
野生藏酋猴理毛行为与肠道寄生虫感染的关系
2024-02-26 10:36张洁陈仕望田红卫孙丙华王希夏东坡
兽类学报 2024年1期
张洁 陈仕望 田红卫 孙丙华 王希 夏东坡*
(1 安徽大学生命科学学院,合肥 230601)(2 黄山生物多样性与短尾猴行为生态学国际联合研究中心,合肥 230601)(3 安徽大学资源与环境工程学院,合肥 230601)
理毛行为 (grooming) 指动物个体为自身或其他个体清理毛发的过程 (Tanaka and Takefushi,1993; Böröczkyet al., 2013; Zhukovskayaet al.,2013)。理毛行为是社会性动物中普遍存在的行为模式,例如有蹄类 (Mooringet al., 1996; Hart,2000; Heitoret al., 2006)、啮齿类 (Ferron and Lefebvre, 1982)、蝙蝠等 (Wilkinson, 1986)。在非人灵长类动物社会群体内,理毛行为是最为普遍并且发生最为频繁的社会行为模式 (Schino, 2007; Schino and Aureli, 2008),主要包括动物个体为自身进行的非社会性理毛 (auto-grooming) 和发生于两两个体之间的社会理毛 (social grooming) (Sachs,1998)。相比较仅具有物理清洁功能的自我理毛,社会理毛还具有重要的社会功能,例如促进个体之间社会联系的加强和群体的稳定等 (Dunbar,1991, 2010; Spruijtet al., 1992; Xiaet al., 2019)。
依据理毛行为参与者的个体角色,参与社会理毛的有两个动物个体,一个是理毛行为的发起者 (理毛者,Groomer),一个是理毛行为的接受者(被理毛者,Groomee)。对被理毛者而言,理毛行为可以为其清理毛发、降低心率、减轻压力 (Keverneet al., 1989; Feh and Mazières, 1993),理毛者也可以通过为其他个体投资理毛行为获得相应益处,例如缓和冲突、加强联系、提升交配机会等(Gumert, 2007; Xiaet al., 2012, 2013; Jianget al.,2019)。依据理毛行为的过程特征,有的理毛行为是理毛者使用双手梳理毛发,并不时地用手指捏住被理毛者毛发上的东西放入嘴里的过程,有的理毛行为是理毛者直接用嘴或舌头舔舐被理毛者身体表面 (羽毛、皮肤、毛发等) 的过程 (李进华,1999;李保国等,2002)。
进一步研究发现,理毛者在获益的同时也面临通过口感染病毒、细菌、病原体或寄生虫的风险(Fromontet al., 1997; Caillaudet al., 2006; Craftet al., 2008; Dreweet al., 2011; Rushmoreet al., 2013,2017)。如对西地大猩猩 (Gorilla gorilla) 的研究发现,参与理毛行为较多的个体感染埃博拉病毒的风险较高 (Caillaudet al., 2006);对猫鼬 (Suricata suricate) 的研究发现,为其他个体理毛较多的个体更容易感染结核杆菌 (Drewe, 2010)。不仅如此,理毛行为也会增加寄生虫感染的概率,如棕色蜘蛛猴(Ateles hybridus) (Rimbachet al., 2015)、藏酋猴(Macaca thibetana) (顾志远,2017)。可见,尽管传统观点认为理毛行为是一种低投资的行为模式(Pérez, 1998),但寄生虫传播的风险可能是理毛者面临的重要代价之一。然而,理毛行为感染何种类型寄生虫及其感染风险如何等这些问题仍需进一步研究。
本研究以栖息于安徽黄山的野生藏酋猴为对象,采集动物个体的粪便样品并检测其中寄生虫的种类,通过分析理毛行为与寄生虫感染的关系,探讨理毛者在理毛投资过程中的代价与风险,也为深入理解社会性动物构建友好关系的投资与收益提供重要科学依据。
1 研究方法
1.1 研究地点和对象
野外研究的地点位于安徽省黄山市的浮溪野生猴谷 (北纬30°29′,东经118°10′)。研究对象为长期跟踪研究的鱼鳞坑A1 群 (YA1 群),群体成员均能通过个体特征 (毛发颜色、手指是否残缺、手指的状态以及眼睛、鼻子、耳朵是否有残缺等) 进行精准识别并系统命名 (李进华,1999)。研究期间,本群共有27 只成年个体,即本研究的观察取样对象 (表1)。
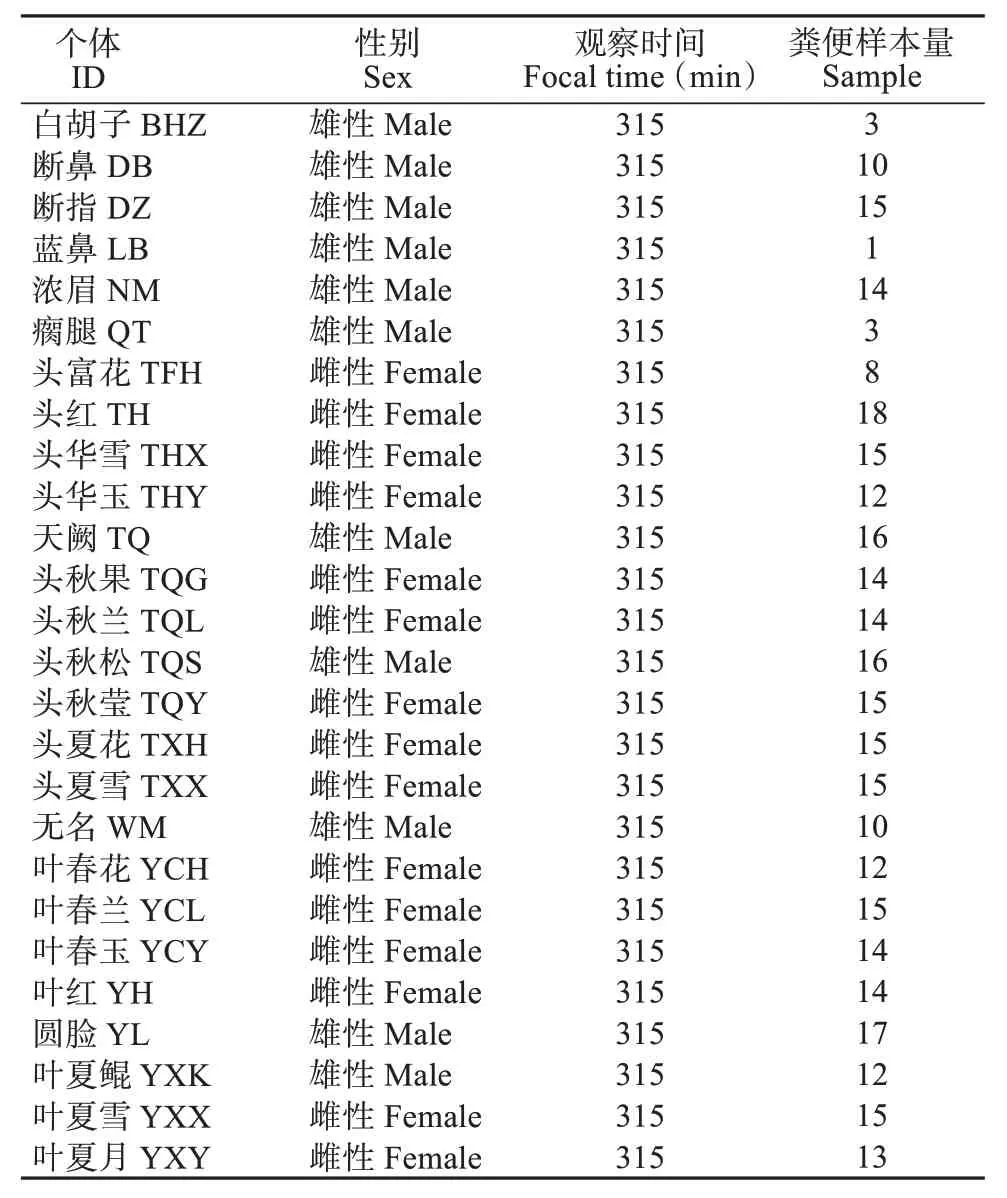
表1 本研究个体与取样基本信息Table1 Sample information of study individuals
1.2 行为数据采集
本研究野外观察时间为2022 年8 月至2023 年4 月,包括采集理毛行为数据和对应个体的粪便样品。受野生动物活动的不可控制性影响,2022 年8 月和12 月及2023 年2—4 月 (共计5 个月) 的理毛行为数据和对应个体的粪便样品采集匹配度较高,用于分析的主要是这期间的数据。
本研究采用焦点动物取样方法 (focal-animal sampling) 和连续记录法 (Continuous recording method) 记录行为数据 (Altmann, 1974),使用事件记录仪和录音笔等并辅以笔录,实时记录动物个体之间的理毛行为。理毛行为定义为一个个体为另一个个体清理毛发的过程 (Tanaka and Takefushi,1993; Böröczkyet al., 2013; Zhukovskayaet al,2013)。动物个体理毛5 s及以上被记录为有效理毛行为;如果理毛行为停止或被打断超过30 s或理毛者转换理毛对象,则被记录为新的理毛行为 (Bermanet al., 2008);如果3 个或3 个以上个体簇拥理毛,则按照两两关系分别记录理毛关系 (Kutsukake and Clutton-Brock, 2010);观察和记录理毛行为发生的环境,并记录每个理毛回合中理毛的发起者和接受者、理毛位点、起止时间等 (夏东坡,2013)。
1.3 粪便样本的采集和保存
研究期间,共采集新鲜粪便样本328份。当观察到猴群中有个体排便时,记录个体名字和排便时间,当该个体离开排便地点后,迅速用一次性无菌手套收集新鲜粪便样品,遗弃沾染尿液或其他液体污染的样本,去除石块泥土等杂物后,装入50 mL的粪便收集瓶中,并在瓶子上标记相应的信息,如动物个体名字、粪便样本采集时间等。加10%福尔马林溶液后,于3 h 内放置于4 ℃冰箱中保存 (用于测定寄生虫种类和负荷)。
1.4 肠道寄生虫的检测
参照Makiya (2001) 的检测方法,本研究采用铜丝筛和尼龙筛过滤离心沉淀法检测野生藏酋猴粪样寄生虫,具体步骤如下:(1) 过滤:称取福尔马林中保存的藏酋猴粪便1g,先经过80 目的铜丝筛,再通过160 目的尼龙筛进行过滤,倒去粪渣,收集滤液于500 mL 锥形量杯中,加清水至250 mL刻度处,静置2 h。(2) 离心沉淀淘洗:倒出上清液,将剩余沉淀混匀后转移至50 mL 离心管,加水至50 mL 刻度处,1 500 r/min 离心2 min。倒出上清,重复离心清洗2 ~ 3 次至上清液完全清澈。(3) 定量镜检:将离心沉淀转移至10 mL 离心管,并加清水至5 mL,混匀后吸取0.5 mL 沉淀在显微镜下镜检。(4) 虫卵鉴定计数:对镜检下所观察到的全部蠕虫虫卵及原虫囊蚴进行鉴定并计数。计数的每种虫卵或囊蚴数 × 20 即为每克虫卵数。本研究中寄生虫检测委托安徽省寄生虫防治研究所完成。
1.5 社会理毛网络构建
基于两两个体之间的理毛行为数据,本研究构建了理毛行为的加权网络,该网络考虑了接触网络中的社交互动频率和邻近网络中的个体间关联指数。在理毛网络中,用于链接的权重表示一对个体进行理毛的时间。通过将“发起”和“接收”理毛的时间加和,将权重应用于理毛网络。使用每个二元的关联指数对邻近网络中的链接进行加权。使用半权重指数 (HWI) (Whitehead,2008),公式如下:
其中mab表示个体a 和个体b 一起出现的时间;m0表示在采样过程中a 和b 都没有出现的时间;ma和mb表示在采样过程中只观察到a 和b 其中的一个(Whitehead and Dufault, 1999)。HWI 的范围从0 到1。0代表从未出现在一起的组合,1代表一直频繁出现在一起的组合。
基于理毛网络结构特征,计算每个个体在网络中的节点强度 (Carteret al., 2013),用于衡量每个动物个体在网络中的重要性。节点强度的值越大,表明个体在网络中的中心位置越高,其在病原体传播中的潜在作用就越大。
1.6 数据处理
所有数据先在Excel 2010 中进行初步汇总和整理,分别筛选动物个体的理毛时间 (包括发起理毛的时间和接收理毛的时间)。发起理毛的时间是指动物个体对其他个体的理毛总时间占目标动物取样总时间的百分比。接收理毛的时间是指其他个体对目标个体的理毛总时间占目标动物取样总时间的百分比。使用uncinet 软件构建个体之间的社会理毛网,从社会理毛网中提取参数 (节点强度)。同时,计算每个个体感染寄生虫的负荷量用于进一步分析。使用Spearman Correlation Analysis 分析理毛时间 (发起理毛的时间、接收理毛的时间) 与动物个体肠道寄生虫感染的相关性,使用Spearman Correlation Analysis 分析动物个体节点强度与肠道寄生虫感染的相关性。所有检验的显著性水平设置为α = 0.05 (双尾检验)。
2 结果
2.1 肠道寄生虫的种类和感染率
在野生藏酋猴粪便样品中共检测到3 类寄生虫,主要包括8种蠕虫类肠道寄生虫:鞭虫 (Trichuris trichura)、蛲虫 (Enterobius vermicularis)、钩虫 (Ancylostomasp.)、猴结节线虫 (Oesophagostomum apiostomum)、线虫 (Capillariaspp.,Trichostrongylusspp.)、粪类圆线虫 (Strongyloides stercoralis)、蛔虫 (Ascarisspp.)、食口线虫 (Oesophagostomumspp.),2 种原虫类肠道寄生虫:球虫(Coccidiasp.)、结肠小袋纤毛虫 (Balantidium coli),及1 种吸虫类肠道寄生虫:后睾吸虫 (Opisthorchis viverrini)。其中,线虫和结肠小袋纤毛虫感染率最高,均为93%;鞭虫和蛔虫次之,感染率分别为54%和50%。后睾吸虫、球虫和食口线虫感染率最低,为4% (表2)。
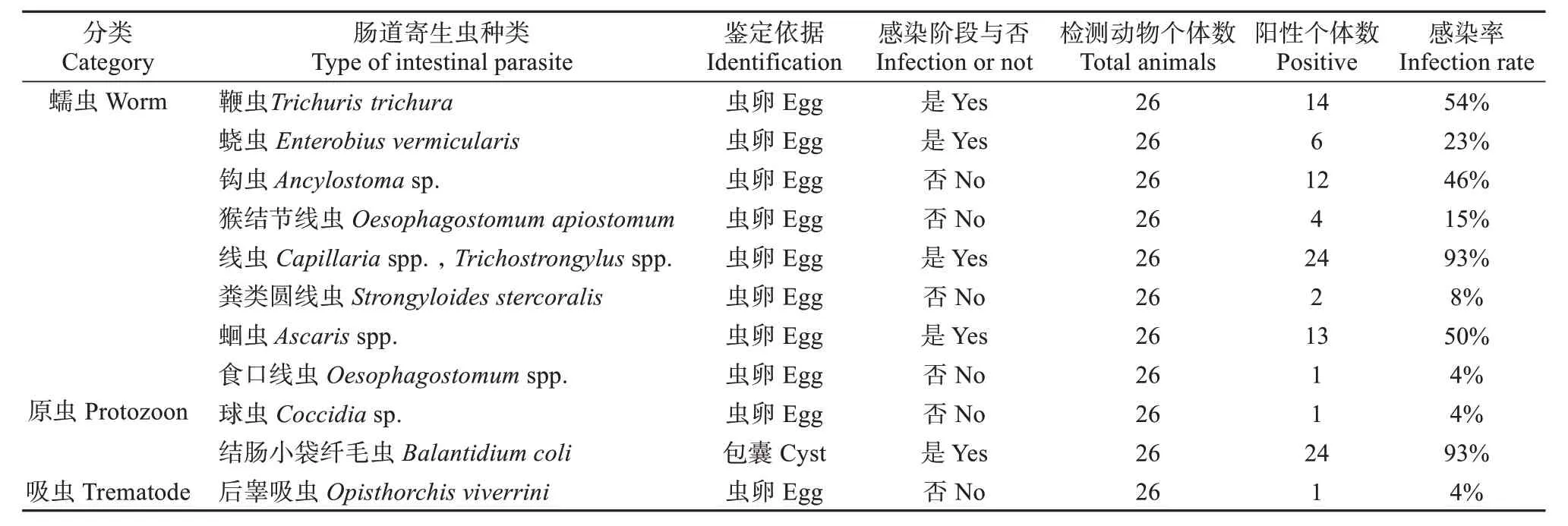
表2 藏酋猴肠道寄生虫感染的种类与感染率Table 2 Types and infection rates of intestinal parasites in Tibetan macaques
2.2 两个体间理毛行为与肠道寄生虫感染的关系
研究发现,野生藏酋猴发起理毛的时间越长,感染结肠小袋纤毛虫的概率越大 (r= 0.223,P=0.015);接收理毛的时间越长,感染蛔虫的概率越低 (r= -0.261,P= 0.004)。其他种类的肠道寄生虫 (鞭虫、钩虫、猴结节线虫、粪类圆线虫等)感染率与动物个体的理毛时间无显著相关 (P>0.05,表3)。
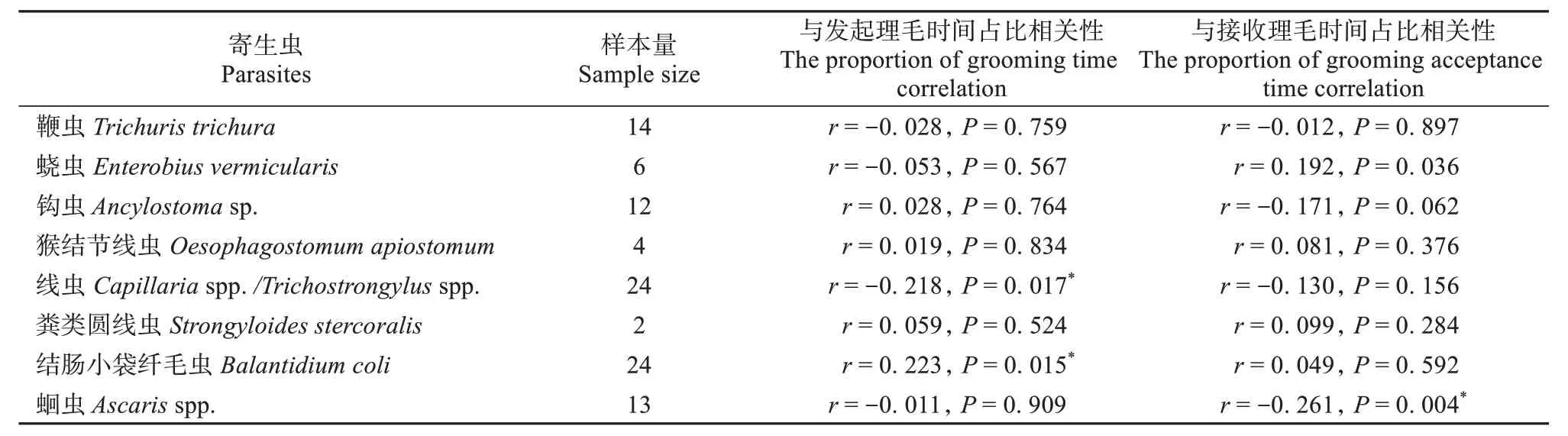
表3 藏酋猴理毛时间占比和理毛接受时间占比与肠道寄生虫的相关性Table 3 The correlation between the proportion of grooming time and the proportion of grooming acceptance time and intestinal parasites of Tibetan macaques
2.3 群体水平理毛行为对动物个体肠道寄生虫感染的影响
基于两个体间的理毛行为构建社会网络 (图1)。研究发现,动物个体在理毛网络中的节点强度与鞭虫、蛲虫、钩虫、猴结节线虫、线虫和粪类圆线虫等肠道寄生虫感染负荷之间无显著相关性 (P>0.05)。与两两个体之间理毛行为和肠道寄生虫感染的关系相似,群体水平上动物个体在理毛网络中的节点强度越大,其感染结肠小袋纤毛虫的概率越高 (r= 0.521,P= 0.006)。然而,动物个体在理毛网络中的节点强度与蛔虫感染负荷之间无显著负相关关系 (P> 0.05,表4)。

图1 藏酋猴社会理毛网络图. 节点表示群体中的所有成年个体;节点越大,节点强度越高;线条粗细表示成员在社交网络中的关联指数;雄性和雌性分别用蓝色方块和红色方块表示;标签上的字母代表每个个体的名字Fig. 1 Social grooming network of Tibetan macaques. The nodes represent all adult individuals in the group; The node size is proportional to the node strength; The line thickness indicates the member’s correlation index in the social network; Males and females are indicated by blue and red squares, respectively; The letters on the label represent each individual’s name
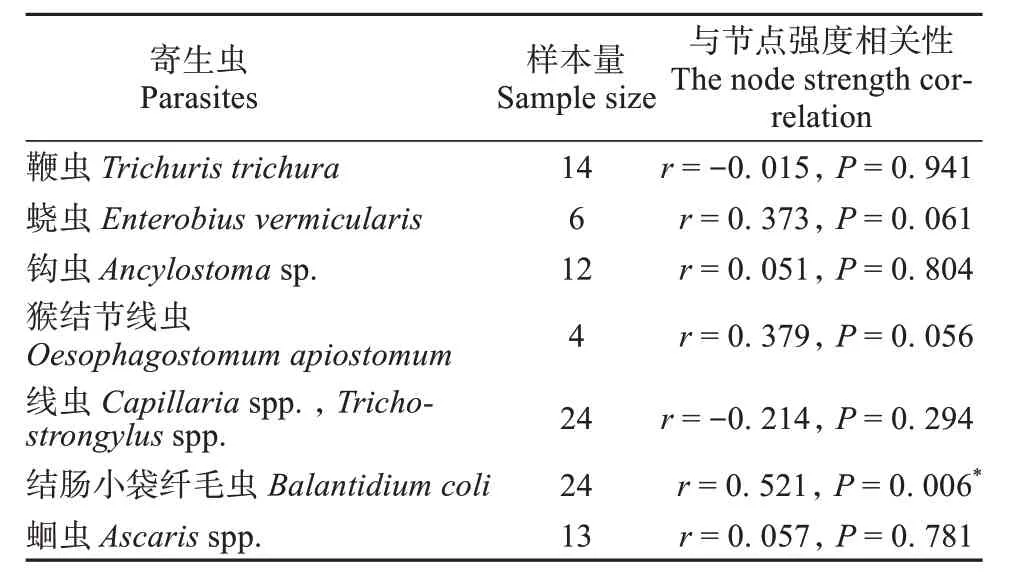
表4 藏酋猴肠道寄生虫负荷与节点强度的相关性Table 4 The correlation between intestinal parasite load and node strength of Tibetan macaques
3 讨论
本研究聚焦理毛行为与肠道寄生虫感染的种类与风险,发现野生藏酋猴群体中感染结肠小袋纤毛虫的概率最高,两两个体之间发起理毛行为持续时间越长,感染结肠小袋纤毛虫的风险越大,同时接收理毛时间越长,感染蛔虫的风险越低。进一步的分析发现,野生藏酋猴群体水平上,理毛网络中节点强度越大的个体,感染结肠小袋纤毛虫的风险越高,但是动物个体的节点强度与蛔虫感染负荷之间却无显著相关关系。结果说明,理毛行为能够增加一些肠道寄生虫感染的风险,为揭示理毛行为与特定肠道寄生虫种类的关系提供了直接证据,也为进一步理解理毛行为投资与收益关系提供了科学依据。
本研究发现野生藏酋猴感染11 种寄生虫,这一结果与前期在同一地点、同一研究对象的结果略有异同。吴伟等 (2018) 通过尼龙筛淘洗法,检测出10种寄生虫。鉴于寄生虫感染的多因素影响,如地域、环境、季节等 (Altizeret al., 2006),尽管野生藏酋猴不同时期感染寄生虫种类有所不同,但是在粪便中均检测到结肠小袋纤毛虫的包囊,且成年个体中具有很高的感染率 (84.62%,吴伟等,2018;93.00%,本研究)。结肠小袋纤毛虫是人体最大的寄生原虫,寄生于人的大肠中,可侵犯宿主的肠壁组织引起结肠小袋纤毛虫疟疾,其生活史简单不需要中间寄主,包括包囊和滋养体两个时期,也是监测动物感染与否的关键指标 (师永华等,2022)。结肠小袋纤毛虫在哺乳动物肠道中普遍存在,如猕猴 (Macaca mulatta) (林开铅等,2011;韦秋奖和王宏,2015)、食蟹猴 (Macaca fascicularis) (谢永平等,2019)、猪 (Annaet al.,2021) 等。近期的研究发现,在笼养环境下,一些非人灵长类动物的肠道中存在结肠小袋纤毛虫滋养体,如长臂猿 (Hoolock leuconedys) (邓长林等,2013)、食蟹猴 (谢永平等,2019)、实验猕猴 (林开铅等,2011)、白头叶猴 (Presbytis leucocephalus)(肖航,2021)。本研究为认识野生藏酋猴群体中结肠小袋纤毛虫较高感染率提供了更多证据,说明自然条件下野生灵长类动物也有感染结肠小袋纤毛虫的风险,且感染风险较高。
理毛行为的基本功能是清洁功能。本研究发现接收理毛行为时间越长的个体感染蛔虫的概率越低,为这一功能提供了证据。同时,理毛行为也具有显著的社会功能 (加强个体间社会联系等)(Dunbar, 1991, 2010; Spruijtet al., 1992; Xiaet al.,2019)。然而,病毒或寄生虫的感染或传播风险是理毛行为投资的一种代价 (Alexander, 1974; Mølleret al., 1993; Altizeret al., 2003),动物个体之间的直接接触会导致特定类型寄生虫感染的风险。在猕猴的研究中发现,与群体成员直接接触越频繁的个体,其寄生性原虫的感染率也越高 (Houptet al., 2016)。其他社会性动物的研究中也发现了相似的结果 (长颈鹿Giraffa camelopardalis, Vander-Waalet al., 2014;猫鼬Suricata suricatta, Drewe,2010),但是感染或传播的寄生虫种类有所差异,如在长颈鹿中社会接触行为与大肠杆菌的传播有关 (VanderWaalet al., 2014),猫鼬的理毛行为与牛分支杆菌 (Mycobacterium bovis) 的感染关系密切(Drewe, 2010)。
本研究发现,作为动物个体之间直接接触方式之一,理毛行为感染和传播结肠小袋纤毛虫的风险较高。
结肠小袋纤毛虫致病性不仅与不同虫株虫体本身的毒力有关,还与宿主的免疫状态和寄生环境有关 (孙志洲,2018;师永华等,2022)。尽管野外研究期间尚未发现动物个体有明显的发病症状,但是野外研究的可控性较低,样品检测的途径相对简单,动物个体的健康状况很难监测,因此理毛行为与结肠小袋纤毛虫感染的关系,尤其是对动物个体健康状况的影响很难定论。更为重要的是,本研究发现,群体水平上理毛网络中节点强度越高的个体,其感染结肠小袋纤毛虫的风险越高。这一结果提示我们,结肠小袋纤毛虫感染不仅是对特定个体理毛行为的代价,而且可能引起整个社会群体的感染风险,应该引起足够的重视。
本研究为了解理毛行为导致感染何种类型寄生虫以及感染风险提供了更多的证据支持,也为动物个体的投资与收益关系提供了更多的科学依据。然而,由于寄生虫种类繁多、生活史特点各异、鉴定指标多样 (包囊、滋养体、成虫等)、野生动物生存环境和群体内动物个体的社会特征 (年龄、顺位等) 也各不相同,因此未来的研究更要针对特定物种、特定环境、特定寄生虫种类以及感染后的健康评价等综合分析,一方面可以科学地监测野生动物的健康,另一方面可以为认识和理解动物个体社会行为的功能与代价提供更多证据支撑。同时,理毛时间长短只是寄生虫传播与否的一种影响因素,其他潜在因素也可能直接影响寄生虫的感染,或者通过影响动物个体之间的关系间接影响寄生虫感染,未来需要更深入的研究。
致谢:感谢安徽大学行为生态研究组全体成员对本文数据采集和处理过程中的帮助和支持!感谢安徽省寄生虫病防治研究所对本论文寄生虫检测的帮助!感谢黄山野生猴谷管理中心的工作人员对野外工作的支持和帮助!感谢房东程海滨一家对日常生活的帮助和照顾!
猜你喜欢
青少年科技博览(中学版)(2023年5期)2023-06-26
数学小灵通(1-2年级)(2022年10期)2022-12-27
时代英语·高三(2022年3期)2022-11-10
生态学报(2022年9期)2022-05-26
军事文摘(2021年22期)2022-01-18
当代水产(2021年5期)2021-07-21
小学生学习指导(中年级)(2020年9期)2020-10-24
时代英语·高三(2019年4期)2019-09-03
渔业科学进展(2018年1期)2018-04-03
海洋与湖沼(2016年1期)2016-01-15
