的影响[J].重庆师范大学学报(自然科学版),2019,36(5):60-65.
[29] 孙春艳,曾令清.颜色标记对两种鱼类有氧运动能力的影响[J].重庆师范大学学报(自然科学版),2020,37(3):67-71.
[30] 刘飞,李斌,王凡.群体条件下红色草金鱼对不同颜色背景的选择偏好[J].河北渔业,2020(9):22-25.
[31] 刘飞,张家旺,张武伟.红白鲫对不同背景颜色的选择偏好性研究[J].水生态学杂志,2019,40(5):73-77.
[32] TAMAZOUZT L,CHATAIN B,FONTAINE P.Tank wall colour and light level affect growth and survival of Eurasian perch larvae(Perca fluviatilis L.)[J].Aquaculture,2000,182(1/2):85-90.
[33] WHITE E M,CHURCH S C,WILLOUGHBY L J,et al.Spectral irradiance and foraging efficiency in the guppy,Poecilia reticulata[J].Animal behaviour,2005,69(3):519-527.
[34] MONK J,PUVANENDRAN V,BROWN J A.Does different tank bottom colour affect the growth,survival and foraging behaviour of Atlantic cod(Gadus morhua)larvae?[J].Aquaculture,2008,277(3/4):197-202.
[35] VILLAMIZAR N,GARCA-ALCAZAR A,SANCHEZ-VAZQUEZ F J.Effect of light spectrum and photoperiod on the growth,development and survival of European sea bass(Dicentrarchus labrax)larvae[J].Aquaculture,2009,292(1/2):80-86.
[36] 高橋明義,水澤寛太,古藤澄久,等.II-4.特定波長光を活用した魚類養殖技術の開発[J].日本水産學会誌,2013,79(5):885.
[37] MESQUITA F O,BORCATO F L,HUNTINGFORD F A.A conditioned response overrides social attraction in common carp:A possibility for low stress sorting strongly schooling fish?[J].Applied animal behaviour science,2015,163:188-193.
[38] 龚雅萍.LED诱捕灯的设计与应用研究[J].浙江海洋学院学报(自然科学版),2012,31(4):371-373.
[39] 钱卫国,陈新军,钱雪龙,等.300W型LED集鱼灯光学特性及其节能效果分析[J].海洋渔业,2011,33(1):99-105.
[40] 李天华.LED水下集鱼灯的研究与设计探讨[J].渔业现代化,2010,37(3):64-67.
[41] 熊正烨,谭中明,梁苏,等.用于水下集鱼灯的LED发光板研究[J].农业科技与装备,2014(7):28-30.
[42] 安樂和彦,遠藤将一,松岡達郎,等.I-1.魚類による光の受容と集魚灯設計への応用[J].日本水産学会誌,2013,79(5):879-880.
[43] SHEN S C,HUANG H J,CHAO C C,et al.Design and analysis of a high-intensity LED lighting module for underwater illumination[J].Applied ocean research,2013,39:89-96.
[44] 陈清香,熊正烨,谭中明,等.2种LED灯光诱蓝圆鲹和竹筴鱼的渔获比较[J].南方水产科学,2013,9(3):80-84.
[45] 黄宇雯.黄金鲈(Perca flavescens)在不同LED光谱下的光色偏好及生理响应的性别差异[D].金华:浙江师范大学,2019.
[46] HERTER K.Die Fischdressuren und ihre sinnesphysiologischen grundlagen[M].Berlin:Akademie-Verlag,1953.
[47] BERNSTEIN J J.Role of the telencephalon in color vision of fish[J].Experimental neurology,1962,6:173-185.
[48] ARORA H L,SPERRY R W.Color discrimination after optic nerve regeneration in the fish Astronotus ocellatus[J].Developmental biology,1963,7:234-243.
[49] TAMURA T,NIWA H.Spectral sensitivity and color vision of fish as indicated by S-potential[J].Comparative biochemistry and physiology,1967,22(3):745-754.
[50] 杨雄里,李震元,潘家模,等.海水鱼趋光特性的电生理研究Ⅱ.蓝圆鲹、鲐鱼视网膜电图b波的光谱敏感性[J].科学通报,1977,22(3):135-137.
[51] 李震元,杨雄里,黄玉霖,等.海水鱼趋光特性的电生理研究-Ⅲ.鲐鱼视顶盖诱发电位的适应特性和光谱敏感性[J].科学通报,1977,22(Z1):217-218,199.
[52] 梁旭方,郑微云,王艺磊.鳜鱼视觉特性及其对捕食习性适应的研究Ⅰ.视网膜电图光谱敏感性和适应特性[J].水生生物学报,1994,18(3):247-253.
[53] 王艺磊,张子平,郑微云.真鲷视网膜结构及视觉特性研究-Ⅲ.真鲷视觉特性[J].热带海洋学报,1994,13(2):75-81.
[54] MAXIMOVA E M,ALIPER A T,DAMJANOVIC' I Z,et al.Ganglion cells with sustained activity in the fish retina and their possible function in evaluation of visual scenes[J].Neuroscience and behavioral physiology,2020,51(1):123-133.
[55] CASPERSSON T.II.-Methods foe the determination of the absorption spectra of cell structures[J].Journal of microscopy,1940,60(1/2):8-25.
[56] LANGWORTHY O R.Cell growth and cell function:A cytochemical study[J].American journal of psychiatry,1951,108(2):154.
[57] MUNK O.Ocular anatomy of some deep-sea teleosts [R].1966.
[58] MUNTZ W R,NORTHMORE D P.Vision and visual pigments in a fish,Scardinius erythrophthalmus(the rudd)[J].Vision research,1970,10(4):281-298.
[59] WOLKEN J J.Photoprocesses,photoreceptors,and evolution[M].New York:Academic Press,1977.
[60] MCFARLAND W N,MUNZ F W.Part III:The evolution of photopic visual pigments in fishes[J].Vision Res,1975,15(10):1071-1080.
[61] 陈明,史丽烈.两种海水鱼的视紫红的吸收光谱:蓝圆鲹(Decapterus maruadsi)和鲐鱼(Pneumatophorus japonicus)[J].生理学报,1979,31(2):193-196.
[62] 史丽烈,陈明.青(Mylopharyngodon piceus)、草(Ctenopharyngodon idellus)、鲢(Hypophthalmichthys molitrix)、鳙(Aristichthys nobilis)和罗非鱼(Tilapia mossambica)的视色素[J].生理学报,1980,32(2):176-180.
[63] 郑微云,李伦平,薛雄志,等.黑鲷视网膜结构与视觉特性研究[J].热带海洋,1995,14(2):53-59.
[64] 李超,王亮,覃乐政,等.4种岩礁性鱼类视网膜感光细胞和最小分辨角的比较[J].水产学报,2014,38(3):400-409.
[65] 车景青,陈京华,胡苗峰.大菱鲆(Scophthalmus maximus)鱼苗视网膜组织结构与视觉特性[J].渔业科学进展,2016,37(2):25-32.
[66] 牛亚兵.玫瑰高原鳅和贝氏高原鳅视觉器官比较组织学研究[D].重庆:西南大学,2017.
[67] LUEHRMANN M,CARLETON K L,CORTESI F,et al.Cardinalfishes(Apogonidae)show visual system adaptations typical of nocturnally and diurnally active fish[J].Molecular ecology,2019,28(12):3025-3041.
[68] DE BUSSEROLLES F,CORTESI F,FOGG L,et al.The visual ecology of Holocentridae,a nocturnal coral reef fish family with a deep-sea-like multibank retina[J].Journal of experimental biology,2021,224:1-16.
[69] 刘旦.笛鲷属鱼类长波段视蛋白(LWS)基因序列与进化的比较分析[D].湛江:广东海洋大学,2012.
[70] LYTHGOE J N,MUNTZ W R A,PARTRIDGE J C,et al.The ecology of the visual pigments of snappers(Lutjanidae)on the Great Barrier Reef[J].Journal of comparative physiology A,1994,174(4):461-467.
[71] YOKOYAMA S.Molecular evolution of vertebrate visual pigments[J].Progress in retinal and eye research,2000,19(4):385-419.
[72] YOKOYAMA S.Molecular evolution of color vision in vertebrates[J].Gene,2002,300(1/2):69-78.
[73] YOKOYAMA S,TAKENAKA N.The molecular basis of adaptive evolution of squirrelfish rhodopsins[J].Mol Biol Evol,2004,21(11):2071-2078.
[74] 周慧.四指马鲅视网膜早期发育及其对不同光周期环境的适应性研究[D].上海:上海海洋大学,2017.
[75] MUSILOVA Z,INDERMAUR A,BITJA-NYOM A R,et al.Evolution of the visual sensory system in cichlid fishes from crater lake Barombi Mbo in Cameroon[J].Molecular ecology,2019,28(23):5010-5031.
[76] TORRES-DOWDALL J,KARAGIC N,HRER A,et al.Diversity in visual sensitivity across Neotropical cichlid fishes via differential expression and intraretinal variation of opsin genes[J].Molecular ecology,2021,30(8):1880-1891.
基金項目 大连市科技创新基金项目(2021JJ11CG001)。
作者简介 郭建磊(1999—),男,山东聊城人,硕士研究生,研究方向:捕捞学、被动声学。*通信作者,实验师,从事鱼类行为学、渔具渔法研究。
收稿日期 2023-05-29
猜你喜欢
中学课程辅导·教师教育(上、下)(2016年17期)2016-12-17 18:17:51
武术研究(2016年7期)2016-12-15 09:35:45
东方教育(2016年4期)2016-12-14 09:19:49
文艺生活·中旬刊(2016年11期)2016-12-13 20:09:15
现代法学(2016年6期)2016-12-08 17:29:53
职工法律天地·下半月(2016年10期)2016-11-30 11:34:40
医学信息(2016年30期)2016-11-28 20:46:18
商(2016年33期)2016-11-24 20:47:47
上海医药(2016年20期)2016-11-09 21:22:20
文艺生活·中旬刊(2016年9期)2016-11-07 00:55:44
