
磷恢复对磷饥饿铜绿微囊藻光合色素和部分抗氧化酶活性的影响
2024-04-29 00:26贾滢暄张树林张达娟戴伟毕相东
中国农业科技导报 2024年1期
关键词:抗氧化酶
贾滢暄 张树林 张达娟 戴伟 毕相东
摘要:为探究磷饥饿及磷恢复对铜绿微囊藻光合色素、藻胆蛋白和抗氧化酶等生理指标的影响,将其进行磷饥饿处理7 d后再进行磷恢复,检测磷恢复前后铜绿微囊藻的藻细胞密度、叶绿素a、类胡萝卜素、藻蓝蛋白(phycocyanin,PC)、别藻蓝蛋白(allophycocyanin,APC)、藻红蛋白(phycoerythrin,PE)、丙二醛(malondialdehyde,MDA)和过氧化氢(H2O2)含量及超氧化物歧化酶(superoxide dismutase,SOD)活性的变化。结果表明,铜绿微囊藻磷饥饿处理7 d后,藻细胞密度为2.54×107 cell·mL-1,显著低于对照组(3.11×107 cell·mL-1)。磷恢复144 h后,处理组藻细胞密度为4.05×107 cell·mL-1,仍显著低于对照组(4.32×107 cell·mL-1);叶绿素a、类胡萝卜素和PC、APC、PE含量均呈现升高趋势,在144 h时分别达到5.96、1.44 μg·mL-1和0.031、0.02、0.065 mg·L-1;MDA、H2O2含量和SOD活性呈先上升后下降趋势,均在48 h达到最大值,较对照组分别增加36.2%、47.7%、51.1%。由此表明,磷恢复后铜绿微囊藻的藻细胞密度、叶绿素a、类胡萝卜素和藻胆蛋白含量虽呈升高趋势,但难以恢复到对照水平;MDA、H2O2含量及SOD活性的变化也说明,从磷饥饿到磷恢复后铜绿微囊藻藻细胞受到氧化损伤,并对细胞膜系统产生破坏。
关键词:磷恢复;磷饥饿;铜绿微囊藻;光合色素;藻胆蛋白;抗氧化酶
doi:10.13304/j.nykjdb.2022.0561
中图分类号:S946.3 文献标志码:A 文章编号:10080864(2024)01007008
随着我国经济发展和生产工艺的增多,大量氮、磷等营养物质排入水体。水体富营养化问题日趋严重[12],常导致蓝藻门种群大量繁殖,成为优势种群,进而形成水华[34]。蓝藻水华不仅使水生态环境恶化,还易产生毒素,通过生物积累影响人类健康。微囊藻(Microcystis)作为重要的蓝藻水华种群之一,受到广泛关注[5]。影响微囊藻生长的因素较多,包括浮游动物捕食等生物因素以及营养盐、光照、温度等非生物因素[6]。长期以来,普遍认为过多的营养元素会促使微囊藻爆发性增长[7],水体中氮、磷浓度与微囊藻的生长关系密切,且磷对铜绿微囊藻生长有较大的限制作用[8]。
磷是藻类重要的营养元素,既是核酸、蛋白质和磷脂的主要成分,又是合成叶绿素的必需物质[9]。磷在水体中的形态主要以溶解态磷和颗粒态磷为主,微生物可直接利用溶解性无机磷[10-11]。微囊藻对磷盐的利用主要通过自身核酸、叶绿素和磷脂等物质的合成来完成。当水体磷水平较高时,微囊藻细胞能吸收大量的含磷营养物质,并在细胞内以不同形式储存,用来支持磷缺乏时藻细胞的正常生长和分裂[5]。因此磷元素被认为是影响微囊藻爆发性增长的重要限制性因素[12]。许海等[13]研究发现,不同形态的磷均能被铜绿微囊藻(Microcystis aeruginosa)吸收利用,且铜绿微囊藻在低磷水平下的生长速率高于高磷水平。岳冬梅等[14]发现,经过氮、磷饥饿的铜绿微囊藻在补充营养盐后都能快速生长,然而,经氮饥饿处理的铜绿微囊藻在氮恢复后虽然可快速生长但容易出现衰亡;而磷饥饿处理的藻细胞在磷恢复后均以较稳定的速率增长,且未出现衰亡,藻细胞的碳固定能力也可以恢复到最初水平。目前研究者大多关注不同氮磷的质量比或不同氮磷营养盐来源、种类对铜绿微囊藻的影响,及在营养盐恢复后微囊藻的生长情况,而有关生理指标及氧化损伤等方面的研究较少。
因此本研究以铜绿微囊藻905藻株为研究对象,分析磷饥饿后恢复供磷对其生理指标(色素、藻胆蛋白及抗氧化酶等)的影响,为探讨磷营养盐在铜绿微囊藻生长中的作用以及预防和治理微囊藻水华提供参考。
1 材料与方法
1.1 试验藻种
铜绿微囊藻905藻株购自中国武汉科学院水生生物研究所。使用BG11 培养基在光照强度40 μmol·m-2·s-1、(27±2) ℃、光照12 h/黑暗12 h的条件下进行培养。
1.2 试验设计
试验设置1个对照组和1个处理组,二者分别使用标准BG11培养基和不添加磷营养盐的BG11培养基,每组分别设置3个平行。铜绿微囊藻初始密度均为2×107 cell·mL-1,进行不同水平磷处理7 d后,将对照组和处理组中的铜绿微囊藻离心,取藻泥,分别接种至标准的BG11培养基中。在磷恢复后的0、12、24、48、72、96、120、144 h取样,测定各项生理指标。
1.3 生理指标的测定
1.3.1 藻细胞密度测定 用血球计数板对铜绿微囊藻进行细胞计数,每个平行样品均重复计数3次,取平均值。
1.3.2 叶绿素a和类胡萝卜素含量测定 参考戴立洲等[15]的方法测定叶绿素a(chlorophyll a,Chla)和类胡萝卜素(carotenoid,Car)含量(μg·mL-1)。具体操作步骤如下:取10 mL 藻液,以4 000 r·min-1 离心10 min,得到藻泥沉淀,加入10 mL的95%(体积分数)乙醇,摇晃使乙醇与藻泥混合均匀,在4 ℃下进行8 h的提取,提取结束后再次离心取上清液,分别在665、649和470 nm波长下测定其吸光光度值,计算叶绿素a和类胡萝卜素含量。
1.3.3 藻胆蛋白含量测定 参考戴立洲等[15]的方法并作修改。取5 mL藻液,在4 000 r·min-1条件下离心10 min,得到藻泥沉淀,向离心管中加入5 mL磷酸缓冲液(0.05 mol·L-1,pH 7.0)充分混匀,包锡纸,放入?80 ℃的超低温冰箱冷冻8 h 以上;然后取出于室温下避光溶解,反复冻融3次,再次离心取上清液,分别测定其在650、620和565 nm的吸光值并记录。计算藻蓝蛋白(phycocyanin,PC)、别藻蓝蛋白(allophycocyanin,APC)和藻红蛋白(phycoerythrin,PE)含量(mg·L-1)。
1.3.4 丙二醛、超氧化物歧化酶和过氧化氢的测定 取2 mL 藻液以4 000 r·min-1 离心10 min,弃上清液,藻泥加入200 μL生理盐水混匀,使用超微粉碎机进行破碎处理,得到破碎液待用。从破碎液中取50 μL,加入200 μL 的生理盐水,以2 500 r·min-1离心10 min,取上清液用于测定蛋白含量;剩余的破碎液以4 000 r·min-1离心10 min,取250 μL上清液以10 000 r·min-1离心10 min,用于测定过氧化氢(H2O2)含量;剩余上清液用于丙二醛(malondialdehyde,MDA)含量和超氧化物歧化酶(superoxide dismutase,SOD)活性的测定。以上指标测定均采用南京建成生物工程研究所试剂盒测定,具体方法参考试剂盒说明书。
1.4 统计分析
采用Excel 2010 和SPSS 24.0 进行数据整理与统计分析。
2 结果与分析
2.1 磷恢复对磷饥饿铜绿微囊藻生长的影响
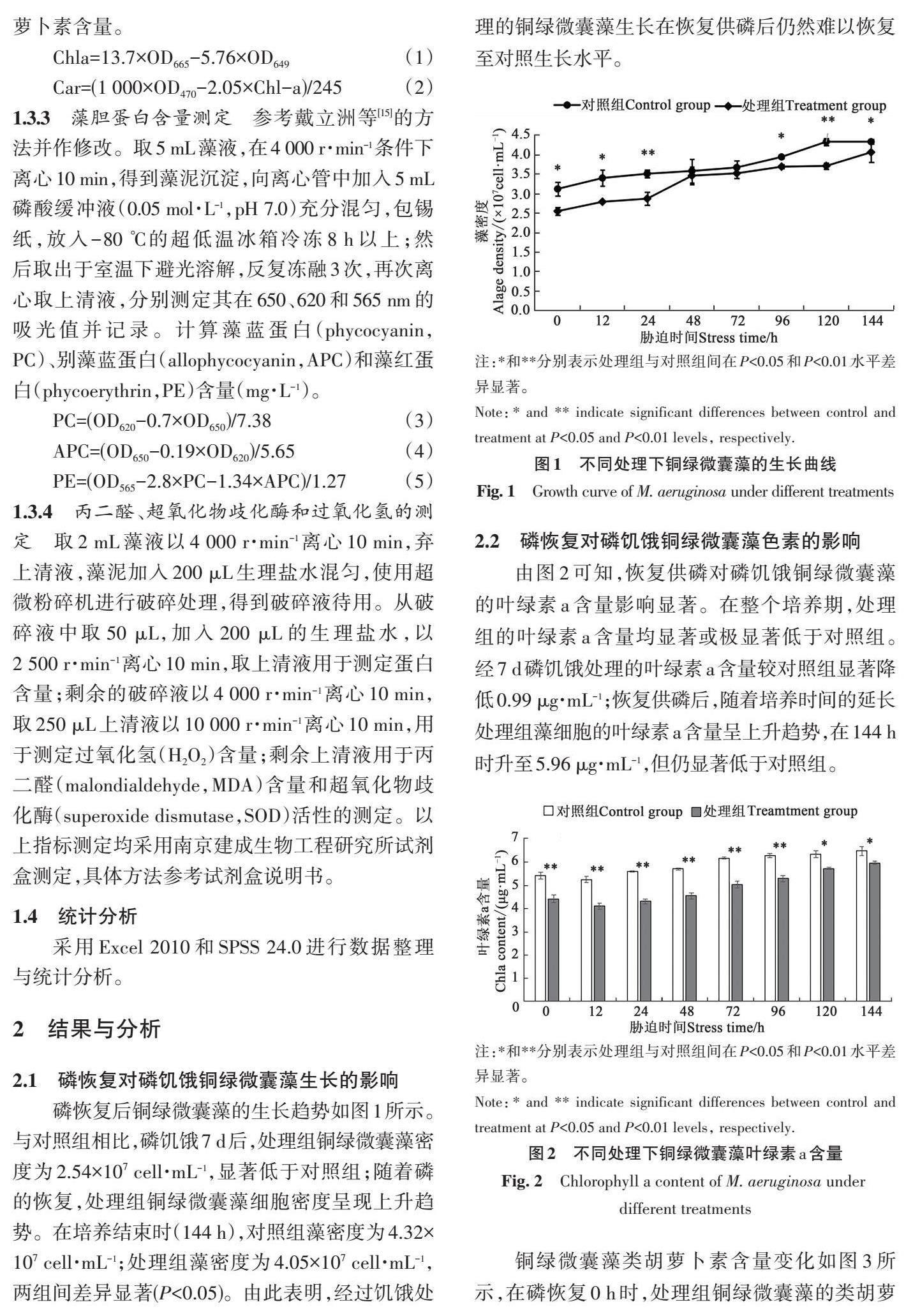
磷恢复后铜绿微囊藻的生长趋势如图1所示。与对照组相比,磷饥饿7 d后,处理组铜绿微囊藻密度为2.54×107 cell·mL-1,显著低于对照组;随着磷的恢复,处理组铜绿微囊藻细胞密度呈现上升趋势。在培养结束时(144 h),对照组藻密度为4.32×107 cell·mL-1;处理组藻密度为4.05×107 cell·mL-1,两组间差异显著(P<0.05)。由此表明,经过饥饿处理的铜绿微囊藻生长在恢复供磷后仍然难以恢复至对照生长水平。
2.2 磷恢复对磷饥饿铜绿微囊藻色素的影响
由图2可知,恢复供磷对磷饥饿铜绿微囊藻的叶绿素a含量影响显著。在整个培养期,处理组的叶绿素a含量均显著或极显著低于对照组。经7 d磷饥饿处理的叶绿素a含量较对照组显著降低0.99 μg·mL-1;恢复供磷后,随着培养时间的延长处理组藻细胞的叶绿素a含量呈上升趋势,在144 h时升至5.96 μg·mL-1,但仍显著低于对照组。
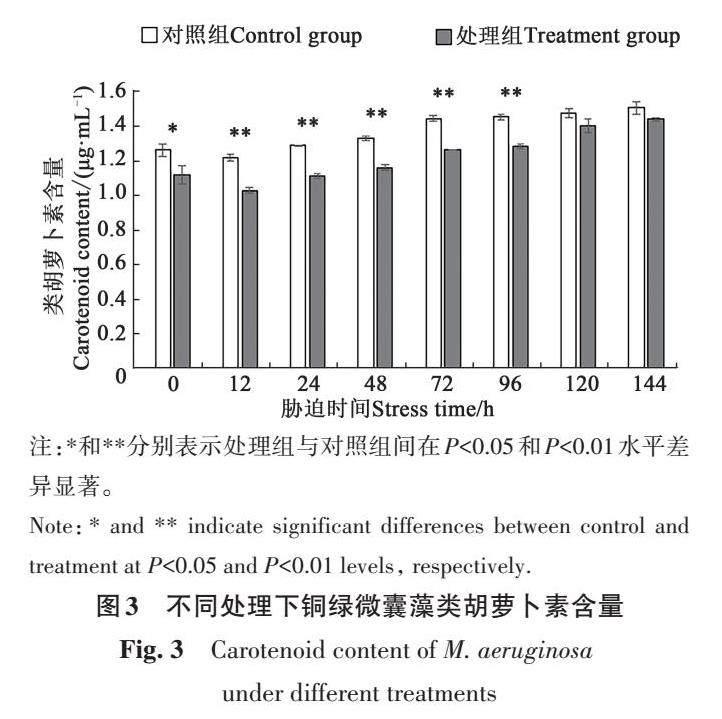
铜绿微囊藻类胡萝卜素含量变化如图3 所示,在磷恢复0 h时,处理组铜绿微囊藻的类胡萝卜素含量为1.12 μg·mL-1,显著低于对照组(0.14 μg·mL-1);在磷恢复144 h时,处理组藻细胞的类胡萝卜素含量升至1.44 μg·mL-1,与对照组差异不显著。
2.3 磷恢复对磷饥饿铜绿微囊藻藻胆蛋白含量的影响
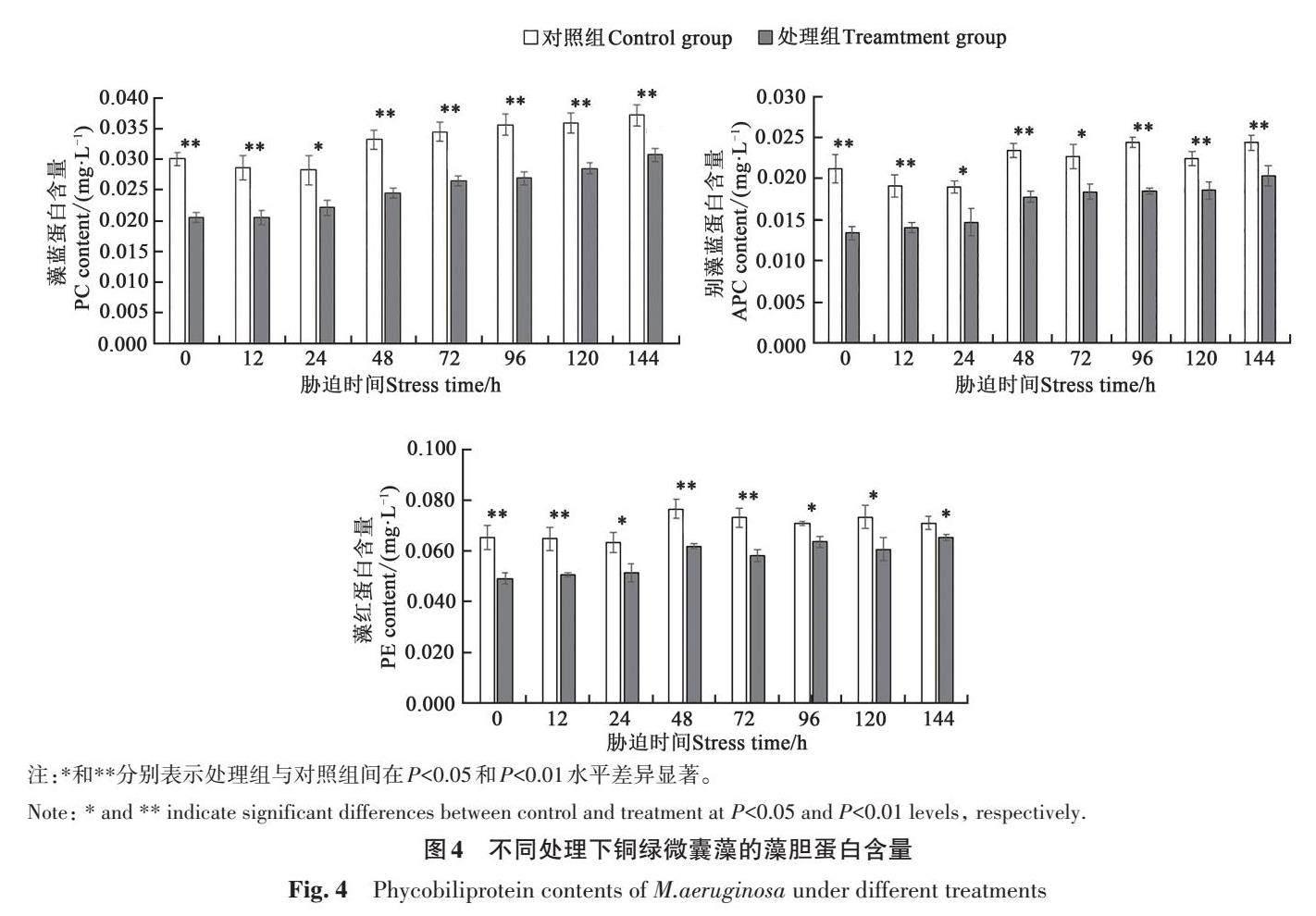
由图4可知,在磷恢复144 h内,处理组铜绿微囊藻的藻蓝蛋白(PC)含量极显著低于对照组。经7 d磷饥饿处理后,处理组PC含量极显著低于对照组;恢复供磷后,随着培养时间的延长,处理组PC含量逐渐上升至0.031 mg·L-1,但仍极显著低于对照组。经7 d磷饥饿处理后,处理组藻细胞的别藻蓝蛋白(APC)含量为0.013 mg·L-1,极显著低于对照组;随着培养时间的延长,APC含量呈上升趋势,在144 h时升至0.020 mg·L-1,但仍极显著低于对照组。经7 d磷饥饿处理后,处理组藻细胞的藻红蛋白(PE)含量极显著低于对照组;在恢复供磷144 h时,对照组与处理组的PE含量分别为0.071和0.065 mg·L-1,二者差异显著。
2.4 磷恢复对磷饥饿铜绿微囊藻MDA 及H2O2含量的影响
由图5可知,对照组铜绿微囊藻细胞MDA含量在培养期相对稳定;经7 d磷饥饿处理后的处理组MDA含量为1.63 nmol·mg-1,与对照组差异不显著,随后MDA含量呈现上升趋势,在48 h时升至2.52 nmol·mg-1,较对照组极显著增加36.2%。在磷恢复48~144 h,处理组MDA含量逐渐下降并趋于稳定;在144 h时处理组与对照组间无显著差异。在恢复供磷0~48 h,处理组藻细胞的H2O2含量逐渐升高,并在48 h时升至10.41 mmol·g-1,较对照组极显著增加47.7%;随后H2O2含量逐渐下降,并在120~144 h趋于稳定,在144 h时处理组H2O2含量为5.40 mmol·g-1,与对照组差异不显著(图5)。
2.5 磷恢复对磷饥饿铜绿微囊藻SOD 活性的影响
磷恢复对磷饥饿藻细胞SOD 活性有显著影响(图6)。恢复供磷后,处理组藻细胞SOD活性显著升高,并在48 h时升至10.62 U·mg-1,较对照组极显著提高51.1%;48 h后,SOD活性逐渐降低并趋于稳定,在144 h时,处理组藻细胞SOD活性为7.70 U·mg-1,较对照组极显著高20.9%。
3 讨论
前人对环境因素对微囊藻生长的影响进行了大量研究[16]。磷是微藻细胞组成和合成ATP的重要元素,参与细胞膜构建、能量传递和信号传导等代谢活动。谢静等[17]研究发现,氮、磷元素均对铜绿微囊藻生长有显著影响,但高磷水平对其生长的影响更为显著。微囊藻的胞内磷主要以有机磷形式存在,因此,其生长速度也取决于胞内磷含量[18],微囊藻对磷的最大摄取速率较高,具有比其他藻类更强的储磷能力。磷限制会影响藻类的生长繁殖、光合作用、产毒及胞内物质合成等[19],当外界环境磷缺乏时,微藻可以利用自身储存的磷营养盐继续生长[20-21]。
藻类依靠光合作用将有机物固定在体内,因此光合作用是藻类生长过程中最重要的生理过程之一[22],叶绿素a与类胡萝卜素在光合作用中起着重要作用。田雅琪[23]研究发现,不同氮、磷水平及氮磷比对微囊藻生长的影响相似,均表现为低水平抑制,高水平促进;随着氮、磷水平的升高,β-胡萝卜素含量呈先增加后下降趋势,叶绿素a呈直线上升趋势。叶倩[24]研究发现,缺磷时铜绿微囊藻的藻细胞中叶绿素a和类胡萝卜素含量都呈现显著下降趋势,其生长受到抑制,并在培养后期逐渐衰亡。李晓梅等[25]指出,磷限制会使三角褐指藻(Phaeodactylum tricornutum Bohlin)叶绿素a含量降低。在缺磷环境中,藻类的细胞分裂受阻,且对光能的吸收和利用能力降低,营养物质可能会更多地流向其他代谢过程,而对光合色素合成上的投入减少。本研究中磷饥饿会抑制铜绿微囊藻的生长,且在磷恢复后处理组铜绿微囊藻的生长速率仍难以恢复;磷饥饿导致藻细胞叶绿素a和类胡萝卜素含量显著降低,随着磷恢复后培养时间的延长,二者含量逐渐升高,但在培养末期仍未恢复至对照水平,可见磷饥饿对铜绿微囊藻的生长及叶绿素a和类胡萝卜素含量有明显的抑制作用。
藻胆蛋白主要存在于蓝藻、红藻、隐藻和少数甲藻中,是重要的捕光色素蛋白,在一些藻类中也可作为储藏蛋白[26]。郑彩云等[27]研究表明,随着氮水平的上升,紫球藻(Porphyridium cruentum)的藻胆蛋白含量明显增多,在一定范围内其含量与氮含量呈正相关。刘洁等[28]指出,低水平Ni增加铜绿微囊藻PC含量,而高水平Ni则降低其含量;但不同Ni水平均使APC含量降低。本研究表明,磷恢复后铜绿微囊藻的藻胆蛋白含量均呈上升趋势,且与叶绿素a 含量和藻细胞生长呈正相关。由此可见,磷恢复后,随着藻细胞叶绿素a含量升高,光合作用能力逐渐提高,对培养基中磷源的利用能力增强,合成藻胆蛋白含量增加[27]。
抗氧化酶系统是生物体减轻细胞遭受氧化损伤的重要防御系统,通过清除细胞中的活性氧(reactive oxide species,ROS)提高细胞对氧化压力耐受性,是生物的一种保护机制。超氧化物歧化酶(SOD)是生物体清除ROS的关键酶,能催化O-2发生歧化反应,生成H2O2,后者再被其他抗氧化酶去氧化生成水分子,是藻体防御ROS的第一道防线。MDA是膜脂过氧化的产物,能够反映膜氧化损伤程度[2930]。H2O2是光合电子传递链的天然产物,对藻类具有毒害作用[31]。Li等[32]探究了磷限制对小球藻(Chlorella)的影响,结果表明随着磷水平的增加藻细胞MDA含量和SOD活性均逐渐降低。王小冬等[33]发现,磷饥饿抑制了赤潮异弯藻(Heterosigma akashiwo)和海洋卡盾藻(Chattonella marina)生长,藻细胞SOD 活性提高以缓解胁迫损伤。马金华等[31]研究发现,链状亚历山大藻(Alexandrium catenella)在衰亡期及高、低氮条件下,藻细胞内H2O2 含量明显上升,说明胁迫条件下藻细胞会产生过量的O-2,致使H2O2积累。杨蕾[34]研究发现,在不同盐胁迫下发状念珠藻(Nostoc flaglliforme)藻细胞的O-2 和H2O2含量随胁迫时间的延长而升高,表明盐胁迫对其膜系统造成破坏。本研究发现,磷恢复后铜绿微囊藻藻细胞的MDA、H2O2含量及SOD活性均呈现先上升后下降的趋势,表明磷饥饿处理使细胞生长受到抑制,造成蛋白质降解,产生大量的ROS,使得H2O2 和MDA 含量上升;细胞的抗氧化系统被激活,SOD等抗氧化酶活性上升,以缓解ROS等有害物质对机体造成的损伤[31]。
参考文献
[1] 张维清,韩丽华,林立清.太湖微囊藻水华暴发现状及其研究进展[J].江西科学,2018,36(5):830-832,876.
ZHANG W Q, HAN L H, LIN L Q. Current situation ofMicrocystis blooms outbreak in Lake Taihu and its researchprogress [J]. Jiangxi Sci., 2018, 36(5):830-832,876.
[2] 田慧捷,韩帮忠.水库中微囊藻毒素监测技术研究[J].中国资源综合利用,2021,39(2):122-124.
TIAN H J, HAN B Z. Study on the monitoring technology ofMicrocystin in the reservoir [J]. China Resour. ComprehensiveUtilization, 2021, 39(2):122-124.
[3] 邢彤. 巢湖蓝藻水华动态变化特征及其控制技术评估研究[D].合肥:合肥学院,2020.
XING T. Dynamic characteristics of cyanobacteria bloom inChaohu Lake and evaluation of its control techniques [D].Hefei: Hefei College, 2020.
[4] 鲁男.环境因子对铜绿微囊藻生长及产毒的影响研究[D].沈阳:辽宁大学,2015.
LU N. Effect of environmental factors on Microcysis and itstoxin production [D]. Shenyang: Liaoning University, 2015.
[5] 许海,吴雅丽,杨桂军,等.铜绿微囊藻、斜生栅藻对氮磷饥饿的耐受能力研究[J].生态科学,2014,33(5):879-884.
XU H, WU Y L, YANG G J, et al .. Tolerance of Microcystisaeruginosa and Scendesmus obliquus to nitrogen andphosphorus deficiency [J]. Ecol. Sci., 2014, 33(5):879-884.
[6] 许慧萍,杨桂军,周健,等. 氮、磷浓度对太湖水华微囊藻(Microcystis flos-aquae)群体生长的影响[J]. 湖泊科学,2014,26(2):213-220.
XU H P, YANG G J, ZHOU J, et al .. Effect of nitrogen andphosphorus concentration on colony growth of Microcystis flosaquaein Lake Taihu [J]. Lake Sci., 2014, 26(2):213-220.
[7] 袁若愚.铜绿微囊藻低磷条件下爆发生长及其化感防治的机理研究[D].青岛:青岛大学,2020.
YUAN R Y. The mechanism of low dissolved phosphorus and allelopathic inhibition on growth of microcystis aeruginosa [D].Qingdao: Qingdao University, 2020.
[8] 郑晓宇,金妍,任翔宇,等.不同氮磷浓度对铜绿微囊藻生长特性的影响[J].华东师范大学学报(自然科学版),2012(1):11-18.
ZHENG X Y, JIN Y, REN X Y, et al .. Influence of nitrogen andphosphorus concentrations on the growth characteristics ofMicrocystis aeruginosa [J]. J. East China Normal Univ. (Nat.Sci.), 2012(1):11-18.
[9] 严杨蔚,代瑞华,刘燕,等.氮和磷对有害藻类生长及产毒影响的研究进展[J].环境与健康杂志,2013,30(4):358-362.
YAN Y W, DAI R H, LIU Y, et al .. Progress of research oneffects of nitrogen and phosphorus on growth and toxinproduction of algae: a review of recent studies [J]. Environ.Health, 2013, 30(4):358-362.
[10] 叶倩.铜绿微囊藻缺氮缺磷下的生理生化响应[D].厦门:厦门大学,2019.
YE Q. Physiological and biochemical responses of microcystisaeruginosa to nitrogen and phosphorus deficiency [D]. Xiamen:Xiamen University, 2019.
[11] XIE E, SU Y P, DENG S Q, et al .. Significant influence ofphosphorus resources on the growth and alkaline phosphataseactivities of Microcystis aeruginosa [J/OL]. Environ. Pollution, 2021,268: 115807 [2022-06-05]. https://doi.org/10.1016/j.envpol.2020.115807.
[12] HEE Y Y, SURATMAN S, TAHIR N M, et al .. Seasonalvariability and fractionation of P-based nutrients in SungaiSetiu Basin, Terengganu, Malaysia [J]. Sains Malays., 2018, 47:883-891.
[13] 许海,陈丹,陈洁,等.氮磷形态与浓度对铜绿微囊藻和斜生栅藻生长的影响[J].中国环境科学,2019,39(6):2560-2567.
XU H, CHEN D, CHEN J, et al .. Effects of nitrogen andphosphorus forms and concentrations on the growth ofMicrocystis aeruginosa and Scenedesmus obliquus [J]. ChinaEnviron. Sci., 2019, 39(6):2560-2567.
[14] 岳冬梅,李洁,肖琳.营养盐恢复对氮磷饥饿铜绿微囊藻生长的影响[J].环境科学,2016,37(11):4220-4227.
YUE D M, LI J, XIAO L. Nutrients recovery on the growth ofnitrogen and phosphorus starved Microcystis aeruginosa [J].Environ. Sci., 2016, 37(11):4220-4227.
[15] 戴立洲,成小英,俞珊,等.豆瓣菜有机提取物对铜绿微囊藻的抑制及成分初步分离[J].环境科学学报,2015,35(12):4159-4168.
DAI L Z, CHENG X Y, YU S, et al .. Inhibitory effects of organicsolvent extract from nasturtium officinale on Microcystisaeruginosa associated with isolation of the allelochemicalingredients [J]. Acta Sci. Circumstantiae, 2015, 35(12): 4159-4168.
[16] 陈国永,杨振波,马昱,等.氮和磷对铜绿微囊藻细胞生长的影响[J].环境与健康杂志,2007(9):675-679.
CHEN G Y, YANG Z B, MA Y, et al .. Effects of nitrogen andphosphorus on growth of Microcystis aeruginosa strains [J].Environ. Health., 2007(9):675-679.
[17] 谢静,程燕,查燕,等.氮磷营养盐对铜绿微囊藻和斜生栅藻生长及竞争的影响[J].江西农业大学学报,2021,43(03):694-702.
XIE J, CHENG Y, CHA Y, et al .. Effects of nitrogen andphosphorus on the growth and competition of Mcirocystisaeruginosa and Scenedesmus obliquus [J]. Acta Agric. Univ.Jiangxiensis, 2021, 43(3):694-702.
[18] 许海,吴雅丽,杨桂军,等.铜绿微囊藻、斜生栅藻对氮磷饥饿的耐受能力研究[J].生态科学,2014,33(5):879-884.
XU H, WU Y L, YANG G J, et al.. Tolerance of Microcystisaeruginosa and Scendesmus obliquus to nitrogen and phosphorusdeficiency [J]. Ecol. Sci., 2014, 33(5):879-884.
[19] 张文珺. 无机磷限制对圆海链藻的生长及转录表达的影响[D].舟山:浙江海洋大学,2021.
ZHANG W J. Effects of inorganic phosphorus limitation on thegrowth and transcription expression of Thalassiosira rotula [D].Zhoushan: Zhejiang Ocean University, 2021.
[20] 秦梦瑶.铜绿微囊藻有机磷酶解利用及影响因素研究[D].南京:南京师范大学,2021.
QIN M Y. Study on enzymatic hydrolysis and utilization oforganic phosphorus in Microcystis aeruginosa and its influencingfactors [D]. Nanjing: Nanjing Normal University, 2021.
[21] 胡春艳.普通小球藻和波吉卵囊藻的生长特性及其对氮磷利用规律研究[D].荆州:长江大学,2021.
HU C Y. Study on the growth characteristics and nitrogen andphosphorus utilization of Chlorella vulgaris and Oothecabojiensis [D]. Jinzhou: Changjiang University, 2021.
[22] 侯新星,田如男.有机酸对铜绿微囊藻生长及光合色素的影响[J].生物学杂志,2021,38(4):65-70.
HOU X X, TIAN R N. Effects of organic acids on growth andphotosynthetic pigment of Microcystis aeruginosa [J]. J. Biol.,2021, 38(4):65-70.
[23] 田雅琦.环境因素对微囊藻色素及脂肪酸组成的影响规律研究[D].杨凌:西北农林科技大学,2019.
TIAN Y Q. Effects of environmental factors on pigment andfatty acid composition of Microcystis aeruginosa [D].Yangling:Northwest A&F University, 2019.
[24] 叶倩.铜绿微囊藻缺氮缺磷下的生理生化响应[D].厦门:厦门大学,2019.
YE Q. Physiological and biochemical responses of Microcystisaeruginosa to nitrogen and phosphorus deficiency [D]. Xiamen:Xiamen University, 2019.
[25] 李小梅,夏建荣.氮磷营养限制影响三角褐指藻光合无机碳利用和碳酸酐酶活性[J].水生生物学报,2013,37(3):405-412.
LI X M, XIA J R. Effects of nitrogen or phosphorus limitationon photosynthetic inorganic carbon utilization and carbonicanhydrase activity in Phaeodactylum tricornutum [J]. Acta Hydrobiol. Sin., 2013, 37(3):405-412.
[26] 徐宏洲,杨宸,彭俊,等.氟苯尼考对铜绿微囊藻生长和生理特征的影响[J].上海海洋大学学报,2021,30(1):120-128.
XU H Z, YANG C, PENG J, et al .. Effects of florfenicol on thegrowth and physiology of cyanobacteria (Microcystis aeruginosa) [J].J. Shanghai Ocean Univ., 2021, 30(1):120-128.
[27] 郑彩云,卢伟婷,王艳,等.不同氮浓度对紫球藻生长及藻胆蛋白和叶绿素a 含量变化的影响[J]. 中国酿造,2013,32(6):133-135.
ZHENG C Y, LU W T, WANG Y, et al .. Effect of differentNaNO3 concentration on Porphyridium growth and the contentchanges of phycobiliproteins and chlorophyll a [J]. ChinaBrew., 2013, 32(6):133-135.
[28] 刘洁,常学秀,黄丽娟,等.Ni元素对铜绿微囊藻的生长、光谱特性及藻胆蛋白含量的影响[J].云南大学学报(自然科学版),2005(4):365-368.
LIU J, CHANG X X, HUANG L J, et al .. Effects of Ni on thegrowth, absorption spectrum and phycobiliprotein content ofMicrocystis aeruginosa [J]. J. Yunnan Univ. (Nat. Sci), 2005(4):365-368.
[29] 付梅,宋秀贤,俞志明,等.伪矮海链藻抗氧化酶活性对磷化氢的响应特征[J].海洋环境科学,2013,32(6):809-813.
FU M, SONG X X, YU Z M, et al .. Responses of activity ofantioxidant enzyme in Thalassiosira pseudonana to phosphine [J].Marine Environ. Sci., 2013, 32(6):809-813.
[30] 郑金秀,彭祺,张甲耀,等.藻类产生及清除过氧化氢的研究[J].微生物学杂志,2006(6):80-84.
ZHENG J X, PENG Q, ZHANG J Y, et al .. Advance inH2O2 producing and scavenging induced by algae [J]. J.Microbiol., 2006(6):80-84.
[31] 马金华,孟希,张淑,等.链状亚历山大藻赤潮衰亡的生理调控[J].生态学报,2013,33(13):3978-3986.
MA J H, MENG X, ZHANG S, et al .. Physiological regulationrelated to the decline of alexandrium catenella [J]. Acta Ecol.Sin., 2013, 33(13):3978-3986.
[32] LI X, PAN J, LU Z, et al .. Arsenate toxicity to the marinemicroalga Chlorella vulgaris increases under phosphoruslimitedcondition [J]. Environ. Sci. Pollut. Res. 2021, 28(36):50908-50918.
[33] 王小冬,王艳.赤潮异弯藻和海洋卡盾藻抗氧化酶活性对氮磷比失衡的响应[J]. 海洋环境科学,2012,31(3):4.
WANG X D, WANG Y. Response of antioxidant enzymeactivities in Heterosigma akashiwo and Chattonella marina tolose-balance of N/P ratio [J]. Marine Environ. Sci., 2012, 31(3):4-16.
[34] 杨蕾.盐胁迫下发状念珠藻抗氧化系统及蛋白质、脂质的研究[D].济南:齐鲁工业大学,2013.
YANG L. Effects of salt stress on the antioxidation system,proteins and lipids of cyanobacteria Nostoc flagelliforme [D].Jinan: Qilu University of Technology, 2013.
(责任编辑:张冬玲)
猜你喜欢
中国医药导报(2016年33期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
安徽农学通报(2016年24期)2017-01-12
现代园艺(2016年7期)2017-01-09
江苏农业科学(2015年11期)2016-01-27
江苏农业科学(2015年11期)2016-01-27
河北渔业(2015年10期)2015-10-15
江苏农业科学(2015年8期)2015-09-10
