
棉花海陆回交群体盛铃期的光合特性及其生理基础
2024-04-29 00:26李生梅庞博耿世伟宋武李红梅马茂森张茹王新燕高文伟
中国农业科技导报 2024年1期
李生梅 庞博 耿世伟 宋武 李红梅 马茂森 张茹 王新燕 高文伟
摘要:为探究棉花海陆回交群体盛铃期的光合特性及其生理基础,筛选海陆杂交后代高光效种质资源,以陆地棉‘系9为母本、海岛棉‘新海16为父本构建的115份BC4F2:4株系群体为研究对象,测定其在田间条件下的光合参数及生理生化指标,利用描述性统计、相关性分析、回归分析、主成分分析和聚类分析评价子代株系的育种潜力。结果表明,回交群体的15个性状变异丰富,变异系数在8.96%~51.09%,其中净光合速率、蒸腾速率、水分利用效率、胞间CO2浓度、气孔导度、水蒸气压亏缺、丙二醛含量、可溶性蛋白含量和超氧化物歧化酶活性9个性状变异系数均在20%以上。相关性分析发现,海陆回交群体不仅在光合或生理生化指标各自内部存在显著相关性,在光合与生理生化指标之间也存在显著相关性,其中叶绿素相对值(soil and plant analyzer develotrnent,SPAD)与14个性状的相关性均达到显著水平。多元逐步回归分析构建出10个光合生理指标的最佳回归方程。主成分分析提取了5个主成分,累计贡献率达78.608%,第1和第4主成分有效解释群体生理指标,第2、第3和第5主成分有效解释群体光合性状。Ward.D层次聚类法将群体划分为4类,第Ⅰ、第Ⅱ、第Ⅲ和第Ⅳ类群占总材料的百分比分别为33.91%、10.43%、29.57%和26.09%,且第Ⅳ类群的30份材料是综合性状较好的棉花种质,可作为适合育种需要的高光效品系。研究结果为棉花高光效育种提供了理论依据及材料支撑。
关键词:海陆杂交棉;光合生理特性;回归分析;主成分分析;聚类分析
doi:10.13304/j.nykjdb.2022.0343
中图分类号:S562 文献标志码:A 文章编号:10080864(2024)01004012
叶片是植物光合作用制造养分的重要场所,是植物重要的营养器官之一,叶片具有光合作用、蒸腾作用、吸收作用、繁殖作用和储藏作用。叶片的净光合速率(net photosynthetic rate, Pn)、叶绿素(chlorophyll, Chl)、可溶性蛋白(soluble protein,SP)、超氧化物歧化酶(superoxide dismutase, SOD)和丙二醛(malondialdehyde , MDA)等指标可以反映其光合性能和衰老状况,从而影响作物的产量和品质[12]。棉花是世界上最重要的经济作物之一[3],陆地棉(Gossypium hirsutum L.)和海岛棉(Gossypium barbadense L.)是2个重要的异源四倍体栽培种,利用海陆种间杂交拓宽陆地棉栽培品种遗传基础是棉花育种的重要途径[4]。此外,开花和结铃阶段是决定棉花产量的关键时期[5]。因此,探究棉花海陆群体盛铃期的光合特性及其生理基础,评价杂交品系的育种潜力,从而筛选出高光效品种(系),对棉花产量和品质的提升具有重要意义。
目前,已有不少学者对多种植物的光合生理特性进行了分析。卢华雨等[6]对209株F2杂交高粱的主要光合性状进行相关性分析发现,蒸腾速率、气孔导度及胞间CO2浓度的提高会明显增加高粱的净光合速率。王建丽等[7]对杂交苜蓿的光合生理特性进行相关分析和回归分析发现,不同生育期对叶片的净光合速率和蒸腾速率起主导作用的环境因子有所差异。韦献东等[8] 对金花茶和山茶的光合生理特性进行主成分分析发现,金花茶耐荫性评价的生理生化指标是水分利用率(water use efficiency,WUE)、蒸腾速率(transpiration rate,Tr)、Pn 和SP,而山茶的评价指标是脯氨酸(proline,Pro)、MDA、过氧化氢酶(catalase,CAT)、过氧化物酶(peroxidase,POD)、SOD、WUE 和SP。牛宁等[9]对150 份来自黄淮海地区的大豆种质资源的光合性状进行主成分分析和聚类分析,选出气孔、水分和光合3个主成分因子,并筛选出23份高光效大豆种质。冯方剑等[10]利用32份棉花品种进行生理指标的主成分和聚类分析发现,叶绿素含量、SOD、POD、Pro、MDA和SP均可作为棉花苗期抗旱性鉴定指标。王笑言等[11]对国内外种植的65份棉花进行光合性状聚类分析,筛选出5份材料可作为光合生理育种中高光合速率的特异种质资源。性状间的相关性在植物育种中具有重要意义,在育种选择过程中可以根据性状间的简单相关性,测量相对简便的性状从而进行目标性状选择,进而提高选种效率[12]。系统聚类法能综合考虑不同指标的作用和影响,有利于综合性状的选育[13]。文国吉等[14]认为,棕色杂交棉高产优质的原因是叶片叶绿素质量分数较高,使叶绿体希尔反应活力增强,淀粉质量分数增大,从而使其净光合速率升高,致使营养物质供应充足。金珠群等[15]和李伶俐等[16]发现,杂交棉取得高产的主要机理是棉花生育后期叶片的叶绿素含量高、光合速率强、SOD 与POD 活性高且MDA含量低。此外,光合性能常作为筛选高光效品种的参考指标。因此,研究棉花盛铃期叶片光合生理特性的遗传变异规律和杂种优势表现,可起到早期鉴定和预先选择的作用。
棉花海陆种间杂交群体不仅具有陆地棉丰产特性和广泛的适应性,还具有海岛棉优良纤维品质。海岛棉与陆地棉的杂交已进行了多年,目前对海陆群体的研究主要集中在农艺性状及产量和纤维品质方面[1718],有关海陆杂交后代光合生理特性的报道较少。因此,本研究以海岛棉和陆地棉回交的115份后代株系为材料,对其盛铃期叶片的光合生理指标进行系统测定和遗传变异分析,旨在了解回交子代叶片光合生理的变异特征,探明其遗传规律,以期筛选出光合与生理综合性状较好的品种(系),为海陆群体高产生理研究及高光效育种提供理论依据与技术参考。
1 材料与方法
1.1 试验材料
供试材料为以陆地棉‘系9和海岛棉‘新海16为亲本构建的115份BC4F2:4株系,材料均由新疆农业大学农学院棉花遗传育种课题组提供,其中,‘系9为轮回亲本(母本),‘新海16为供体亲本(父本)(表1)。
1.2 试验设计
田间试验于2018年在沙湾144团新疆农业大学棉花基地(43°20′—45°20′N、84°45′—86°40′E)进行,2018年4月21日播种,采用随机区组设计,1膜6行种植模式,每份材料种植2行,行长2.0 m,株距0.1 m,行距0.7 m,播幅2.35 m,常规田间管理。
1.3 测定项目与方法
1.3.1 光合参数的测定 2018年7月29日至8月2日(盛铃期),选取生长一致的连续5株棉株的倒3功能叶进行光合生理指标的测定。采用英国汉莎CIRAS-3便携式光合仪,在10:30—13:00(此时间段为当地测量最佳时间)对棉花叶片进行光合参数的测定[19],测定时设定光量子通量密度为1 100 μmol·m-2·s-1,环境温度为(31±2)℃,相对湿度为50%±3%,CO2 浓度为(370±5)μmol·mol-1。主要测定倒3功能叶的净光合速率(Pn)、蒸腾速率(Tr)、水分利用效率(WUE)、胞间CO2 浓度(intracellularcarbon dioxide concentration, Ci)、气孔导度(stomatalconductance, Gs)、水蒸气压亏缺(vapour pressuredeficit,VPD)。采用SPAD-502便携式叶绿素测定仪测定倒3功能叶的叶绿素相对值(soil and plantanalyzer develotrnent, SPAD)值。WUE=Pn/Tr (1)
1.3.2 生理生化指标的测定 测定SPAD值和光合参数后,摘取相应叶片用锡箔纸包好并用防水马克笔做好标记置于液氮保存,以备后续测定生理生化指标。叶绿素采用简易提取法(95%乙醇避光浸提48 h,期间注意振荡)提取,采用多功能酶标仪(Synergy H1,美国)在波长663、645 和470 nm 下测定OD 值,计算叶绿素a(Chla)、叶绿素b(Chlb)、总叶绿素(ChlT)、叶绿素a/b(Chla/b)和类胡萝卜素(carotene,Car)含量;丙二醛(MDA)含量采用硫代巴比妥酸法测定,在波长450、532、600 nm下测定OD值;可溶性蛋白(SP)含量采用考马斯亮蓝法测定,在波长595 nm下测定OD值;超氧化物歧化酶(SOD)活性采用氮蓝四唑法测定,在波长560 nm下测定OD值;每个棉花株系的生理指标重复测定3 次,具体步骤参照孙丰磊等[19]的方法。
1.3.3 农艺性状和纤维品质的分析 海陆群体农艺性状(株高、有效成铃数、衣分)和纤维品质(纤维长度、纤维强度)的表型参照李生梅等[20]测定的数据进行分析。
1.4 数据处理与分析
试验数据使用Excel 2010 整理,采用SPSS26.0和R 3.6.0软件进行描述性统计、频率分析、相关性分析、逐步回归分析、主成分分析和聚类分析,采用Duncan法进行多重比较。
2 结果与分析
2.1 海陆群体盛铃期光合生理指标的遗传变异分析
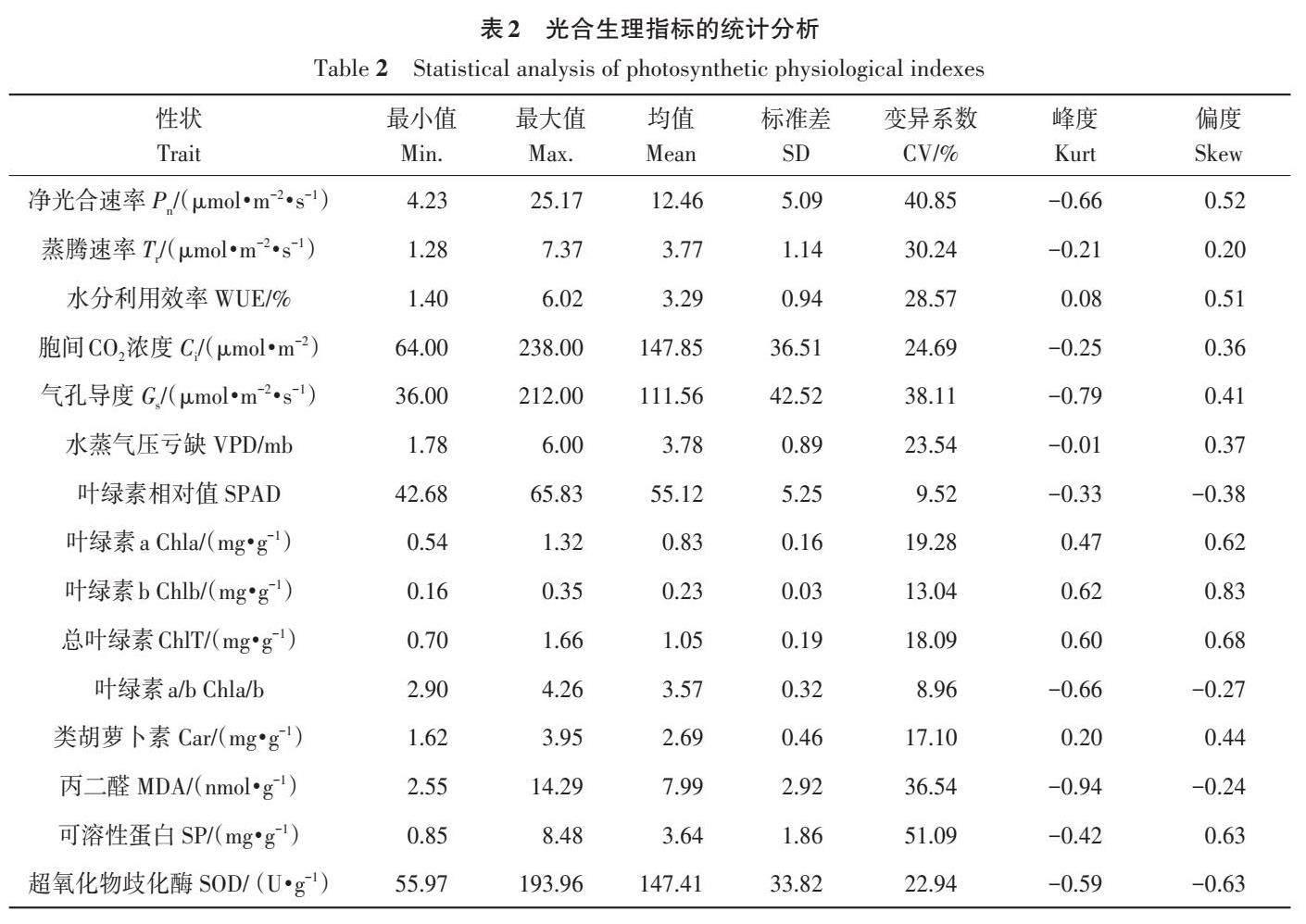
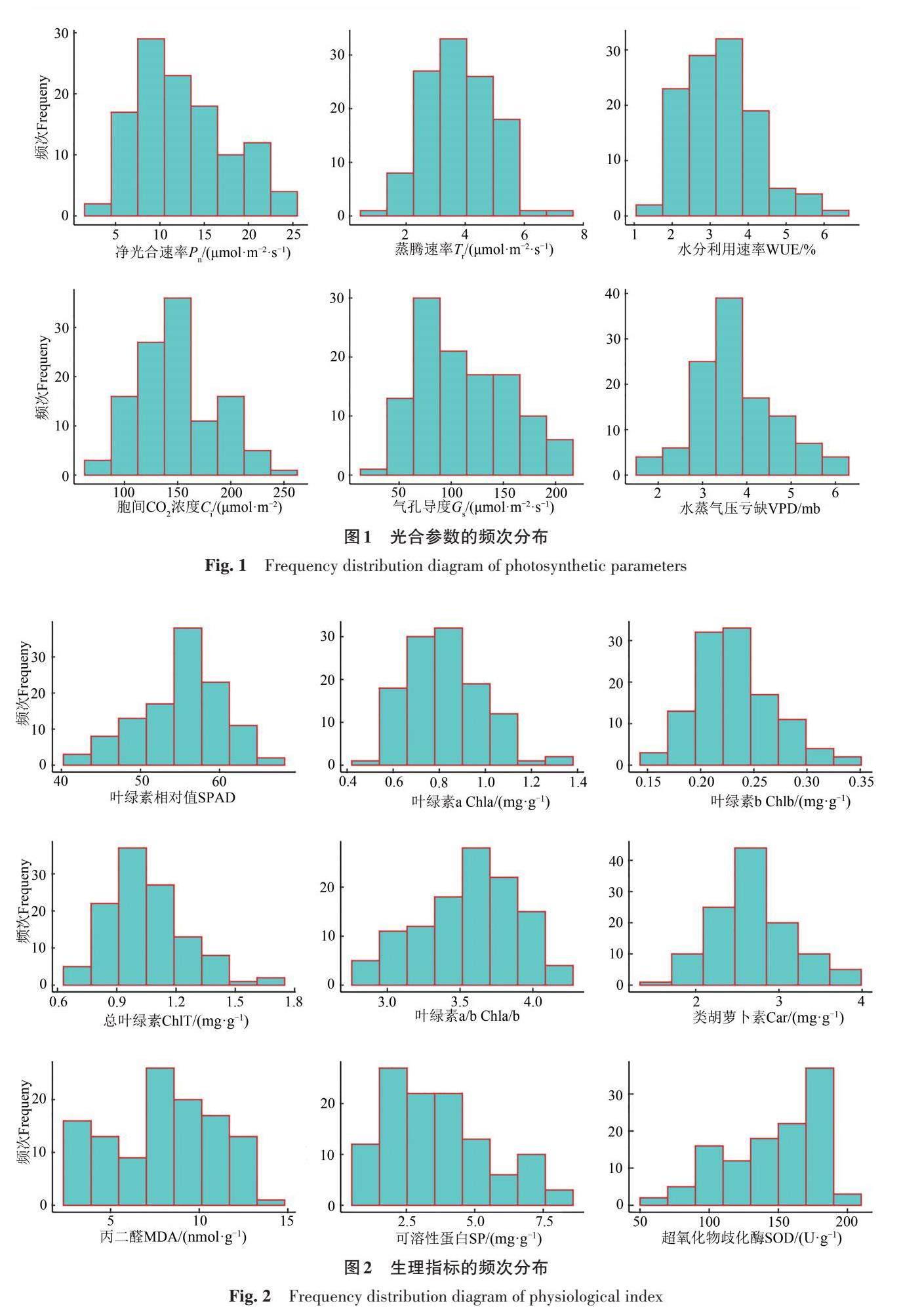
由表2可知,海陆杂交回交群体15个光合生理指标的变异系数在8.96%~51.09%,其中可溶性蛋白含量和净光合速率的变异系数较大,分别为51.09%和40.85%,其次为气孔导度、丙二醛含量、蒸腾速率、水分利用效率、胞间CO2浓度、水蒸气压亏缺以及超氧化物歧化酶活性,这9个性状的变异系数均在20%以上,这些性状在群体中存在广泛的遗传变异,说明株系间差异较大。
棉花叶片光合生理指标的峰度和偏度(表2)的绝对值均小于1,结合15个性状的频次分布(图1、2)可知,海陆群体的各性状在盛铃期近似服从于连续性正态分布。
2.2 海陆群体盛铃期光合生理指标的相关性分析
对15个光合生理指标进行相关性分析(表3)发现,净光合速率(Pn)与蒸腾速率(Tr)、水分利用效率(WUE)、气孔导度(Gs)、叶绿素相对值(SPAD)、叶绿素a(Chla)、总叶绿素(ChlT)、叶绿素a/b(Chla/b)呈极显著正相关;与胞间二氧化碳浓度(Ci)和水蒸气压亏缺(VPD)呈极显著负相关。蒸腾速率与气孔导度(Gs)、叶绿素a/b(Chla/b)、超氧化物歧化酶活性(SOD)呈极显著正相关。水分利用效率(WUE)与蒸腾速率(Tr)、叶绿素b(Chlb)、类胡萝卜素(Car)、丙二醛(MDA)和可溶性蛋白(SP)无显著相关性。与胞间二氧化碳浓度(Ci)呈显著相关的性状均为负相关。水蒸气压亏缺(VPD)与所有光合性状和超氧化物歧化酶(SOD)呈显著相关;叶绿素含量(SPAD)与绝大多数性状均达到显著或极显著水平;丙二醛(MDA)只与叶绿素呈显著负相关,可溶性蛋白(SP)与部分生理指标呈显著正相关,与光合性状均不相关。
2.3 海陆回交群体盛铃期光合生理指标的多元逐步回归分析
为进一步验证上述结果,对光合和生理指标进行逐步回归分析,筛选有显著影响的因素,剔除影响不显著的因素,构建最佳回归方程。由表4可知,每个回归方程都具有极显著的统计学意义(P<0.01),其中,Gs、Ci、WUE、Tr和VPD组合能解释Pn96.2%的总变异;Pn、WUE和Chla/b组合能解释Tr变异的92.6%;VPD、Ci、Pn、Tr 和SOD 组合能解释WUE变异的93.4%;Pn、Gs、VPD和WUE组合能解释Ci变异的68.5%;Pn、Ci、MDA和Tr组合能解释Gs变异的76.2%;WUE、Ci和SP组合能解释VPD变异的81.6%;Chla、Pn、Car 和MDA 能共同解释SPAD变异的36.2%;Chlb、SP、ChlT、Chla/b、SOD和Tr能共同解释Car变异的77.6%;SPAD能够单独解释MDA 变异的9.1%;ChlT 和Car 能解释SP 变异的11.7%;VPD、SP、Tr和Chla/b能共同解释SOD变异的19.7%,说明建立的线性回归模型对海陆群体的光合生理指标具有重要的参考意义。
2.4 海陆回交群体盛铃期光合生理指标的主成分分析
对群体的115份光合生理指标进行主成分分析(表5),提取特征值>1.0的主成分,结果表明,提取的5个主成分特征值分别是4.835、2.663、1.873、1.356和1.064,贡献率分别是24.411%、18.264%、14.630%、12.198% 和9.105%,累计贡献率达78.608%,包含了光合生理指标的绝大部分信息。
主成分1中叶绿素a、叶绿素b和总叶绿素特征向量值较大,说明主成分1主要由叶绿素相关指标决定;主成分2中净光合速率和气孔导度特征向量值最大,主成分3中水蒸气压亏缺和蒸腾速率具有正向较大载荷,说明第2主和第3主成分可以划分群体子代株系的光合性状;主成分4中可溶性蛋白具有负向最大特征向量值,超氧化物歧化酶和丙二醛具有正向较大值,说明主成分4主要由生理生化相关指标决定;主成分5主要由胞间二氧化碳浓度(Ci)决定。
2.5 海陆回交群体盛铃期光合生理指标的聚类分析
利用R语言软件对115份子代株系进行系统聚类,根据Ward.D层次聚类法将群体划分为4个类群(图3),分别计算各类群性状的平均值,并进行方差分析(表6)。结果表明,第Ⅰ类群包含39份材料,占海陆群体的33.91%,主要特征为净光合速率较低(10.04)、水分利用效率低(2.60),水蒸气压亏缺(4.48)和超氧化物歧化酶活性高(156.00),对于这类材料在种植过程中应注意适时灌溉。第Ⅱ类群包含12份材料,占海陆群体的10.43%,主要特征为净光合速率低(6.43)、蒸腾速率低(2.16)、气孔导度低(65.27)、水蒸气压亏缺低(3.15)、叶绿素含量低(47.84)、可溶性蛋白低(2.13),胞间二氧化碳浓度(191.83)和丙二醛含量高(9.39)。第Ⅲ类群包含34份材料,占海陆群体的29.57%,主要特征为净光合速率较高(13.81)、类胡萝卜素高(3.20)、可溶性蛋白高(4.16),胞间二氧化碳浓度(133.03)及丙二醛含量低(6.82)。第Ⅳ类群包含30份材料,占海陆群体的26.09%,主要特征为净光合速率高(16.50)、蒸腾速率高(4.23)、水分利用效率高(3.96)、气孔导度高(142.72)、叶绿素含量高(57.35),过氧化物歧化酶活性低(133.39),类群Ⅳ材料综合性状较好,可作为育种需要的高光效品系。此外,结合类群Ⅳ的农艺及品质田间表型[23],发现HL-15和HL-215株系成铃数多,HL-76株系株型高,HL-52和HL-75株系具有高衣分和强纤维。
3 讨 论
光合生理特性是作物生长和产量形成的重要生理基础。光合作用是棉花生长发育必不可少的生理活动,它与叶绿素含量、叶绿体结构、渗透调节物质等密切相关,极易受外界环境的影响,从而影响棉花的产量和品质[2122]。王继庆等[23]研究表明,群体性状变异程度越大,对种质变异和创新贡献率就越高。本研究通过对海陆群体盛铃期的15个光合生理指标的遗传变异进行分析发现,变异范围在8.96%~51.09%,可溶性蛋白和净光合速率的变异系数最大,这与Lamine[24]对玉米F1的光合生理遗传倾向研究结果一致,说明群体光合遗传基础较广泛,有利于种质材料的筛选。
净光合速率可反映光合能力的强弱[21];气孔是植物体内与外界环境进行水气交换的通道,气孔的大小、数量和调节功能与蒸腾作用等生理过程有关;叶绿素含量是植物生理特性的重要指标;超氧化物歧化酶(SOD)是一种抗氧化酶,可以延缓植株衰老,SOD活性越高对于植株的生长发育越有利[2526];MDA能够引起细胞膜功能紊乱,MDA含量越高对植株生长越不利,这些渗透调节物质及叶绿素含量等会影响植物的形态和生理生化过程,进而影响光合产物的积累[2728]。因此,净光合速率与其他指标的相关性是品种选育的重要参照。本研究相关性分析显示,海陆群体中Pn与Tr、WUE、Gs、SPAD、Chla、ChlT 和Chla/b 呈极显著正相关,与Ci和VPD呈极显著负相关,与Chlb、Car、SP和SOD呈正相关,与MDA呈负相关。索荣臻等[13]发现,Pn与CE、WUE和Tr呈极显著正相关,与Ci和SPAD呈极显著负相关;Lamine[24]研究杂交玉米认为,Pn与SPAD呈显著正相关;江锡兵等[29]通过种间杂交板栗发现,Pn 与Gs、Tr、WUE、Chla、Chlb和ChlT呈极显著正相关;郭丽丽等[30]对牡丹的研究结果表明,Pn与叶绿素和SP呈极显著正相关,叶绿素与SP也是呈极显著正相关;Verma等[31]研究甘蔗发现,Pn 与Gs、Tr、Chla、Chlb、ChlT 和SPAD呈极显著正相关,与SP和SOD呈极显著负相关,与MDA呈负相关。本研究与上述研究的结果并不完全相同,可能是由于研究对象不同。同时,基于以上结果本研究发现在大多数植物中Pn与Tr 和水分利用效率(WUE)均呈正相关。为使试验结果更具准确性和代表性,本研究进一步构建出10个指标的最佳回归方程,后期进行目标性状选择时,建立的线性回归模型可作为辅助参考,有利于提高选择效率。
主成分分析可以将多个指标转化为少数几个不相关的综合指标,利用该方法可以评价作物品种各综合指标的主成分,可为合理选配亲本及快速筛选评价指标提供理论依据。本研究主成分结果表明,海陆群体被划分为5个主成分,累计贡献率达78.608%,其中,主成分1(Chla、Chlb、ChlT)和主成分4(SP、SOD、MDA)主要解释海陆群体的生理指标,主成分2(Pn、Gs)、主成分3(VPD、Tr)和主成分5(Ci)主要解释海陆群体的光合性状。为更真实地反映品种的综合性状,需进一步采用聚类分析[32]进行划分。姜梦辉等[33]通过聚类分析对海陆群体的抗旱等级进行了划分。本研究通过Ward.D 层次聚类法将115 份材料划分为4 个类群,结果清晰且简便可靠,便于海陆群体的综合评价。其中,第Ⅳ类群包含30份材料,主要特征为净光合速率高、蒸腾速率高、水分利用效率高、气孔导度高、叶绿素含量高和超氧化物歧化酶活性低。提高叶片光合特性和活性氧清除酶活性对叶片衰老有显著的调控效应,有利于提高作物产量[34]。因此,海陆群体类群Ⅳ材料综合性状较好,可作为品种选育目标,但是此类材料活性氧清除酶活性较低,因此,可以从第Ⅲ类群中挑选SOD活性高和MDA含量低的材料作为备选材料,以期发现适合育种需要的高光效品系。本研究仅解析了海陆群体的光合及生理特性,初步结合了光合生理指标综合性状较好的类群Ⅳ进行田间表型观察,发现不同株系表现不一,包括成铃数多少、株型高矮,衣分高低和品质优劣。因此,在今后的研究中将从光合生理角度分析海陆群体产量提高和品质改良的机制,进一步为海陆群体育种提供理论依据。
综上所述,海陆群体盛铃期的光合生理遗传多样性丰富,依据10个光合生理指标的辅助参考模型,有利于提高目标性状的选择效率。海陆群体类群Ⅳ的30份材料在盛铃期综合性状表现较好,在育种中作为亲本选择时应加以重点应用。
参 考 文 献
[1] 张咪咪, 何丽娜, 何觉民, 等. 小麦与野燕麦杂交后代品系光合特性的研究[J]. 麦类作物学报, 2012, 32(5): 945-948.
ZHANG M M, HE L N, HE J M, et al .. Study on thephotosynthetic performance of the new lines developed fromthe cross between Triticum aestivum and Avena fatua [J]. J.Triticeae Crops, 2012, 32(5): 945-948.
[2] 董合忠, 李维江, 唐薇, 等. 棉花生理性早衰研究进展[J]. 棉花学报, 2005, 17 (1): 56-60.
DONG H Z, LI W J, TANG W, et al .. Research progress inphysiological premature senescence in cotton [J]. Cott. Sci.,2005, 17 (1): 56-60.
[3] LI S M, GENG S W, PANG B, et al .. Revealing geneticdifferences in fiber elongation between the offspring of seaisland cotton and upland cotton backcross populations basedon transcriptome and weighted gene coexpression networks [J/OL].Genes, 2022, 13(6): 954 [2022-03-23]. https://doi.org/10.3390/genes13060954.
[4] 朱协飞, 王鹏, 司占峰, 等. 基于陆地棉背景的海岛棉染色体片段导入系产量性状QTL 定位[J]. 作物学报, 2017,43(12): 1784-1790.
ZHU X F, WANG P, SI Z F, et al .. QTL Mapping for yieldcomponents in Gossypium barbadense chromosome segmentintrogression lines based on Gossypium hirsutum background [J].Acta Agron. Sin., 2017, 43(12): 1784-1790.
[5] ZHANG L, ZHANG G W, WANG Y H, et al .. Effect of soilsalinity on physiological characteristics of functional leaves ofcotton plants [J]. J. Plant Res., 2013, 126(2): 293-304.
[6] 卢华雨, 李延玲, 罗峰, 等. 粒用高粱4个主要光合性状数量遗传分析[J]. 江苏农业科学, 2018, 46(17): 76-80.
LU H Y, LI Y L, LUO F, et al .. Genetic analysis of four mainphotosynthetic characters of grain sorghum [J]. Jiangsu Agric.Sci., 2018, 46(17): 76-80.
[7] 王建丽, 张永亮, 朱占林, 等. 杂花苜蓿叶片光合生理生态特性[J]. 草地学报, 2006, 14(2): 138-141.
WANG J L, ZHANG Y L, ZHU Z L, et al .. Analysis onecophysiological characteristics of leaf photosynthesis ofMedicago varia. cv. gannong [J]. Acta Agrestia Sin., 2006, 14(2):138-141.
[8] 韦献东, 陶志华, 王艺锦, 等. 郁闭度对金花茶和山茶的生理特性影响及主成分分析[J]. 福建农业学报, 2019, 34(8):905-911.
WEI X D, TAO Z H, WANG Y J, et al .. Effects of canopydensity on physiology and biochemistry of Camellia nitidissimachi and Camellia Japonica L. as evaluated by principalcomponent analysis [J]. Fujian J. Agric. Sci., 2019, 34(8):905-911.
[9] 牛宁, 李振侠, 金素娟, 等. 黄淮海地区大豆光合特性及高光效种质筛选[J]. 中国油料作物学报, 2018, 40(4): 524-532.
NIU N, LI Z X, JIN S J, et al .. Comparison of photosyntheticgas exchange parametersfor 150 soybean germplasm fromHuanghuaihai region [J]. Chin. J. Oil Crop Sci., 2018, 40(4):524-532.
[10] 冯方剑, 宋敏, 陈全家, 等. 棉花苗期抗旱相关指标的主成分分析及综合评价[J]. 新疆农业大学学报, 2011, 34(3):211-217.
FENG F J, SONG M, CHEN Q J, et al .. Analysis andcomprehensive evaluation on principal component of relativeindices of drought resistance at the seedling stage of cotton [J].J. Xinjiang Agric. Univ., 2011, 34(3): 211-217.
[11] 王笑言, 张春梅, 魏亦农, 等. 新疆滴灌条件下65份棉花资源材料的光合速率[J]. 棉花学报, 2006, 18(6): 396-397.
WANG X Y, ZHANG C M, WEI Y N, et al .. Photosyntheticrates of 65 cotton varieties under mulch-drip irrigation inXinjiang [J]. Cott. Sci., 2006, 18(6): 396-397.
[12] 李华丽, 陈美霞, 阮奇城, 等. 烟草F2:3家系7个主要产量性状的遗传变异分析[J]. 中国农学通报, 2011, 27(16): 94-98.
LI H L, CHEN M X, RUAN Q C, et al .. Genetic variationanalyze of seven yield traits of F2:3 family in tobacco [J]. Chin.Agric. Sci. Bull., 2011, 27(16): 94-98.
[13] 索荣臻, 王明玖, 赵天启, 等. 野生大豆与栽培大豆杂交后代不同品种(系)的光合特性比较与聚类分析[J]. 中国油料作物学报, 2020, 42(2): 255-263.
SUO R Z, WANG M J, ZHAO T Q, et al .. Comparison ofphotosynthetic characteristics and cluster analysis in Glycinesoja and strains from Glycine soja × Glycine max cross [J].Chin. J. Oil Crop Sci., 2020, 42(2): 255-263.
[14] 文国吉, 袁淑娜, 刘英新, 等. 棕色杂交棉高产优质的光合生理特性[J]. 浙江大学学报(农业与生命科学版), 2011,37(1): 54-60.
WEN G J, YUAN S N, LIU Y X, et al .. Photosyntheticphysiological characteristics of brown-colored hybrid cottonwith high yield and good quality [J]. J. Zhejiang Univ. (Agric.Life Sci.), 2011, 37(1): 54-60.
[15] 金珠群, 陈仲华, 黄一青, 等. 抗虫杂交棉慈抗杂3号若干生育与生理特性的杂种优势研究[J]. 棉花学报, 2004, 16(6):347-351.
JIN Z Q, CHEN Z H, HUANG Y Q, et al .. Studies on heterosisof growth and some physiological traits in hybrid Bt-cottoncikangza 3 [J]. Cott. Sci., 2004, 16(6): 347-351.
[16] 李伶俐, 林同保, 房卫平, 等. 杂交棉叶片衰老特点及高产生理机制研究[J]. 河南农业大学学报, 2006, 40(4): 341-345.
LI L L, LIN T B, FANG W P, et al .. Research on high yieldphysiological mechanism and leaf senescence characteristicsof hybrid cotton [J]. J. Henan Agric. Univ., 2006, 40(4):341-345.
[17] SHI Y Z, LIU A Y, LI J W, et al .. Dissecting the genetic basisof fiber quality and yield traits in interspecific backcrosspopulations of Gossypium hirsutum × Gossypium barbadense [J].Mol. Gene Genom., 2019, 294(6): 1385-1402.
[18] MD HARUN OR RASHID. 利用CSSLs进行棉花抗黄萎病、纤维品质和产量性状全基因组QTL定位[D]. 北京: 中国农业科学院, 2017.
RASHID M H O. Genome wide QTL mapping for resistance toVerticillium wilt, fiber quality and yield traits in cottonchromosome segment substitution lines [D]. Beijing: ChineseAcademy of Agricultural Sciences, 2017.
[19] 孙丰磊, 张玻, 曲延英, 等. 花铃期干旱胁迫对不同棉花品种光合特性影响及抗旱性评价[J]. 干旱地区农业研究,2018, 36(5): 7-13, 20.
SUN F L, ZHANG B, QU Y Y, et al .. Effects of drought stressduring the blooming period on photosynthetic characteristicsand assessment drought resistance of different cotton varieties [J].Agric. Res. Arid Areas, 2018, 36(5): 7-13, 20.
[20] 李生梅, 杨涛, 黄雅婕, 等. 海陆回交群体主要农艺性状与纤维品质关系的探讨[J]. 中国农业科技导报, 2021, 23(8):16-24.
LI S M, YANG T, HUANG Y J, et al .. Discussion on therelationship between main agronomic traits and fiber quality inbackcross populations of Gossypium hirsutum L. × Gossypiumbarbadense L. [J]. J. Agric. Sci. Technol., 2021, 23(8): 16-24.
[21] 王丽妍, 徐宝慧, 杨成林, 等. 北方地区不同花生品种光合生理特性的比较[J]. 华南农业大学学报, 2010, 31(4): 12-15.
WANG L Y, XU B H, YANG C L, et al .. Comparative studieson photosynthetic physiological characteristics of differentpeanut varieties in northern region of China [J]. J. South ChinaAgric. Univ., 2010, 31(4): 12-15.
[22] 葛勇, 李威, 刘艳改, 等. 中棉所63棉铃对位叶光合生理性能的杂种优势研究[J]. 棉花学报, 2016,28(3): 250-259.
GE Y, LI W, LIU Y G, et al .. Heterosis studies onphotosynthetic and physiological performance of thesubtending leaf of cotton boll of CCRI 63 [J]. Cott. Sci., 2016,28(3): 250-259.
[23] 王继庆, 任毅, 时晓磊, 等. 小麦籽粒超氧化物歧化酶(SOD)活性全基因组关联分析[J]. 中国农业科学, 2021, 54(11):2249-2265.
WANG J Q, REN Y, SHI X L, et al .. Genome-wide associationanalysis of superoxide dismutase (SOD) activity in wheat grain [J].Sci. Agric. Sin., 2021, 54(11): 2249-2265.
[24] DIALLO LAMINE. 玉米自交系及F1 代生理遗传倾向研究[D]. 长春:吉林大学, 2016.
LAMINE D. Genetic analysis of the photosyntheticcharacteristics of maize inbred lines and their hybrids [D].Changchun: Jilin University, 2016.
[25] CASSON S, GRAY J E. Influence of environmental factors onstomatal development [J]. New Phytol., 2008, 178(1): 9-23.
[26] WANG H F, HUO Z G, ZHOU G S, et al .. Estimating leafSPAD values of freeze-damaged winter wheat using continuouswavelet analysis [J]. Plant Physiol. Biochem., 2016, 98: 39-45.
[27] CHEN J, WANG W H, LIU T W, et al .. Photosynthetic andantioxidant responses of Liquidambar formosana and Schimasuperba seedlings to sulfuric-rich and nitric-rich simulatedacid rain [J]. Plant Physiol. Biochem., 2013, 64: 41-51.
[28] 陈文杰, 郭小红, 汤复跃, 等. 不同生育时期大豆叶片光合特性变化及相关性研究[J]. 中国油料作物学报, 2017, 39(3):360-365.
CHEN W J, GUO X H, TANG F Y, et al .. Changes andrelationship of photosynthetic traits of soybean leaves atdifferent growth stages [J]. Chin. J. Oil Crop Sci., 2017, 39(3):360-365.
[29] 江锡兵, 章平生, 杨龙, 等. 板栗和锥栗种间杂交F1代叶片表型及其遗传变异研究[J]. 园艺学报, 2019, 46(11): 2129-2142.
JIANG X B, ZHANG P S, YANG L, et al .. Genetic variation ofleaf phenotypic traits in F1 progeny of interspecific crossbetween castanea mollissima and C. henryi [J]. Acta Hortic.Sin., 2019, 46(11): 2129-2142.
[30] 郭丽丽, 郭大龙, 赵威, 等. 不同品种油用型牡丹的光合与生理特性及其与结实率的相关性分析[J]. 上海农业学报,2017, 33(3): 54-59.
GUO L L, GUO D L, ZHAO W, et al .. The photosynthetic andphysiological characteristics of different oil tree peony varietiesand their correlation between seed setting rate [J]. Acta Agric.Shanghai, 2017, 33(3): 54-59.
[31] VERMA K K, SONG X P, ZENG Y, et al .. Foliar application ofsilicon boosts growth, photosynthetic leaf gas exchange,antioxidative response and resistance to limited water irrigation in sugarcane (Saccharum officinarum L.) [J]. Plant Physiol.Biochem., 2021, 166: 582-592.
[32] 陈华萍, 王照丽, 魏育明, 等. 四川小麦地方品种农艺性状与品质性状的聚类分析[J]. 麦类作物学报, 2006, 25 (6):29-34.
CHEN H P, WANG Z L, WEI Y M, et al .. Cluster analysis ofagronomic and quality characters in si chuan wheat landraces [J]. J.Triticeae Crops, 2006, 25 (6): 29-34.
[33] 姜梦辉, 孙丰磊, 杨阳, 等. 棉花陆海重组自交系群体花铃期抗旱性鉴定及评价[J]. 干旱区研究, 2020, 37(6): 1635-1643.
JIANG M H, SUN F L, YANG Y, et al .. Identification andevaluation of drought resistance of upland-island recombinationinbred line population at blossoming and boll-forming stages [J].Arid Zone Res., 2020, 37(6): 1635-1643.
[34] 谭彩霞, 封超年, 郭文善, 等. 不同品质类型小麦旗叶光合特性及其与产量的相关性研究[J]. 扬州大学学报(农业与生命科学版), 2019, 40(6): 30-34.
TAN C X, FENG C N, GUO W S, et al .. Photosyntheticphysiological characteristics in flag leaf of different qualitytypes of wheat and its correlation with yield [J]. J. YangzhouUniv. (Agric. Life Sci.), 2019, 40(6): 30-34.
(责任编辑:胡立霞)
猜你喜欢
大学教育(2016年11期)2016-11-16
大经贸(2016年9期)2016-11-16
中小企业管理与科技·上旬刊(2016年10期)2016-11-15
价值工程(2016年29期)2016-11-14
考试周刊(2016年84期)2016-11-11
中国市场(2016年33期)2016-10-18
中国市场(2016年33期)2016-10-18
商(2016年27期)2016-10-17
科技视界(2016年20期)2016-09-29
企业导报(2016年9期)2016-05-26
