
不同铁环境对蓝莓生长及叶片叶绿素荧光特性的影响
2024-04-29 00:26杨圣艳曹漫郭宝石杨超侯智霞
中国农业科技导报 2024年1期
杨圣艳 曹漫 郭宝石 杨超 侯智霞
摘要:铁(Fe)是植物必需的微量元素,参与光合作用。为探究Fe对蓝莓(Vaccinium spp.)重要生命活动的影响,以蓝莓‘莱克西为试验材料,设置正常铁(Na-Fe-EDTA)、只有难溶铁(Fe2O3)及缺铁(不外加任何铁元素)共3种铁环境,分析其苗木生长发育及其叶片相对叶绿素含量(SPAD值)、叶绿素含量、叶绿素荧光参数等的差异。结果表明,在难溶铁和缺铁2种胁迫环境下,蓝莓各生长指标、光合色素含量及SPAD值均显著低于正常铁环境(P<0.05),而难溶铁环境下蓝莓苗木的基生枝数量和生长量、地径、叶面积及叶片光合色素含量显著低于缺铁环境(P<0.05);正常铁环境中叶片PSⅡ实际光合效率高于难溶铁和缺铁环境,而光抑制程度显著低于这2种铁胁迫环境;难溶铁、缺铁环境中的蓝莓苗木叶片受损、叶片的光合能力减弱,其中难溶铁环境与缺铁环境相比,光系统中心转换效率更不稳定,叶片受光抑制程度最高;叶面积、单株生物量、基生枝生长量与各叶绿素荧光参数间具有显著相关性(P<0.01),且叶片光合色素含量与SPAD值间呈显著正相关(P<0.01),但各光合色素和SPAD值与各荧光参数间相关性不显著。综上所述,难溶铁环境、缺铁环境使蓝莓苗木生长发育受抑制与该环境下造成叶片叶绿素含量降低有关,胁迫环境下叶片PSⅡ反应中心光能的吸收和转换减弱,电子传递受阻,光合效率降低,为蓝莓铁素营养合理利用方式的深入挖掘提供参考依据。
关键词:蓝莓;铁;叶绿素荧光;生长发育
doi:10.13304/j.nykjdb.2022.0490
中图分类号:S663 文献标志码:A 文章编号:10080864(2024)01005211
蓝莓是杜鹃花科(Ericaceae)越橘属(Vaccinium)多年生灌木,果实为蓝色浆果,富含维生素、膳食纤维和花青素等活性物质,具有极高的保健作用和经济价值。当前我国人工栽培蓝莓面积不断增加,已扩展至27 个省(市、自治区)[1]。蓝莓根系浅,以须根为主,没有根毛[2],对土壤条件要求比较高,易受土壤质地、酸碱度、通透性、有机质含量、菌根状况等理化性状的影响[3-5]。蓝莓适宜在强酸、湿润、有机质含量高、通气状况良好的环境条件下生长[6],适生土壤pH4.0~5.0[4]。铁(Fe)是植物正常生长所必需的微量元素,通常以Fe2+状态被植物吸收利用,在植物生命过程中起着不可或缺的作用[7]。土壤pH 7.0或7.5时蓝莓根系活力显著下降[8],根部铁的运转和吸收减少,导致地上部分出现缺铁性黄化,叶片表现为黄化、脉间失绿等症状。施用Na-Fe-EDTA螯合铁肥,能够有效地改善碱性土壤中蓝莓缺铁黄化现象[9],其中螯合铁浓度50 μmol·L-1是蓝莓生长发育较适宜的铁素水平[10]。地球上大多数铁以Fe3+的形式存在于矿物晶体中,生物有效性非常低[7]。为了适应环境,植物在进化过程中形成了不同的铁吸收机制,如蓝莓等双子叶植物和禾本科植物在吸收铁时通过活化根基铁,将根表Fe3+还原为Fe2+,吸收并运输至各器官供生长发育所需[1112]。
在植物体内,铁参与光合作用,植物体80%的铁分布于叶绿体,是合成叶绿素的必需元素[13]。而土壤供铁水平与叶绿素含量具有相关性,当植物受到缺铁胁迫时,叶片中类囊体膜数量减少、叶绿体结构异常,叶绿素含量及叶绿素相对含量(SPAD值)降低,叶片出现黄化现象[14-18]。叶绿素a是叶绿素荧光的主要来源,包括光反应和暗反应在内的多数光合过程的变化向光系统Ⅱ(photosystem Ⅱ complex, PSⅡ)反馈,从而引起叶绿素a荧光的变化,即几乎所有的光合作用过程都能通过叶绿素荧光反映。而叶绿素荧光参数较表观性气体指标更能反映植物叶片内在性特点,在光合作用生理生态和逆境生理等领域得到广泛研究。如黄宗安等[19]发现盘菜缺铁时PSⅡ最大光化学效率(maximum photochemical quantum yieldof PSⅡ, Fv/Fm)显著降低,非光化学猝灭系数(nonphotochemicalquenching, NPQ)升高;姜闯道等[20]研究发现,在Fe2+质量浓度小于0.01 g·L-1的环境下,缺铁导致叶片中剩下大量激发能,其NPQ显著高于对照组。在不同铁水平胁迫处理下,植物光能的捕获和传递能力减弱,PSⅡ反应中心受到损伤,导致激发能耗散增加、电子传递效率降低,叶片光保护能力减弱[2122]。
全球有1/3的土壤是石灰性土壤,缺铁问题比较严重。在蓝莓栽培过程中,其叶片多出现黄化现象,进而导致蓝莓果实产量和品质降低,本研究设置了不同铁素水平的培养环境,旨在通过检测植物的叶绿素荧光特性及叶绿素含量分析叶片光合能力的差异,结合苗木的生长特性,分析不同铁素环境对蓝莓苗木生长发育的影响,为蓝莓生产中合理利用铁素营养提供技术指导。
1 材料与方法
1.1 试验材料
以生长一致、健壮、无病虫害的1 年生蓝莓‘莱克西盆栽苗为试验材料。
1.2 试验设计
以100%蛭石为栽培基质,营养钵直径8 cm、高9 cm,栽植于室温25 ℃的植物培养室。设置3 种铁环境处理:①缺铁环境,不添加任何铁素营养(B1);②以正常生长的铁素环境做为对照,由50 μmol·L-1 Na-Fe-EDTA 提供Fe2+(B2);③难溶三价铁的环境,由50 μmol·L-1 Fe2O3 提供Fe3+(B3)。每种铁素环境3 次重复,每个重复20 株苗,3 种铁素环境共180 株,培育180 d。以改良的去铁离子Hogland营养液作为蓝莓生长的营养供给来源(表1),即除铁离子外,其他营养组成均相同,营养液pH 4.8[4]。培养期间每15 d 浇1 次营养液,每个营养钵浇150 mL·次-1,处理期间根据蓝莓生长需求适当浇水。培养120 d时开始整株取样,以后每30 d取1次,共取样3次。
1.3 测定方法
1.3.1 生长发育指标测定 每个处理随机选取12株,每30 d观察记录蓝莓生长发育状态,直至试验结束,并统计各处理基生枝数量、基生枝生长量(cm)、地径(mm),直至取样结束。称取各处理苗木的鲜重(g),并计算平均单株生物量(g)。计算各处理基生枝数量、基生枝生长量、地径的总和,作为形态生长分析的指标。随机选取新梢顶部向下第4~6片叶片测量叶面积(cm2),每个处理取24片,并计算其平均叶面积。
1.3.2 光合生理指标测定 ①叶绿素相对含量(即SPAD 值)的测定。用手持便携式SPAD-502Plus叶绿素仪由植株新枝顶部至下第4~6片叶进行SPAD值测定[23],测定时避开主叶脉,每片叶子选取3个点进行测量。
② 叶绿素a、叶绿素b 和总叶绿素的检测。将测完SPAD 值的叶片剪细碎,称取0.2 g 置于15 mL棕色离心管中,加入5 mL无水乙醇和5 mL80% 丙酮,将绿色叶片浸泡至白色,在663、646、470 nm波长下测定上清液的吸光值。色素含量的计算参照Lichtenthaler[24]的方法。
③叶绿素荧光参数的检测。利用便携式的脉冲振幅制叶绿素荧光仪PAM-2500测量植物的光合作用活性。首先,每个处理随机选择3株蓝莓苗,将其指定为测定叶绿素荧光参数的固定苗株。将蓝莓苗于夜间充分暗适应后(30 min以上),随机选取植株新梢自上向下第4~8片成熟叶,每个处理取12个避开主叶脉且不重复的位点,用暗适应叶夹夹住检测叶片,仪器发出检测光得到初始荧光产量(original fluorescence yield, Fo)、最大荧光产量(maximal fluorescence yield, Fm)和PSⅡ反应中心最大光能转换效率(Fv/Fm)。设置8个光强梯度,每个梯度照射10 s,利用光响曲线(lightcurve)检测到PS Ⅱ 实际光合效率(effectivephotochemical quantum yield of PSⅡ, Y(Ⅱ))、PSⅡ反应中心关闭程度(QA restore state, 1-qP)、非光化学淬灭系数(NPQ)、PS Ⅱ 电子传递速率(electron transport rate, ETR)和PSⅡ非调节性能量耗散的量子产量[quantum yield of non-lightmoluced non-photochemical fluorescence quenching,Y(NO)]等叶绿素荧光动力学参数,结果取均值。
1.4 数据分析
运用 Microsoft Excel 2016 整理生理指标,Origin 2021作相关图表。用软件ImageJ测量叶面积,采用 SPSS 26.0对蓝莓叶片的叶绿素(叶绿素a、叶绿素b、总叶绿素)含量及SPAD值、叶绿素荧光参数等进行方差分析(P<0.05),用Pearson分析法分析不同生长指标、叶绿素含量、SPAD值、叶面积及各叶绿素荧光参数间的相关性。
2 结果与分析
2.1 不同铁环境对蓝莓生长的影响
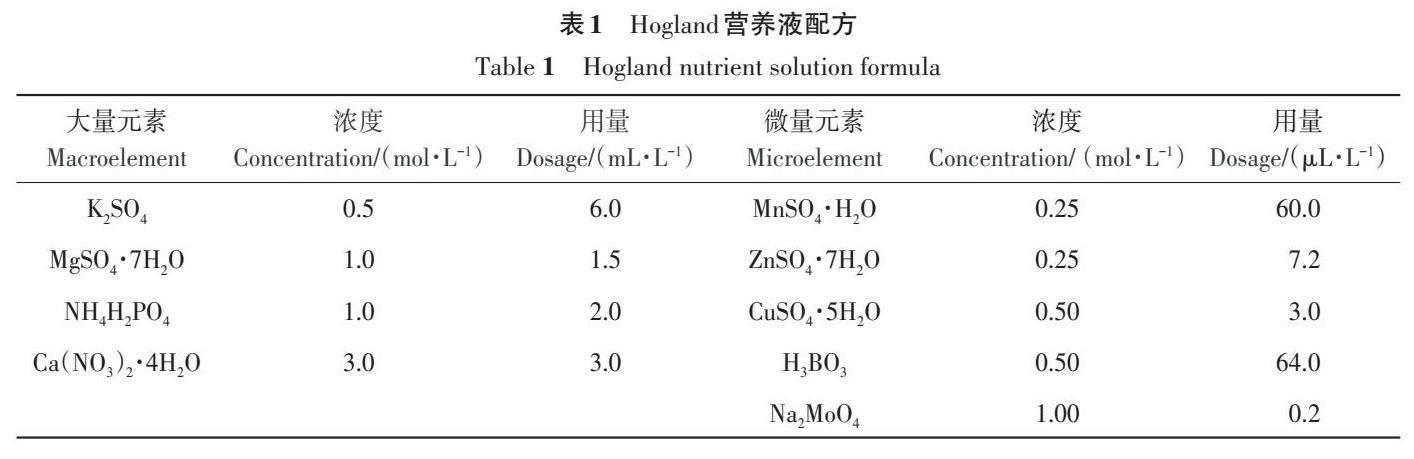
蓝莓为灌木,苗木萌蘖能力较强,试验之初选用的蓝莓苗仅具单枝,随着处理时间的延长苗木基生枝数量不断增加,统计各铁素环境下基部抽生总基生枝数量。由图1 可知,B2 处理在培育180 d后基生枝数量最多,而蓝莓在B1和B3铁胁迫处理后仍能继续生长,但生长势相比B2处理显著下降(P<0.05),且B3 处理基生枝数量显著少于B1处理(P<0.05)。培育期间,B2、B1处理在90~120 d内基生枝萌蘖能力最强,基生枝数量分别增加了26.67%、20.00%,而B3 处理培育180 d后仅增加了18.18%。
蓝莓基生枝生长量是反映其生长能力的基本指标。随着培育时间的延长,3种铁环境间基生枝生长量差异显著(P<0.05),蓝莓苗受B1、B3胁迫处理的影响越来越明显(图1),B1、B3处理基生枝生长量均显著小于B2,且B3处理显著小于B1(P<0.05)。培育180 d后,B2处理基生枝生长量最高,其生长量增加了101.65%,B1、B3处理分别增长了64.59%、54.22%。培育120 d 后,B2 处理地径显著大于B1、B3处理(P<0.05);B2、B1处理在90~150 d 地径生长量较大,分别增长了75.81%、55.14%,培育180 d后,B1处理地径几乎不再增长,而B3处理地径减少,部分枝条枯死,死亡率为3.39%。
以上表明,蓝莓‘莱克西在正常铁环境中正常生长,但在无铁环境以及难溶铁环境下生长受到抑制。另外,经3种不同铁素处理后蓝莓基生枝数量、基生枝生长量、地径均在培育90~150 d阶段表现出较强的生长势,各指标增长率较高,说明这个时间段是植物生长活力最旺盛的阶段。而在150~180 d期间各形态指标增长量逐渐减弱,甚至降低。
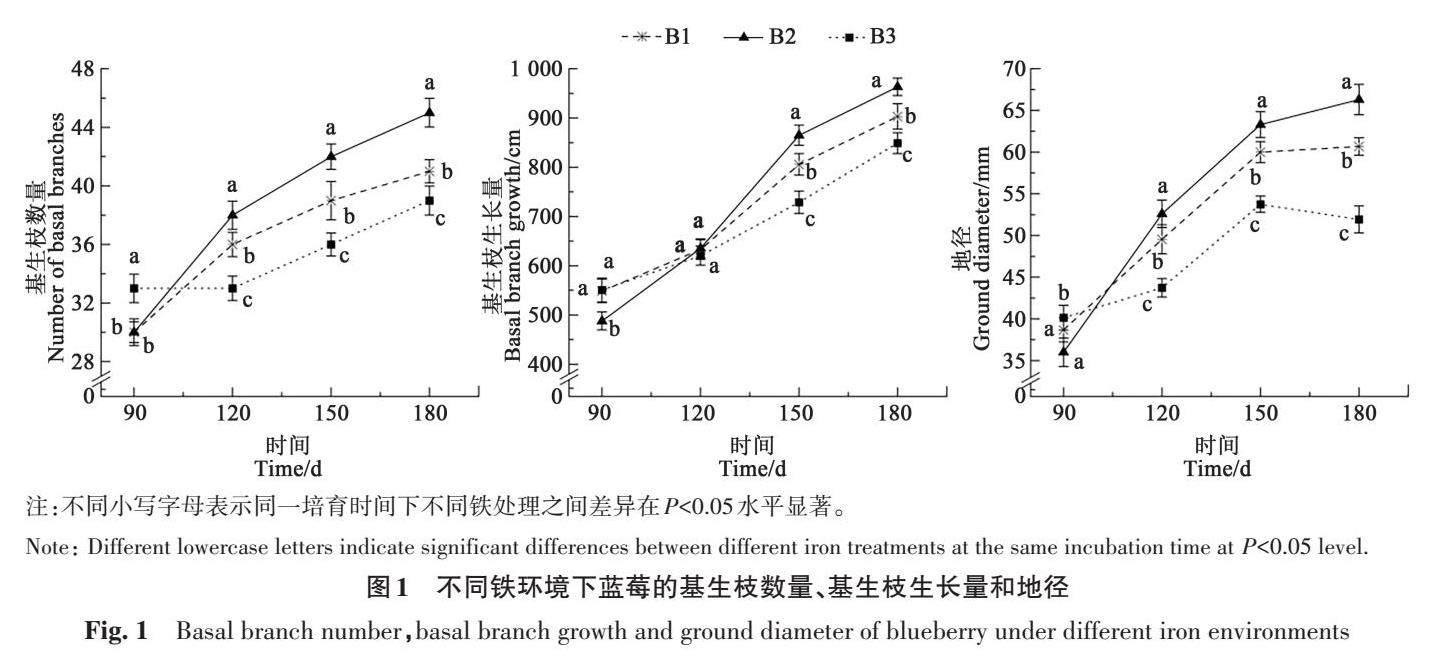
2.2 不同铁环境对蓝莓单株生物量、叶面积的影响
如图2所示,B1、B3处理单株生物量显著低于B2(P<0.05)。B1处理在第120~150 天生长势较第150~180 天弱,单株生物量整体增长了26.00%;B3处理单株生物量增长较B1更低,仅增长了16.80%,但二者间差异不显著。B2处理的叶面积在5.07~8.62 cm2,蓝莓平均叶面积随着处理时间的延长不断增加,增长量达53.77%;B3处理叶面积增长量较低,仅为11.26%;3种铁环境间蓝莓叶面积差异显著(P<0.05),B1、B3处理下蓝莓生长受胁迫,导致蓝莓叶面积变小。其中,B3处理产生的抑制效应更强,叶片受损伤程度较大,不利于蓝莓生物量的积累。
2.3 不同铁环境对蓝莓叶绿素及其相对含量(SPAD 值)的影响
随着处理时间的增加,B2处理叶片为绿色,B1、B3处理叶片表现出脉间叶色为黄色,叶片薄而透,叶缘卷曲,叶尖部分出现枯死。由表2可知,B2处理的SPAD值显著高于B1、B3处理(P<0.05),B3处理的SPAD值不断降低,在试验末期3个处理的SPAD值表现为:B3
2.4 不同铁环境对蓝莓叶绿素荧光参数的影响
Y(Ⅱ)是光照条件下PSⅡ的实际光合效率,能够间接反映线性电子传递的活性。由图3可知,蓝莓在B2处理环境下培育150 d后Y(Ⅱ)呈下降趋势,下降了39.10%;B1、B3处理的Y(Ⅱ)均低于B2,B1处理在60~120 d内实际光合效率急剧降低72.76%,120 d 后逐渐提高并高于B3 处理;培育150 d后B3处理PSⅡ电子传递活性低于B2、B1处理,其光合电子传递活性最低。
ETR 代表PSⅡ电子传递速率。B2 处理的PSⅡ电子传递速率最高(图3);B1 处理在培育60~120 d内降幅达到最大,为72.73%,培育150 d后又增加;B3处理在培育150 d后PSⅡ电子传递效率低于B2和B1处理。
NPQ是一种能量耗散机制,测量诱导型非光化学淬灭,反映植物的光保护能力。B2处理在整个培育期NPQ均最低(图3);B3处理培育90 d后NPQ 增加了51.81%;B1 处理在培育期间NPQ 呈先降低后升高的趋势,且均高于B2、B3处理,此时苗木的热耗散作用加强。与B2处理相比,B1、B3处理均使蓝莓生长受抑制,植物通过加强热耗散适应逆境胁迫。
Y(NO)是非调节性能量耗散的量子产率,反映了植物受光抑制的程度。B2处理非调节性能量耗散的量子产率在90 d后逐渐减弱并趋于稳定,且表现为最低(图3);B3处理的光抑制程度分别高于B2、B1处理15.88%、6.45%,在培育至90 d时B3光抑制程度达到峰值,90 d后Y(NO)不断降低直至稳定;B1处理受光抑制程度处于B2和B3处理之间,B3处理使蓝莓叶片受光损伤的程度更高。
1-qP表示了PSⅡ反应中心的关闭程度,反映植物逆境环境下光系统中心的活性。由图3知,B2处理的1-qP最低,表明蓝莓遭受胁迫程度更强。培育120 d后植物PSⅡ反应中心的活性依次为B2>B1>B3,B3处理使蓝莓PSⅡ反应中心关闭程度达到最大。
Fv/Fm是暗适应下PSⅡ反应中心完全开放时的最大光化学效率,反映了光系统中心转换效率的稳定性。B2处理最大光化学效率在0.73左右,其效率最稳定(图3);B1、B3处理在PSⅡ反应中心的Fv/Fm降低至0.5以下,B3处理蓝莓潜在的最大光合能力最弱,B1处理次之;3种不同铁素环境处理间PSⅡ最大光合效率具有差异。
以上结果表明,各荧光参数体现了蓝莓在缺铁环境和难溶铁环境下,蓝莓叶片光系统受损导致植物光合作用不能正常进行。结合3种铁处理后蓝莓生长发育特点,蓝莓叶片生长遭受B1、B3胁迫处理而受抑制,叶片色素合成受阻,光合电子传递速率减弱。
2.5 不同生长发育指标和各叶片光合能力参数间的相关性
对各指标参数进行Pearson相关性分析(表3)可知,叶面积与单株生物量、基生枝数量、Y(Ⅱ)、ETR、Fv/Fm均呈极显著正相关(P<0.01),平均叶面积与Y(NO)、1-qP呈极显著负相关(P<0.01);蓝莓单株生物量与基生枝数量、Y(Ⅱ)、ETR、Fv/Fm呈极显著正相关(P<0.01),与Y(NO)、1-qP 呈极显著负相关(P<0.01);叶绿素a与叶绿素b、总叶绿素、SPAD值间均呈极显著正相关(P<0.01),与Fv/Fm间具有正相关性,但不显著;不同叶绿素荧光参数间也具较大的相关性,其中Y(II)与ETR、Fv/Fm呈极显著正相关(P<0.01),与Y(NO)、1-qP呈极显著负相关(P<0.01);而Y(NO)和1-qP呈极显著正相关(P<0.01)。
3 讨论
3.1 不同铁环境对蓝莓苗木营养器官生长发育的作用
本研究设置了正常铁、难溶铁以及缺铁3种铁环境,结果表明,蓝莓在Na-Fe-EDTA提供的正常Fe2+环境中生长良好,在难溶Fe2O3、缺铁环境下蓝莓生物生长量较Na-Fe-EDTA低,基生枝数量、基生枝生长量、地径、叶面积、单株生物量随着胁迫时间的延长其增长量逐渐减小,该结果与Larbi等[25]和黄小辉等[26]的研究结果相似,当植物生长必需的营养缺失时植物生物量显著降低。研究结果验证了蓝莓主要吸收利用Fe2+ [12],也是铁吸收机理I植物的主要特征。在缺铁环境下,各生长指标表明蓝莓生长受抑制。当只提供难溶铁时,蓝莓无法正常吸收利用铁营养,该环境下蓝莓的叶面积较小,其叶片生长量进一步体现了难溶铁环境下蓝莓受胁迫程度较高,也有研究发现在低铁胁迫下不同品种玉米的叶面积变小[27]。叶面积减小间接影响植物光合反应效率,叶片接收的光能变少,又因铁营养不足导致叶片受损,光合能力减弱,即难溶铁或缺铁环境均会使植物遭受铁胁迫。陈娜[10]关于不同螯合铁铁素水平对蓝莓生长发育影响的研究中,铁浓度50 μmol·L-1时蓝莓生物量、叶绿素含量等表现最佳,进一步说明蓝莓适生于Na-Fe-EDTA提供的Fe2+环境中。
3.2 不同铁环境对蓝莓叶片光合能力的作用
叶片是植物进行光合作用的主要部位,是植物吸收、转换、传递光能的重要场所[2829]。而铁是参与叶绿素合成的主要微量元素,铁元素影响并作用于叶绿体,对叶绿体的结构和功能具有重要的维持作用[30]。牛永昆等[31]对蕨麻的研究表明,缺铁使SPAD值降低,大量铁则显著提高了SPAD值,充足的Fe2+环境有利于植物叶片叶绿素合成。本研究中不同铁环境对蓝莓叶绿素a、叶绿素b、总叶绿素及SPAD值的影响具有显著性差异(P<0.05),难溶Fe3+、缺铁环境下蓝莓叶片叶绿素a、叶绿素b、总叶绿素含量较正常铁低,和光合色素含量间具有显著正相关的SPAD值也降低,这与已有研究结果[17,31-34]相似。而高等植物的叶绿素合成是复杂的过程,其中任何环节受抑制都会影响叶绿素含量的变化[35]。难溶铁和缺铁环境可能使叶绿素合成的某一环节受抑制,导致光合色素含量降低,而关于这部分的内容有待进一步研究。Sun等[36]研究表明,叶绿素a对于金属胁迫较叶绿素b敏感,而叶绿素荧光主要来源于PSⅡ的叶绿素a,其中光反应和暗反应等多种光合过程的变化均反馈给PSⅡ,从而引起叶绿素a的荧光变化。
植物的光合作用能通过叶绿素荧光反映,进一步体现植物在不同环境条件下的生长状态或受胁迫抑制状况[37]。本试验中,蓝莓‘莱克西培育在温室内,其光照强度较大田更弱,各处理下苗木均生长于弱光环境中,导致PSⅡ反应中心最大光化学效率整体偏低,而本研究主要利用叶绿素荧光参数来体现蓝莓在正常铁、难溶铁、缺铁3种环境下的差异。Fv/Fm反映PSⅡ光化学效率的稳定性,本研究中正常铁环境下蓝莓的最大光化学效率维持在0.73左右,在难溶铁及缺铁环境下蓝莓叶片随着胁迫天数增加Fv/Fm 不断降低至0.5 以下,而最大光能转换效率越高植物光能转化为化学能的能力越强[38]。Y(Ⅱ)、ETR在2种铁胁迫环境中不断下降,说明叶片遭受损伤导致其吸收的光能无法有效地转换为化学能[31]。同时,与Fv/Fm、Y(Ⅱ)、ETR具有显著负相关的Y(NO)、NPQ、1-qP的变化更进一步说明了难溶铁以及缺铁环境使蓝莓遭受铁胁迫,该环境中叶片Y(NO)、NPQ、1-qP随胁迫时间延长显著高于正常铁环境,数值越大表示植物遭受的胁迫越强,叶片PSⅡ反应中心关闭的程度越高,蓝莓受光抑制的程度越大,植物会通过启动热耗散机制耗散过剩的激发能来保护光合器官免受损伤[3940]。而肖家欣等[41]研究发现,叶片光合能力在低铁处理下弱于缺铁,这与本研究结果具有相似性,可能蓝莓‘莱克西在利用铁营养能力方面较差。蓝莓叶面积、单株生物量等生长发育特征与叶绿素荧光参数显著相关,也体现了蓝莓在仅有难溶铁及缺铁环境下无法获得铁营养供给,各营养器官生长发育受到抑制。
参 考 文 献
[1] 李亚东,盖禹含,王芳,等. 2021年全球蓝莓产业数据报告 [J/OL].吉林农业大学学报,2022,44(1):1-12.
LI Y D, GAI Y H, WANG F, et al ..Global blueberry industrydata report 2021 [J/OL]. J. Jilin Agric. Univ., 2022,44(1):1-12.
[2] 尤式备,徐佳慧,郭怡文,等.蓝莓根毛缺失的机制及内生菌根真菌的促生作用[J].浙江大学学报(农业与生命科学版),2020,46(4):417-427.
YOU S B, XU J H, GUO Y W, et al ..Mechanism of root hairloss in blueberry and growth promoting effect of endophyticmycorrhizal fungi [J]. J. Zhejiang Univ. (Agric. Life Sci.), 2020,46(4):417-427.
[3] 孙贵宝.浅谈欧美及日本等国蓝莓栽培的立地条件[J].北方园艺,2006(4):103-105.
SUN G B.Study on the site conditions of blueberry cultivationin Europe, America and Japan [J].Northern Hortic., 2006(4):103-105.
[4] 李亚东,陈伟,张志东,等.土壤pH值对越桔幼苗生长及元素吸收的影响[J].吉林农业大学学报,1994,16(3):51-54.
LI Y D, CHEN W, ZHANG Z D, et al ..Effects of soil pH ongrowth and elemental absorption of bilberry seedlings [J]. J.Jilin Agric. Univ., 1994,16(3):51-54.
[5] 曹增强,徐莹莹,张宁,等.不同pH对蓝莓组培苗生长和元素吸收的影响[J].中国农业大学学报,2016,21(2):50-57.
CAO Z Q, XU Y Y, ZHANG N, et al ..Effects of different pHvalues on the growth and element absorption of blueberrytissue culture seedlings [J].J. Chin. Agric. Univ., 2016,21(2):50-57.
[6] COVILLE F V.Experiments in Blueberry Culture [M].Whitefish:Kessinger Publishing, 1911:1-138.
[7] 陈雅彬,李永强,孙琳,等.非酸性根际土壤对蓝莓铁元素吸收及其代谢相关基因表达的影响[J].园艺学报,2015,42(2):233-242.
CHEN Y B, LI Y Q, SUN L, et al .. Effects of non-acidicrhizosphere soil on iron uptake and metabolization-relatedgene expression in blueberry [J].Acta Hortic. Sin., 2015,42(2):233-242.
[8] 叶美娟.蓝莓缺铁响应机制研究[D].金华:浙江师范大学,2017.
YE M J. Research on the mechanism of iron deficiency inblueberry [D]. Jinhua: Zhejiang Normal University, 2017.
[9] 孙攀,杨静慧,冀馨宁,等.不同铁肥对蓝莓黄化叶片光合特性的影响[J].天津农林科技,2019(2):16-18.
SUN P, YANG J H, JI X N, et al .. Effects of different Ironfertilizer on photosynthetic characteristics of chlorinatedblueberry leaves [J]. Sci. Technol. Tianjin Agric. For., 2019(2):16-18.
[10] 陈娜.铁、钾营养对越橘苗生长和生理特性的影响[D].大连:大连理工大学,2010.
CHEN N. Effects of iron and potassium on growth andphysiological characteristics of Huckleberry seedlings [D].Dalian: Dalian University of Technology, 2010.
[11] EIDE D, BRODERIUS M, FETT J, et al .. A novel ironregulatedmetal transporter from plants identified by functionalexpression in yeast [J]. Proc. Natl. Acad Sci. USA,1996,93(11):5624-5628.
[12] GUERINOT M L, YI Y. Iron: nutritious, noxious, and notreadily available [J]. Plant Phys., 1994,104(3):815-820.
[13] TERRY N, ABAD?A J.Function of iron in chloroplasts [J]. J.Plant Nutr., 1986,9(3-7):609-646.
[14] ZHOU H J, KORCAK R F, WERGIN W P, et al .. Cellularultrastructure and net photosynthesis of appleseed lings,underiron stree [J]. J. Plant Nutr., 1984,7(6):911-928.
[15] 伍丽华,代林利,胡永颜,等.供铁水平对杉木幼苗叶片叶绿素荧光参数和抗氧化酶活性的影响[J].江西农业大学学报,2021,43(5):1087-1097.
WU L H, DAI L L, HU Y Y, et al ..Effects of iron supply onchlorophyll fluorescence parameters and antioxidant enzymeactivity of Cunninghamia lanceolata seedlings [J].Acta Agric.Univ. Jiangxiensis, 2021,43(5):1087-1097.
[16] BERTAMINI M, MUTHUCHELIAN K, NEDUNCHEZHIANN. Iron deficiency induced changes on the donor side of PSⅡin field grown grapevine (Vitis vinifera L. cv. Pinot noir) leaves [J].Plant Sci., 2002,162(4):599-605.
[17] 邱强,饶德民,赵婧,等.不同铁效率大豆品种叶片和根系超微结构的比较研究[J].大豆科学,2017,36(6):927-931.
QIU Q, RAO D M, ZHAO J, et al .. Comparison of leaf and rootultrastructure of soybean varieties with different iron efficiency [J].Soybean Sci., 2017,36(6):927-931.
[18] 郭燕萍,顾家家,陈冰聪,等.蓝莓铁代谢调控机制初探[J].浙江师范大学学报(自然科学版),2021,44(3):311-318.
GUO Y P, GU J J, CHEN B C, et al .. Research on theregulation mechanism of iron metabolism in blueberry [J]. J.Zhejiang Norm. Univ. (Nat. Sci.), 2021,44(3):311-318.
[19] 黄宗安,徐坚,史建磊, 等.缺铁胁迫对温州盘菜幼苗叶片光合特性和抗氧化酶活性的影响[J]. 浙江农业学报,2014,26(6):1495-1500.
HUANG Z A, XU J, SHI J L, et al .. Effects of iron deficiencystress on photosynthetic characteristics and antioxidantenzyme activities of cabbage seedlings [J]. Acta Agric.Zhejiangensis, 2014,26(6):1495-1500.
[20] 姜闯道,高辉远,邹琦. 缺铁使大豆叶片激发能的耗散增加[J].植物生理与分子生物学学报,2002(2):127-132.
JIANG C D, GAO H Y, ZOU Q.Effect of iron deficiency onexcitation energy dissipation in soybean leaves [J]. Physiol.Mol. Biol. Plants, 2002(2):127-132.
[21] 杨颖丽,司廉邦,李嘉敏,等.铁胁迫对小麦幼苗叶绿素荧光与光合特性的影响[J]. 兰州大学学报(自然科学版),2019,55(6):814-820.
YANG Y L, SI L B, LI J M, et al ..Effects of iron stress onchlorophyll fluorescence and photosynthetic characteristics ofwheat seedlings [J]. J. Lanzhou Univ. (Nat. Sci.), 2019, 55(6):814-820.
[22] ADAMSKI J M, PETERS J A, DANIELOSKI R, et al .. Excessiron-induced changes in the photosynthetic characteristics ofsweet potato [J]. J. Plant Physiol., 2011,168(17):2056-2062.
[23] 陈薇.温室、大棚、露地三种栽培模式下越橘光合特性研究[D].长春:吉林农业大学,2006.
CHEN W. Photosynthetic characteristics of huckleberry undergreenhouse, greenhouse and open field cultivation [D].Changchun: Jilin Agricultural University, 2006.
[24] LICHTENTHALER H. Chlorophylls and carotenoids: pigmentsof photosynthetic biomembranes [J]. Meth. Enzymol.,1987,148C:350-382.
[25] LARBI A, MORALES F, L?PEZ-MILL?N A F, et al ..Technical advance: reduction of Fe(Ⅲ)-chelates by mesophyllleaf diska of sugar beet.multi-component origin and effects Fedeficiency [J]. Plant cell Physiol., 2001, 42 (1):94-105.
[26] 黄小辉,吴焦焦,王玉书,等.不同供氮水平核桃的生长及叶绿素荧光特性[J/OL]. 南京林业大学学报(自然科学版),2022 [2022-05-12].https://kns.cnki.net/kcms/detail/32.1161.S.20220129.1658.002.htm.
HUANG X H, WU J J, WANG Y S, et al .. Growth andchlorophyll fluorescence characteristics of walnut withdifferent nitrogen supply levels [J/OL]. J. Nanjing For. Univ.(Nat. Sci.), 2022 [2022-05-12]. https://kns. cnki. net/kcms/detail/32.1161.S.20220129.1658.002.htm.
[27] 龙文靖,万年鑫,辜涛,等.玉米苗期耐低铁能力的综合评价及其预测[J].植物遗传资源学报,2015,16(4):734-742.
LONG W J, WAN N X, GU T, et al .. Comprehensive evaluationand prediction of low iron tolerance of maize seedling [J]. J.Plant Genet. Resour., 2015,16(4):734-742.
[28] 游丹丹,张子威,王贞红.西藏墨脱县4个茶树良种叶片表型及光合特性分析[J/OL]. 分子植物育种,2023,21(17):5794-5801.
YOU D D, ZHANG Z W, WANG Z H. Analysis of leafphenotype and photosynthetic characteristics of four teacultivars in Motuo county, Xizang province [J/OL]. Mol. PlantBreeding,2023,21(17):5794-5801..
[29] 厉广辉,张昆,刘风珍,等.不同抗旱性花生品种的叶片形态及生理特性[J].中国农业科学,2014,47(4):644-654.
LI G H, ZHANG K, LIU F Z, et al .. Leaf morphology andphysiological characteristics of peanut cultivars with differentdrought resistance [J]. Sci. Agric. Sin., 2014,47(4):644-654.
[30] TAGLIAVINI M, SCUDELLARI D, MARANGONI B, et al ..Acid-spray regreening of kiwifruit leaves affected by limeinduced Fe chlorosis [J]. Iron Nutr. Soils Plants,1995:191-195.
[31] 牛永昆,李军乔,曲俊儒,等.铁胁迫对蕨麻叶绿素荧光及生理特性的影响[J].天津农业科学,2022,28(2):1-6.
NIU Y K, LI J Q, QU J R, et al .. Effects of iron stress onchlorophyll fluorescence and physiological characteristics ofpotentilla anserine [J]. Tianjin Agric. Sci., 2022,28(2):1-6.
[32] 姚宇洁,姜存仓.缺铁胁迫柑橘砧木幼苗的光合特性和叶绿体超微结构 [J].植物营养与肥料学报,2017,23(5):1345-1351.
YAO Y J, JIANG C C. Photosynthetic characteristics andchloroplast ultrastructure of citrus rootstock seedlings underiron deficiency stress [J]. J. Plant Nutr. Fert., 2017,23(5):1345-1351.
[33] 王凯,吴祥云,段海侠,等.辽西北主要绿化树种叶绿素含量分异特征及与SPAD 值关系[J]. 植物研究, 2014, 34(5):634-641.
WANG K, WU X Y, DUAN H X, et al .. Characteristics ofchlorophyll content and its relationship with SPAD value ofmain tree species in Northwest Liaoning province [J]. Bull. Bot.Res., 2014,34(5):634-641.
[34] MOLASSIOTIS A, TANOU G, DIAMANTIDIS G, et al .. Effectsof 4-month Fe deficiency exposure on Fe reduction mechanism,photosynthetic gas exchange, chlorophyll fluorescence andantioxidant defense in two peach rootstocks differing in Fedeficiency tolerance [J]. J. Plant Physiol., 2006, 163(2):176-185.
[35] 李佳佳,于旭东,蔡泽坪,等.高等植物叶绿素生物合成研究进展[J].分子植物育种,2019,17(18):6013-6019.
LI J J, YU X D, CAI Z P, et al .. Research progress ofchlorophyll biosynthesis in higher plants [J]. Mol. Plant Breed.,2019,17(18):6013-6019.
[36] SUN X L, XU Y, ZHANG Q Q, et al .. Combined effect of waterinundation and heavy metals on the photosynthesis andphysiology of Spartina alterniflora [J]. Ecotox. Environ. Safe., 2018,153(5):248-258.
[37] MORANT-MANCEAU A, PRADIER E, TREMBLIN G.Osmotic adjustment, gas exchanges and chlorophy Ⅱfluorescence of a hexaploid triticale and its parental speciesunder salt stress [J]. J. Plant Physiol., 2004,161(1):25-33.
[38] 闫超.不同蓝莓品种光合特性初步研究[D].哈尔滨:东北农业大学,2016.
YAN C.Preliminary study on photosynthetic characteristics ofdifferent blueberry cultivars [D]. Harbin: Northeast AgriculturalUniversity, 2016.
[39] DEMMIG-ADAMS B, ADAMS W W, HEBER U, et al ..Inhibition of zeaxanthin formation and of rapid changes inradiationless energy dissipation by dithiothreitol in spinachand chloroplast [J]. Plant Physiol., 1990,92(2):293-301.
[40] 彭海欢,翁晓燕,徐红霞,等.缺钾胁迫对水稻光合特性及光合防御机制的影响[J].中国水稻科学,2006,20(6):621-625.
PENG H H, WENG X Y, XU H X, et al .. Effects of potassiumdeficiency stress on photosynthetic characteristics andphotosynthetic defense mechanism of rice [J]. Chin. J. RiceSci., 2006,20(6):621-625.
[41] 肖家欣, 齐笑笑, 张绍铃. 铁胁迫对三种柑橘砧木的生长、生理特性及铁分布的影响[J]. 广西植物, 2011, 31(1): 97-101.
XIAO J X, QI X X, ZHANG S L. Effects of Iron stress ongrowth, physiological characteristics and iron distribution ofthree citrus rootstocks [J]. J. Guihaia, 2011, 31(1): 97-101.
(责任编辑:胡立霞)
猜你喜欢
河北果树(2021年4期)2021-12-02
音乐教育与创作(2020年2期)2020-06-19
商周刊(2017年5期)2017-08-22
作文周刊·小学一年级版(2017年26期)2017-08-10
山东农业科学(2017年2期)2017-03-15
农家科技下旬刊(2016年9期)2016-12-15
现代园艺(2016年2期)2016-03-15
江苏农业科学(2015年11期)2016-01-27
江苏农业科学(2015年8期)2015-09-10
