
外源氯化钙对大蒜幼苗盐胁迫伤害的缓解作用
2017-02-15 20:17朱利君闫秋洁
江苏农业科学 2016年8期
朱利君+闫秋洁


摘要:使用不同浓度的氯化钙对盐胁迫下大蒜(Allium sativum L.)幼苗进行处理,对大蒜幼苗的株高、根长、超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、过氧化氢酶(CAT)活性、抗坏血酸过氧化物酶(APX)活性和叶绿素、脯氨酸、丙二醛(MDA)、H2O2含量进行了测定。结果表明,150 mmol/L NaCl胁迫显著抑制了大蒜幼苗的生长,添加不同浓度的CaCl2后,大蒜幼苗的株高、根长、叶绿素含量、抗氧化保护酶活性逐渐增加,脯氨酸、MDA、HO2含量逐渐降低。当在150 mmol/L NaCl中添加30 mmol/L CaCl2后,大蒜幼苗的株高、根长、叶绿素含量、抗氧化保护酶活性达到最大,脯氨酸、MDA、H2O2含量降到最低。因此,外源CaCl2可以缓解NaCl胁迫对大蒜幼苗的伤害,最适用浓度为 30 mmol/L CaCl2。
关键词:外源钙;盐胁迫;大蒜;幼苗;生理特性;抗氧化酶
中图分类号: S633.401文献标志码:
文章编号:1002-1302(2016)08-0242-03
大蒜(Allium sativum L.),多年生草本植物,百合科葱属。现代科学分析证明,大蒜含有蛋白质,味道香辛,因具有丰富的营养保健和药用价值[1]而成为近几年研究的热点。土壤盐渍化是农作物生长中常见的自然逆境之一。中国是世界上盐碱土大国之一。盐胁迫是影响植物生长发育和农作物产量的主要环境胁迫因子之一,中国盐荒地约有2 000万hm2,次生盐渍化土壤670万hm2,约占全国总土地面积的14%。研究大蒜抗盐生理机制,对减少土壤盐渍化的危害、提高大蒜产量具有重要意义[2]。
植物在受到逆境胁迫时能提高游离Ca2+的浓度,并通过Ca2+与钙调蛋白结合启动一系列生理生化过程,对逆境的感受、传递、响应和适应过程中发挥重要作用。钙能提高植物组织或细胞的多种抗性和抗多种矿质元素毒害胁迫等。适量的钙离子能降低质膜透性,阻止胞内钾离子的外渗和钠离子的进入,从而提高植物的耐盐性,促进植物的生长[3]。有关钙对盐胁迫影响的报道,多集中在大田作物,如小麦、玉米、大豆[4-6]等。外源钙能否缓解盐胁迫对大蒜幼苗的伤害,到目前为止还未见报道。本试验采用不同浓度外源钙作用于盐胁迫下的大蒜幼苗,测定大蒜幼苗的株高、根长,抗氧化保护酶SOD、POD、CAT、APX活性,脯氨酸、MDA、叶绿素、H2O2含量,筛选出缓解大蒜幼苗盐胁迫的最适外源钙浓度,以期为大蒜在生产种植提供相关参考依据。
1材料与方法
1.1材料
选取籽粒饱满健康、大小一致的大蒜鳞茎,漂洗干净,用10%次氯酸钠溶液消毒10 min,用蒸馏水冲洗4次,于25 ℃恒温水浴锅中浸种24 h,置于铺有3层纱布的培养皿中,分别用0(对照)、50、100、150、200 mmol/L NaCl 溶液处理,每处理100粒,3次重复。22 ℃恒温箱催芽,第5天统计其芽长、根长。以芽长超过种子长度的50%为发芽标准[3]。待萌发后,选取未处理的大蒜分别用0、150 mmol/L NaCl、150 mmol/L NaCl+15 mmol/L CaCl2、150 mmol/L NaCl+20 mmol/L CaCl2、150 mmol/L NaCl+30 mmol/L CaCl2、150 mmol/L NaCl+40 mmol/L CaCl2、150 mmol/L NaCl+50 mmol/L CaCl2分别处理,每个处理加入1/2 Hoagland营养液。每处理30株,3次重复。每天更换1次处理液,处理 10 d 后,检测大蒜苗的株高和根长,同时检测其SOD、POD、CAT、APX活性,H2O2、叶绿素、脯氨酸、MDA含量。
1.2测定方法
SOD活性测定参照Stewart等的方法[7]。 APX活性测定参照Dong等的方法[8]。POD活性、CAT活性和H2O2、叶绿素、脯氨酸、MDA含量测定参照张志良等的方法[9]。
1.3数据处理
数据用SPSS 17.0统计软件处理,用One-way ANOVA进行单因素方差分析,若影响显著,再用Duncan法对平均值进行多重比较。
2结果与分析
2.1不同浓度的NaCl胁迫对大蒜幼苗芽长和根长的影响
不同浓度NaCl胁迫处理的大蒜,随着NaCl浓度的升高,大蒜幼苗的芽长和根长逐渐降低。
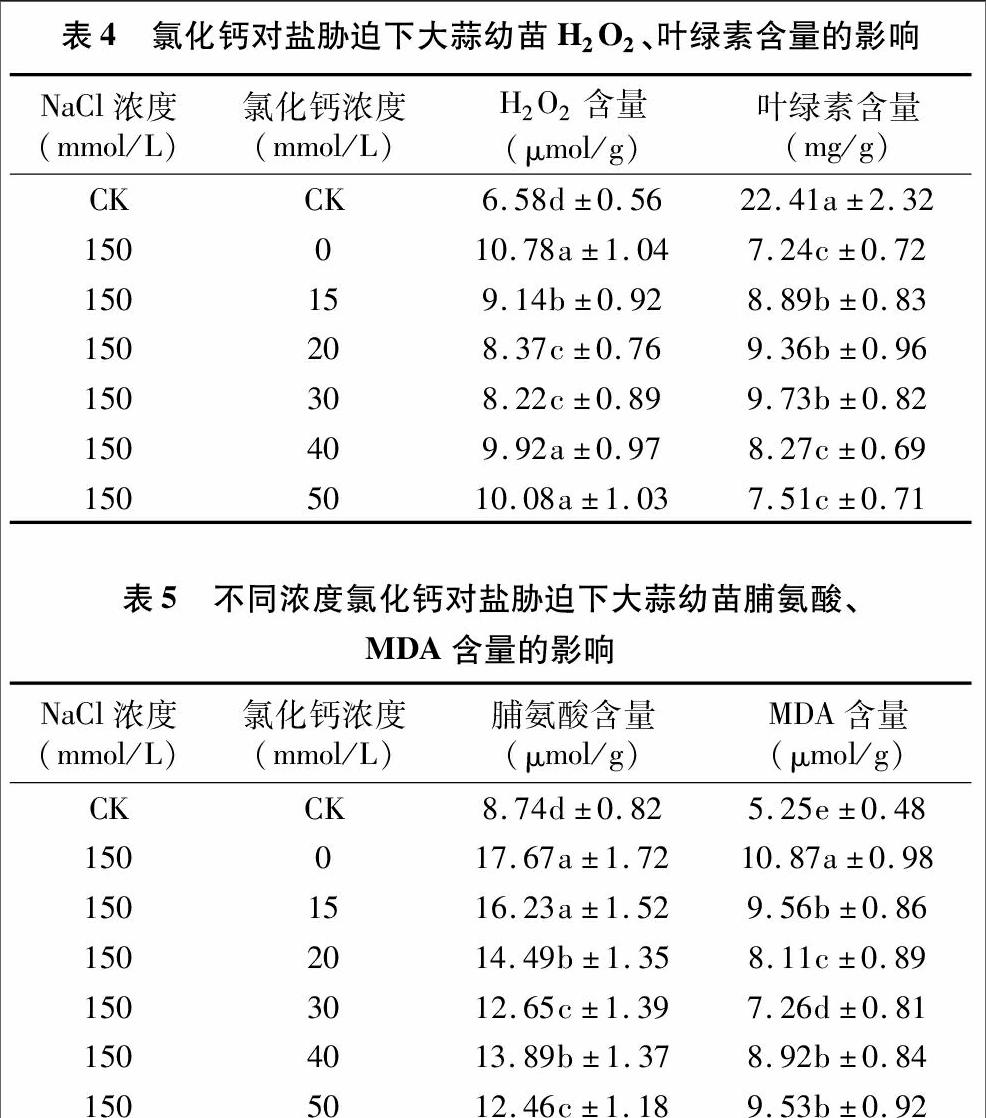
2.4不同浓度氯化钙对盐胁迫下大蒜幼苗的H2O2、叶绿素含量的影响
150 mmol/L NaCl处理大蒜幼苗的H2O2含量升高,叶绿素含量降低,与对照相比差异显著(表4)。添加不同浓度CaCl2后,大蒜幼苗的H2O2含量呈先降后升的变化趋势,叶绿素含量呈先升后降的变化趋势。当CaCl2浓度为 30 mmol/L 时,大蒜幼苗的H2O2含量降到最低,与单一盐胁迫处理相比,下降了23.75%,差异显著;而叶绿素含量达到最高,与单一盐胁迫处理相比,增加了25.59%,差异显著,表明CaCl2可以有效提高NaCl胁迫大蒜幼苗的叶绿素含量,缓解盐胁迫对幼苗光系统的损伤。
2.5不同浓度氯化钙对盐胁迫下大蒜幼苗脯氨酸、MDA含量的影响
150 mmol/L NaCl处理大蒜幼苗的脯氨酸、MDA含量均显著升高,與对照相比显著差异(表5)。添加不同浓度CaCl2后,大蒜幼苗的脯氨酸、MDA含量呈先降后升的变化趋势。当CaCl2浓度为30 mmol/L时,大蒜幼苗的脯氨酸、MDA含量降[CM(25]到最低,与单独盐胁迫处理相比,分别下降了2841%、[CM)]
3讨论
在150 mmol/L NaCl胁迫下,大蒜幼苗的生长受到抑制,当添加CaCl2后,大蒜幼苗的耐盐性明显增强。在添加不同浓度的CaCl2后,与单一盐胁迫相比,大蒜幼苗株高、根长均有所增加,幼苗的叶绿素含量和抗氧化酶活性增加、脯氨酸和MDA含量下降。当添加CaCl2 为30 mmol/L 时,大蒜幼苗的株高、根长、叶绿素含量、抗氧化酶活性达到最大,脯氨酸、MDA含量降到最低。因此,缓解NaCl胁迫对大蒜幼苗伤害的最适CaCl2浓度为30 mmol/L。
盐胁迫主要包括离子不平衡、渗透胁迫、离子毒害和营养缺乏等[10]。盐胁迫下植物生长受到抑制,能耗增加,光合下降,胞内物质外渗,造成植物代谢状况恶化,植物体启动自身的CAT、POD等保护酶系统以抵御活性氧自由基对植物细胞造成的伤害[11]。本试验中150 mmol/L NaCl胁迫严重抑制了大蒜幼苗的生长,抗氧化保护酶SOD、POD、CAT、APX活性显著提高,当添加不同浓度的CaCl2 后,抗氧化保护酶的活性进一步提高,来缓解NaCl胁迫对大蒜幼苗生长的影响。当添加CaCl2 为30 mmol/L时,抗氧化保护酶的活性达到最大。
植物体内叶绿素含量的多少,很大程度上决定光合作用的强弱,并与植物生长和生理状况紧密相关,是衡量植株代谢的一个重要指标[12]。本试验150 mmol/L NaCl胁迫处理使大蒜叶片叶绿素含量下降,添加不同浓度的CaCl2后,大蒜叶片中叶绿素含量显著提高,当CaCl2为30 mmol/L时,叶绿素含量达到最大。本结论与米银法等研究外源钙对盐胁迫下菊花生理效应的影响结论[3]一致。可能Ca2+有利于叶绿体膜结构的稳定性,进而提高叶绿素含量。
一般情况下,植物体内游离脯氨酸含量很低,但在逆境条件下脯氨酸含量可能增数十倍至百倍,因此,植物体内脯氨酸含量已成为抗逆性的一项生理指标[13]。本试验氯化钠胁迫下大蒜叶片中脯氨酸含量大量积累,添加氯化钙后脯氨酸的含量显著低于单一盐胁迫处理。当CaCl2为30 mmol/L时,脯氨酸含量降到最低,这与程玉静等研究外源硝酸钙对黄瓜幼苗盐胁迫伤害的缓解作用的结论[14]一致。可能是鈣离子改善了植株的水分状况,缓解渗透胁迫,使脯氨酸代谢达到平衡。
植物器官在逆境或衰老时,往往发生膜脂过氧化作用,丙二醛是其产物之一,其含量愈高过氧化作用愈强[15]。本试验中外源氯化钙处理显著降低了盐胁迫下大蒜幼苗的MDA含量,与曾长立等研究外源钙对盐胁迫下芸薹属植物幼苗的生理效应的结论[15]一致。可能是Ca2+降低了质膜透性,从而减轻对植物的伤害。
参考文献:
[1]梅四卫,朱涵珍. 大蒜研究进展[J]. 中国农学通报,2009,25(8):154-158.
[2]刘旭昊,王颖,谭梁静. 多效唑对盐胁迫下大蒜幼苗生长的影响[J]. 贵州农业科学,2010,38(9):42-44.
[3]米银法,王进涛,朱坤. 外源钙对盐胁迫下菊花生理效应影响[J]. 科技导报,2010,28(17):83-86.
[4]赵可夫,卢元芳,衣建龙,等. 钙对小麦幼苗降低盐害效应的影响[J]. 植物学报,1993,35(1):51-56.
[5]刘雪琴,仝瑞建,施佳妮. 外源钙对盐胁迫下玉米萌发与幼苗生长的影响[J]. 中国农学通报,2010,26(17):197-200.
[6]宋晓艳,安君. 盐胁迫对野大豆种子萌发特性的影响[J]. 内蒙古农业科技,2008(1):41-43.
[7]Stewart R R,Bewley J D. Lipid peroxidation associated with accelerated aging of soybean axes[J]. Plant Physiology,1980,65(2):245-248.
[8]Dong C J,Li L,Shang Q M,et al. Endogenous salicylic acid accumulation is required for chilling tolerance in cucumber (Cucumis sativus L.) seedlings[J]. Planta,2014,240(4):687-700.
[9]张志良,瞿伟菁,李小方. 植物生理学实验指导[M]. 4版. 北京:高等教育出版社,2009.[ZK)]
[10]朱义,何池全,杜玮,等. 盐胁迫下外源钙对高羊茅种子萌发和幼苗离子分布的影响[J]. 农业工程学报,2007,23(11):133-137.
[11]朱晓军,杨劲松,梁永超,等. 盐胁迫下钙对水稻幼苗光合作用及相关生理特性的影响[J]. 中国农业科学,2004,37(10):1497-1503.
[12]Ashraf M,Foolad M R. Roles of glycin ebetaine and proline in improving plant abiotic tress esistance[J]. Environmental and Experimental Botany,2007,59:206-216.
[13]张燕,方力,李天飞,等. 钙对低温胁迫的烟草幼苗某些酶活的影响[J]. 植物学通报,2002,19(3):342-347.
[14]程玉静,郭世荣,张润花,等. 外源硝酸钙对黄瓜幼苗盐胁迫伤害的缓解作用[J]. 西北植物学报,2009,29(9):1853-1859.
[15]曾长立,董元火. 外源钙对盐胁迫下芸薹属植物幼苗的生理效应[J]. 中国油料作物学报,2008,30(4):433-437,442.
猜你喜欢
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年12期)2017-01-21
现代园艺(2016年7期)2017-01-09
现代园艺(2016年9期)2016-07-16
现代园艺(2016年2期)2016-03-15
河北渔业(2015年10期)2015-10-15
