
库姆塔格沙漠可培养放线菌多样性及抑菌潜力评估
2024-05-18 19:02李佳慧汶瑛田茂刘光琇张威章高森薛林贵
中国抗生素杂志 2024年4期
李佳慧 汶瑛 田茂 刘光琇 张威 章高森 薛林贵


摘要:沙漠土壤蕴藏丰富的放线菌资源,尤其是稀有放线菌类群,是挖掘新放线菌资源的理想生境。本研究开展库姆塔格沙漠可培养放线菌抑菌活性筛选,以期为该地区放线菌资源开发和利用提供菌株资源。根据分离菌株的16S rRNA基因序列分析放线菌多样性;从库姆塔格沙漠4份土样中分离纯化到370株放线菌,分布于11个目12个科的22个属,其中链霉菌属和糖丝菌属为优势菌属,分别占分离放线菌菌株的63%和13.8%。对14株放线菌潜在新种(16S rRNA基因序列相似性低于98.65%)进行高氏一号、ISP2、TSB培养基液体发酵,获得提取浓缩物样品进行抑菌活性筛选,发现11株潜在新种的发酵产物有广谱抗菌活性。本研究表明,库姆塔格沙漠中含有丰富的放线菌资源,具有从中发现放线菌新菌种和抗生素的潜力。
关键词:库姆塔格沙漠;放线菌;16S rRNA基因测序;抑菌活性
中图分类号:R978文献标志码:A
Assessment of the diversity and antibacterial potential of culturable actinomycetes in the Kumtag Desert
Li Jiahui1, Wen Ying2,3,4, Tian Mao2,3,4, Liu Guangxiu2,3, Zhang Wei2,3, Zhang Gaosen2,3, and Xue Lingui1
(1 College of Biological and Pharmaceutical Engineering, Lanzhou Jiaotong University, Lanzhou 730000; 2 Key Laboratory of Desert and Desertification, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000; 3 Key Laboratory of Extreme Environmental Microbial Resources and Engineering, Lanzhou 730000; 4 University of Chinese Academy of Sciences, Beijing 100049)
Abstract Desert soil contains abundant actinomycetes, especially many rare strains, thus, it is an ideal habitat for excavating new actinomycete resources. In this study, the screening for antibacterial and culturable actinomycetes isolated from Kumtag Desert soil was carried out to provide strains for developing and utilizing actinomycete resources in this area. Actinomycete diversity was analyzed based on the 16S rRNA gene sequence of isolated strains; From 4 soil samples, 370 actinomycete strains, distributed in 22 genera of 12 families and 11 orders were isolated. Amongst them, Streptomyces and Saccharothrix were the dominant genera, accounting for 63% and 13.8% of the isolated actinomycete strains, respectively. Fourteen potential new actinomycete strains (16S rRNA gene sequence similarity less than 98.65%) were fermented in Gauze's synthetic, ISP2 and TSB broth media, respectively. Their fermentation broth was extracted for bacteriostatic activity screening, showing that fermentation products of 11 potential new species had broad-spectrum antibacterial activity. This study indicated that the Kumtag Desert was rich in actinomycete resources, for which potential new species of actinomycetes and antibiotics could be discovered.
Key wordsKumtag desert; Actinomycetes; 16S rRNA gene sequencing; Antibiotic activity
近年來,随着“超级细菌”的不断出现,耐药菌对人类健康造成了极大威胁。据美国FDA新药评价中心的数据,近30多年来新批准的抗生素数量逐年减少[1],寻求高效新型抗生素刻不容缓。据统计,临床上60%以上的抗生素来自放线菌[2-3]。目前,从传统栖息地分离到的放线菌产生的抗生素大多是重复分离,成本高且效率低。所以,越来越多的研究人员正在探索从极端环境或地球上未开发的栖息地分离出新的放线菌资源[4-9]。极端环境放线菌可长期在多重胁迫因子例如高温、低温、高酸、高碱或高盐等的极端环境中生长,使它们产生了独特的基因型和生理特性[10],具备产生新颖代谢产物的潜力[11]。
沙漠土壤蕴藏着丰富的稀有放线菌和放线菌新物种[12-15]。已从沙漠来源放线菌中分离得到多种具有新颖结构和生物活性的次级代谢产物。例如,在新疆沙漠中发现了一株诺卡菌属新种Nocardia XJ31,并从中分离得到了12个铁载体和2个蒽酮类化合物,其中10个铁载体被鉴定为新化合物,2个蒽酮类化合物表现出较好的抗结核活性[16];Liu等[17]從塔克拉玛干沙漠分离到一株糖丝菌属新种Saccharothrix 16Sb2-4,从中分离鉴定出了4种16元大环内酯类抗生素,这些化合物都通过抑制pDualrep2系统中的蛋白质合成而显示出抗菌活性。此外,Boubetra等[18]在撒哈拉沙漠中获得了一株糖丝菌属Saccharothrix SA198,其产生的2个新骨架化合物A4和A5,具有显著的抗细菌和真菌活性。因此,沙漠环境中的放线菌,具有巨大的潜在应用价值。
库姆塔格沙漠位于塔里木盆地东南的阿尔金山北麓,区域降水极为稀少,年均蒸发量2800~3000 mm;年平均温度10 ℃左右,是中国第四大流动沙漠,也是我国极为干旱、自然环境极为严酷的沙漠[19]。
目前,对该地区的细菌研究主要集中在可培养细菌多样性、抗辐射抗氧化的特征及环境对细菌群落结构的影响等[20-22],有关该地区放线菌多样性及其抑菌活性菌株筛选鲜有报道。根据扩增子研究结果发现该地区细菌丰富度较高,放线菌门是主要的细菌门[20-21]。
另外,可培养研究也支持放线菌为该地区最主要优势菌门的结论,其中链霉菌属(9.08×105 CFU/g)为优势菌属,占分离出的放线菌门可培养细菌总数量的
92% [22];柏晓玉等[20]对该地区细菌进行培养发现,链霉菌占所有可培养放线菌的65.76%。以上结果表明,库姆塔格沙漠地区蕴含着丰富的放线菌资源,是挖掘极端放线菌资源的理想生境。
本研究对库姆塔格沙漠放线菌进行分离培养、纯化和活性测定,通过16S rRNA基因测序对培养获得的菌株进行系统发育分析,并测定菌株的抑菌活性,以期为该地区放线菌资源开发和利用提供菌株资源。
1 材料与方法
1.1 材料
1.1.1 土样采集
2020年8月,采集库姆塔格沙漠区土壤样品4份,在对应地区进行编号(表1),每一样点采集200~300 g/袋,用无菌采样袋将所采集的样品暂时4 ℃
低温保存,然后在-20 ℃下进行保存,进行后续的可培养试验。
1.1.2 培养基
(1)分离培养基 选用12种不同的培养基进行培养, 配方参考德国菌种保藏中心(https://www.dsmz.de)的部分培养基。M1(高氏一号培养基G);M2(淀粉酪蛋白琼脂培养基SCA);M3(胰酪大豆胨琼脂培养基TSA);M4(0.1×TSA);M5(R2A);M6(PYGV);M7(甘油-天门冬酰胺培养基ISP5);M8(MM);M9(ISP3);M10(胰蛋白胨大豆肉汤培养基TSB);M11(0.1×TSB);M12(腐殖酸维生素琼脂培养基HVA)。
(2)纯化培养基 ISP4培养基配方参考德国菌种保藏中心(https://www.dsmz.de)。
(3)种子液培养基 0.1×TSA—胰蛋白胨1.7 g,
大豆蛋白胨0.3 g,NaCl 0.5 g,葡萄糖0.25 g,K2HPO4 0.05 g,H2O 1000 mL;R2A—R2A (BD,Difco) 3.2 g,H2O 1000 mL。
(4)发酵培养基 高氏一号、TSB和ISP2液体培养基配方参考德国菌种保藏中心,均不加琼脂。
(5)检定菌培养基 LB—胰蛋白胨10 g,酵母提取物5 g,NaCl 5 g,H2O 1000 mL,琼脂18 g。
1.1.3 主要仪器和试剂
生物安全柜(苏州智净净化有限公司);PCR扩增仪(吉泰生物有限公司);电泳仪(山东博晟生物技术有限公司);立式压力蒸汽灭菌锅(上海云泰仪器仪表有限公司);旋转蒸发仪(吉泰生物有限公司);电热恒温培养箱和恒温振荡培养箱(上海一恒科学仪器有限公司)。
Ex Taq DNA聚合酶,南京诺唯赞生物科技股份有限公司;Agarose,兰杰柯生物科技有限公司;其他试剂均为国产分析纯。
1.1.4 指示菌株
革兰阳性菌株:金黄色葡萄球菌(Staphylococcus aureus)、表皮葡萄球菌(Staphylococcus epidermidis)、蜡样芽胞杆菌(Bacillus cereus)、苏云金芽胞杆菌(Bacillus thuringiensis)、藤黄微球菌(Micrococcus luteus)、巴氏葡萄球菌(Staphylococcus pasteuri)。
革兰阴性菌株:大肠埃希菌(Escherichia coli)、肺炎克雷伯菌(Klebsiella pneunoniae)、铜绿假单胞菌(Pseudomonas aeruginosa),以上供试菌由甘肃省极端环境微生物资源与工程重点实验室提供。
1.2 方法
1.2.1 土壤样品的处理
在生物安全柜中,取5 g土壤样品置于50 mL装有5~10个玻璃珠且灭过菌的锥形瓶中,再将20 mL已灭菌生理盐水加入每个锥形瓶中,以200 r/min的速度在30℃恒温摇床上振荡40 min;振荡后取出静置
3 min,在900 ?L生理盐水中加入上清液100 ?L,混合均匀后标记为原液的10-1,再进行10倍梯度稀释至原液的10-5。在不同稀释浓度的土壤悬液中吸取
200 ?L,在上述12种分离培养基中进行涂布,37 ℃下培养7~14 d,培养1周后对平板进行计数和不同形态、颜色菌落的挑取、分离和纯化。
1.2.2 菌株的分离纯化
观察菌落形态,在同一分离平板内根据菌落形态特征去重后,挑取单菌落,编号,然后采用分区划线的方法接种在ISP4固体培养基上进行菌株纯化。
1.2.3 菌株的保藏
向冻存管(Corning,USA)中加入1.5 mL的25%的甘油水溶液,然后挑取已纯化的放线菌,混合均匀后保存至-80 ℃冰箱中。
1.2.4 基于16S rRNA基因序列的放线菌多样性分析
(1) 放线菌菌株基因组DNA提取 放线菌基因组DNA的提取按照试剂盒Solarbio?LIFE SCIENCES(北京)步骤进行。
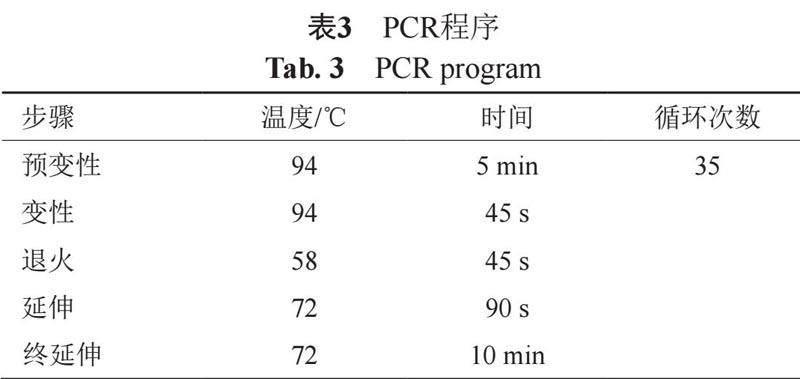
(2) 16S rRNA基因序列的PCR扩增 以提取的放线菌总DNA为模板,根据(表2)中的条件和(表3)中的程序来进行扩增。
PCR扩增产物在1%琼脂糖凝胶中进行电泳检测,PCR扩增产物送至西安擎科生物技术有限公司进行16S rRNA基因测序。
(3)序列比对及系统发育树的构建 将菌株的16S rRNA基因序列在EzBioCloud数据库进行比对(www.ezbiocloud.net);使用MEGA 11.0软件,采用邻接法进行聚类分析,构建系统发育树。系统进化矩阵采用Kimura two parameter模型进行评估,并重复取样1000次进行自展值分析,以评估进化树拓扑结构的稳定性。
1.2.5 放线菌抑菌活性检测
(1) 首先,在固体培养基上挑取适量生长良好的菌株,并将其转移到含有100 mL适合其生长的液体培养基的500 mL锥形瓶中。然后,放入摇床中,在180 r/min、30 ℃的条件下震荡培养2 d。接下来,加入高氏一号发酵培养基或TSB或ISP2,继续培养7 d。
完成发酵后,用8000 r/min的离心条件去除菌体,留下上清液,再用等量的乙酸乙酯萃取上清液。最后,使用旋转蒸发仪将提取物浓缩,再用2 mL甲醇溶解即可得到乙酸乙酯粗提物。
(2) 将实验室保存的指示菌接种到LB固体培养基中进行活化,接着,挑取活化好的指示菌加入到含有20 mL LB液体培养基的50 mL离心管中,在37 ℃、
180 r/min的摇床上震荡培养24~48 h,直至菌液生长繁殖。同时,配制LB固体培养基,并将其温度降至约55 ℃左右。随后,将0.1%的指示菌悬液加入到LB固体培养基中,并倾注于方形平板备用。
(3) 待上述方形平板的培养基凝固后,用5~6 mm的打孔器在方形平板上間隔1 cm打出小孔,吸取
20 μL乙酸乙酯粗提物注入小孔中,每个平板上留出一个孔加入20 μL浓度为 2 mg/mL的卡那霉素作为阳性对照,37 ℃培养24~48 h后观察并记录抑菌圈的有无和大小。
2 结果与分析
2.1 库姆塔格沙漠可培养放线菌数量特征
培养初期(2周后),对库姆塔格沙漠的两个样地(HLES和HLBS)放线菌进行了分离,计数不同培养基中分离到的菌落总数及放线菌数目(图1)所示。M8中分离到的细菌菌落数目最多,达1.12×104 CFU/g,M7分离到的菌落数最少(0.58×103 CFU/g)。从分离到的放线菌数来看,M1中分离到的放线菌数量最多(1.88×103 CFU/g),M12的最少(0.2×103 CFU/g)。综合菌落数目,M8、M1、M4 3种培养基的分离效果较好;而在M5培养基的平板上观察发现,菌落的种类相较于其他培养基较高。因此,选择这4种培养基来分离培养SSD和HMX两个样地的土壤放线菌。
2.2 库姆塔格沙漠可培养放线菌的多样性分析
从库姆塔格沙漠地区的土壤样品中分离纯化获得的纯培养物根据菌落形态、颜色、大小等特征进行初步重排,选择420株菌进行16S rRNA基因扩增和序列比对,结果表明,其中370株为放线菌,根据最新文献的修订[23],可以分为79个不同放线菌分类单元。它们隶属于11个目12个科的22个属,分离结果如(表4)所示,分别为:链霉菌属(Streptomyces)、糖丝菌属(Saccharothrix)、伦茨菌属(Lentzea)、假诺卡菌属(Pseudonocardia)、放线动孢菌属(Actinokineospora)、拟孢囊菌属(Kibdelosporangium)、拟无枝菌酸菌属(Amycolatopsis)、小单孢菌属(Micromonospora)、原小单孢菌属(Promicromonospora)、壤球菌属(Agrococcus)、节杆菌属(Arthrobacter)、戈登菌属(Gordonia)、拟诺卡菌属(Nocardiopsis)、贫养杆菌属(Modestobacter)、芽生球菌属(Blastococcus)、动球菌属(Kineococcus)、浮霉状菌属(Planctomonas)、微杆菌属(Microbacterium)、考克菌属(Kocuria)、污物假节杆菌属(Pseudarthrobacter)、Auraticoccus、类诺卡菌属(Nocardioides)。其中,链霉菌属为库姆塔格沙漠地区的优势菌属,共分离到233株,占分离到的放线菌菌株数量的63%,其次是糖丝菌属、伦茨菌属和节杆菌属,分别占13.8%、4.3%和3.8%。此外,其余菌属都不超过10株。部分代表菌株及其同源菌株基于16S rRNA基因序列构建的Neighbour-joining系统进化树见(图2)。
2.3 可培养放线菌潜在新种分析
根据放线菌的16S rRNA基因序列相似性低于98.65%可以被认为是不同物种的原则[9]。对分离到的可培养放线菌进行了潜在新颖性分析,结果显示,共有14株放线菌的16S rRNA基因序列与其最近缘的典型菌株序列相似性均低于98.65%,其中,链霉菌属7株,伦茨菌属、贫养杆菌属、污物假节杆菌属、类诺卡菌属、节杆菌属、动球菌属和糖丝菌属各1株,其相似性处于95.79%~98.64%之间。14株潜在放线菌新种中,6株是从HMX样地分离得到,4株是从SSD样地分离得到,4株是从HLES样地中分离得到,1株是从HLBS样地中分离得到,具体见表5。
2.4 抑菌活性筛选结果
对14株潜在放线菌新种分别进行了不同发酵培养基的摇瓶发酵,菌株发酵液乙酸乙酯萃取浓缩物对9种指示菌的抑菌活性统计结果显示(表6),不论是哪种发酵培养基,所有的检测菌株都至少对一种指示菌具有抑菌活性。部分菌株抑菌活性检测结果见(图3)。
其中,高氏一号培养基发酵时,SSD54、HMX87、HMX112、HMX145、HLES74、HLES54的发酵液乙酸乙酯萃取物对6种及以上指示菌有抑制作用,具有广谱抗菌活性,包括3株链霉菌和3株稀有放线菌(假诺卡菌属、节杆菌属和伦茨菌属)。HMX145、HMX169、HLES74的发酵液乙酸乙酯萃取物均对3种指示菌表现出强抑菌活性(抑菌圈直径>20.00 mm);而仅有HMX87对大肠埃希菌有良好的抑制作用,抑菌圈直径达17.00 mm。
ISP2培养基发酵时,SSD35、HMX87、HMX169、SSD95、HLES74、HLES153发酵液乙酸乙酯萃取物对6种及以上指示菌有抑制作用,同样具有广谱抗菌活性。但是,ISP2培养基发酵时,所有菌株的发酵液乙酸乙酯萃取物抑菌活性良好(抑菌圈直径<20.00 mm)。
TSB培养基发酵时,SSD35、HMX112、HMX169、HMX40、HLES54发酵液乙酸乙酯萃取物对6种及以上指示菌有抑制作用,具有广谱抗菌活性。同样,TSB培养基发酵时,只有HMX112对巴氏葡萄球菌和苏云金芽胞杆菌表现出显著的抑菌活性(抑菌圈直径>25.00 mm),并且其抑菌圈直径均大于阳性对照组;其他菌株的发酵液乙酸乙酯萃取物抑菌活性均良好(抑菌圈直径<20.00 mm)。
3 结论与讨论
来自库姆塔格沙漠区4份土壤样品中,分离到可培养的放线菌370株,分布于11个目12个科的22个属,除分离到链霉菌属外,还包括糖丝菌属、伦茨菌属、污物假节杆菌属、拟孢囊菌属、贫养杆菌属、芽生球菌属、浮霉状菌属、类诺卡菌属等21个稀有放线菌属,其中包括14株潜在放线菌新种。已有研究表明,从塔克拉玛干沙漠中分离到的放线菌分属于16科24属,链霉菌属和拟诺卡菌属为优势菌属,还分离到类诺卡菌属、库克菌属、糖单胞菌属、迪茨菌属等稀有放线菌[11]。另外,在西藏仲巴五彩沙漠中,优势菌仍然是链霉菌属,还有诺卡菌属、伦茨菌属、考克菌属、糖丝菌属和动球菌属等放线菌[9]。本研究从库姆塔格沙漠分离的放线菌中,链霉菌属也为优势菌屬,这可能与其能够产生抗干燥和耐热的孢子有关,孢子可以在干燥环境中长期存在[24]。而与文献不同的是本研究还分离到了拟孢囊菌属、贫养杆菌属、芽生球菌属、浮霉状菌属、污物假节杆菌属等一些稀有放线菌属。
70%的天然抗生素都是由放线菌产生的[25-27]。沙漠环境中的放线菌尤其是一些新物种已成为了新颖多样的次级代谢产物和新型抗生素的重要来源,例如,Abdelkader等[28]从分离自阿塔卡马沙漠土壤的链霉菌Streptomyces asenjonii KNN42.f中分离到3个具有广谱抗菌活性的新β-二酮类化合物;Bao等[29]从1株来自西藏仲巴五彩沙漠土壤的链霉菌XZH99T中分离到了7个angucycline类抗生素,其中3个为新化合物。活性检测表明,这些化合物对人肿瘤细胞具有显著的细胞毒性;从塔克拉玛干沙漠中分离出一株链霉菌8P21H-1,其表现出强大的抗菌活性和对蛋白质生物合成的抑制活性,通过分离得到乙酰灰绿蛋白和脱硫灰绿蛋白[30];此外,1株从阿根廷阿塔卡玛沙漠分离出的链霉菌新种Steptomyces leeuwenhoeki C34T,能够产生罕见的二十二元大环内酯类抗生素Chaxalactins A~C和新型安莎类抗生素chaxamycins A~D,这些新型抗生素具有良好的热稳定性和溶解性,并且表现出抗菌和抗肿瘤活性[31-32]。从沙漠放线菌中分离到具有良好的抗菌、抗肿瘤活性的物质,对生物医药领域具有积极影响。
本研究通过对14株潜在放线菌新种进行液体发酵及抑菌活性检测发现,它们至少对1种检定菌有抑菌活性,且对革兰阳性菌的抑制效果强于革兰阴性菌;其中,抑制苏云金芽胞杆菌的效果最为突出,抑菌活性最强的是利用TSB发酵的HMX112菌株,对巴氏葡萄球菌有显著的抑制效果,抑菌圈直径达30.30 mm,而阳性对照卡那霉素(2 mg/mL)抑制巴氏葡萄球菌的抑菌圈为21.30 mm,表明HMX112的抑菌活性极强;14株潜在放线菌新种中有11株对6种及以上指示菌有抑制作用,具有广谱的抑菌活性,其次级代谢产物值得下一步研究。此外,菌株HLES74、HMX112、HMX145、HMX169对革兰阳性菌(Staphylococcus aureus、Staphylococcus pasteuri、Bacillus thuringiensis、Staphylococcus epidermidis)较强的抑制效果,抑菌圈直径均大于20.00 mm,其中包括链霉菌、节杆菌、动球菌。本研究获得的菌株为筛选相应活性物质或挖掘新的生物活性物质提供了丰富的菌种资源,后续有待通过基因组信息分析、质谱分子网络技术[33]分析菌株次级代谢产物的产生能力,以进行进一步的次级代谢产物化学研究。
综上所述,库姆塔格沙漠环境中放线菌资源丰富并且一些潜在放线菌新种的抑菌活性较好,具有产生活性次级代谢产物的潜力。本研究为库姆塔格沙漠放线菌挖掘提供了经验,并获得了一批具有潜在应用价值的抑菌活性菌株,是后续挖掘沙漠放线菌高活性天然产物的基础。
参 考 文 献
叶景静. 广西茅尾海红树林植物根际土壤放线菌多样性及抗菌活性研究[D]. 桂林: 桂林医学院, 2023.
吕佩帅, 张丽红, 徐福洲, 等. 海洋放线菌资源、研究方法与生物活性研究进展[J]. 生物资源, 2021, 43(3): 232-245.
李菲, 楊玲, 王巧贞, 等. 红树林放线菌抗沃柑病原真菌的研究[J]. 中国抗生素杂志, 2022, 47(7): 638-646.
阚雨. 甘肃巴丹吉林沙漠细菌多样性及其酶活性和抗菌活性的研究[D]. 贵州: 贵州医科大学, 2021.
黄娇, 闫兵法, 黄英. 青藏高原阿里、那曲和海西地区土壤可培养放线菌的多样性[J]. 微生物学报, 2017, 57(9): 1342-1351.
Liu T, Wu S, Zhang R, et al. Diversity and antimicrobial potential of Actinobacteria isolated from diverse marine sponges along the Beibu Gulf of the South China Sea[J]. FEMS Microbiol Ecol, 2019, 95(7): fiz089.
Zhang D, Shu C Y, Lian X Y, et al. New antibacterial bagremycins F and G from the marine-derived Streptomyces sp. ZZ745 [J]. Mar Drugs, 2018, 16(330): 1-7.
Kamaruddin M, Marzuki I, Burhan A, et al. Screening acetylcholinesterase inhibitors from marine-derived actinomycetes by simple chromatography[J/OL]. IOP Conf Series: Earth Environm Sci, 2021, 679(1). https://doi.org/10.31219/osf.io/m2ku4.
李亚美, 布合力其汗·白克力, 鲍洁, 等. 西藏仲巴五彩沙漠放线菌资源勘探及生物活性筛选[J]. 微生物学通报, 2018, 45(8): 10.
Wen Y, Zhang G S, Bahadur A L, et al. Genomic investigation of desert Streptomyces huasconensis D23 reveals its environmental adaptability and antimicrobial activity[J]. Microorg, 2022, 10(12): 2408.
董艳萍, 郭琳, 旭格拉·哈布丁, 等. 塔克拉玛干沙漠南麓土壤放线菌资源勘探及抗菌活性筛选[J]. 中国抗生素杂志, 2013, 38(4): 241-247.
Sivakala K K, Gutiérrez-García K, Jose P A, et al. Desert environments facilitate unique evolution of biosynthetic potential in Streptomyces[J]. Molecules, 2021, 26: 588.
Cortes-Abayay C, Dorador C, Schumann P, et al. Streptomyces huasconensis sp. nov., an haloalkalitolerant actinobacterium isolated from a high-altitude saline wetland at the Chilean Altiplano[J]. Int J Syst Evol Microbiol, 2019, 69: 2315-2322.
Cortés-Albayay C, Jarmusch S A, Trusch F, et al. Downsizing class Ⅱ Lasso peptides: Genome mining-guided isolation of huascopeptin containing the first Gly1-Asp7 macrocycle[J]. Org Chem, 2020, 85: 1661-1667.
Wen Y, Zhang G S, Bahadur A L, et al. Streptomyces gobiensis sp. nov., an antimicrobial producing actinobacterium isolated from soil under black Gobi rocks[J]. Int J Syst Evol Micr, 2022, 72(3). doi: 10.1099/ijsem.0.005318.
Zhang L, Zhang J, Ren B, et al. Characterization of anti-BCG benz[α]anthraquinones and new siderophores from a Xinjiang desert-isolated rare actinomycete Nocardia sp. XJ31[J]. Appl Microbiol Biotechnol, 2020(12): 1-12.
Liu S, Wang T, Lu Q, et al. Bioprospecting of soil-derived actinobacteria along the Alar-Hotan Desert Highway in the Taklamakan desert[J]. Front Microbiol, 2021, 12: 604999.
Boubetra D, Sabaou N, Zitouni A, et al. Taxonomy and chemical characterization of new antibiotics produced by Saccharothrix SA198 isolated from a Saharan soil[J]. Microbiol Res, 2013, 168(4): 223-230.
梁爱民. 库姆塔格沙漠沉积物来源、搬运过程及贡献量化研究[D]. 北京: 中国科学院大学, 2020.
柏晓玉, 何子璇, 李雪, 等. 库木塔格沙漠土壤细菌多样性及具有群体感应抑制活性放线菌筛选[J].微生物学通报, 2023, 50(11): 4770-4783.
李娟, 刘阳, 刘光琇, 等. 鄯善库木塔格沙漠北缘细菌群落结构特征及影响因素[J]. 干旱区研究, 2023, 40(8): 1358-1368.
刘阳, 田茂, 王馨悦, 等. 可培养细菌多样性及抗辐射-抗氧化相关性特征——以库姆塔格沙漠东缘为例[J]. 中国环境科学, 2021, 41: 5921-5932.
Goker M. Filling the gaps: Missing taxon names at the ranks of class, order and family[J]. Int J Syst Evol Microbiol, 2022, 72: 5638.
Liang Y P, Zong Z F, Ma Q. Inhibiting and promoting effect on plants of six strains endophytic actinomycetes isolated from wild plants[J]. J Northwest A&F Univ, 2007, 35(7): 131-136.
李一青, 李艳琼, 李铭刚, 等. 稀有放线菌产生的抗生素[J]. 中国抗生素杂志, 2008, 33(4): 193-197.
Azman A S, Othman I, Fang C M, et al. Antibacterial, anticancer and neuroprotective activities of rare actinobacteria from mangrove forest soils[J]. Indian J Microbiol, 2017, 57(2): 177-187.
Bérdy J. Bioactive microbial metabolites[J]. J Antibiot, 2005, 58(1): 1-26.
Abdelkader, Mohamed S A, Philippon, et al. Asenjonamides A-C antibacterial metabolites isolated from Streptomyces asenjonii strain KNN 42.f from an extreme-hyper arid Atacama Desert soil[J]. J Antibiot, 2018, 71(4): 425-431.
Bao J, He F, Li Y, et al. Cytotoxic antibiotic angucyclines and actinomycins from the Streptomyces sp. XZHG99T[J]. J Antibiot, 2018, 71(12): 1018-1024.
Wang T, Li F, Lu Q, et al. Diversity, novelty, antimicrobial activity, and new antibiotics of cultivable endophytic actinobacteria isolated from psammophytes collected from Taklamakan Desert[J]. J Pharm Anal, 2021, 11(2): 241-250.
Rateb M E, Houssen W E, Harrison W T, et al. Diverse metabolic profiles of a Streptomyces strain isolated from a hyper-arid environment[J]. J Nat Prod, 2011, 74(9): 1965-1971.
Rateb M E, Houssen W E, Arnold M, et al. Chaxamycins A-D, bioactive ansamycins from a hyper-arid desert Streptomyces sp.[J]. J Nat Prod, 2011, 74(6): 1491-1499.
Quinn R A, Nothias L F, Vining O, et al. Molecular networking as a drug discovery, drug metabolism, and precision medicine strategy[J]. Trends Pharmacol Sci, 2017, 38(2): 143-154.
基金項目:第三次新疆综合科学考察项目(No. 2022xjkk1200);国家自然科学基金(No. U22A20451);中国科学院“西部之光”计划(No. xbzg-zdsys-202105);甘肃省科技计划项目(No. 23JRRA574)。
作者简介:李佳慧,女,生于1997年,硕士,研究方向:极端环境微生物学、微生物工程、应用微生物学,E-mail: 2051005547@qq.com
*通信作者,E-mail: xuelg@mail.lzjtu.cn
猜你喜欢
河南医学研究(2022年19期)2022-10-19
当代水产(2022年1期)2022-04-26
国际呼吸杂志(2019年22期)2019-12-09
生态学报(2019年11期)2019-07-08
少儿科学周刊·少年版(2015年3期)2015-07-07
少儿科学周刊·少年版(2015年3期)2015-07-07
应用海洋学学报(2014年4期)2014-11-22
天然产物研究与开发(2014年6期)2014-04-27
食品工业科技(2014年23期)2014-03-11
食品工业科技(2014年15期)2014-03-11
