
海南长臂猿的毛色变化
2024-02-26 10:36祝常悦钟旭凯王昱心李文永齐旭明范朋飞
兽类学报 2024年1期
祝常悦 钟旭凯 王昱心 李文永 齐旭明 范朋飞*
(1 中山大学生命科学学院,广州 510275)(2 海南热带雨林国家公园霸王岭分局,昌江 572722)
长臂猿科 (Hylobatidae) 动物是分布于东南亚地区热带、亚热带森林中的小型类人猿。目前全球有4 属20 种 (Roos, 2016; Fanet al., 2017; Bartlett and Sheldon, 2022)。尽管部分颅骨和牙齿的形态学研究指出,白掌长臂猿 (Hylobates lar)、白眉长臂猿属 (Hoolock) 及合趾猿(Symphalangus syndactylus) 的颌面、眉骨或上犬齿存在一定程度的性别差异 (Frisch, 1963; Balolia, 2021),但相较于其他大型类人猿,长臂猿科动物种内雌雄个体在体型和犬齿上差异较小 (Leutenegger and Cheverud, 1985;Bartlett and Sheldon, 2022)。白眉长臂猿属和冠长臂猿属 (Nomascus) 所有物种以及长臂猿属的戴帽长臂猿 (H.pileatus),成年雌雄个体都表现出明显的毛色性二型:雄性除面部外的整体毛色以黑为主,而雌性偏浅,因物种和个体的差异呈现出浅褐、浅黄、橙黄、金黄等同色系不同梯度的毛色差异 (Mootnick and Fan, 2011; Harding, 2012)。合趾猿和长臂猿属除戴帽长臂猿外的其他物种成年个体的毛色性二型差异较小 (Roos, 2016)。
在灵长类动物中,个体发育过程中毛色的变化通常是性成熟的指征 (Bradley and Mundy, 2008;Light and Bartlett, 2017)。在僧面猴属 (Pithecia)、吼猴属 (Alouatta)、山魈属 (Mandrillus) 和狒狒属(Papio) 等类群中都有表现出毛色性二型的物种。这类物种中,雄性在进入第二性征发育后期会随着荷尔蒙的变化展现出更鲜艳夺目的毛色或更具装饰性的局部体色,而雌性个体则看起来更不显眼 (Clutton-Brock, 1985; Mittermeieret al., 2013)。但在长臂猿科的白眉长臂猿属和冠长臂猿属动物中,均为雄性保持着未成熟前的暗色 (周身以黑色为主),而雌性经历由暗至亮的毛色变化,其中以冠长臂猿属内的种间变化最为丰富 (Mootnick and Fan, 2011)。
毛色变化可作为判断长臂猿个体年龄的有效线索之一,白眉长臂猿属和冠长臂猿属的物种都存在随年龄发生毛色变化的现象 (Pocock, 1905;Geissmann, 1991; Mootnick, 2006; Mootnick and Fan, 2011; Harding, 2012)。在野外观察中,往往难以获得长臂猿个体的准确年龄。长臂猿的年龄组划分可能因不同物种甚至不同个体生活史的差异而大相径庭。综合6种来自笼养和野外长臂猿个体的发育记录 (Geissmann, 1991; Brockelmanet al.,1998; Reichard and Barelli, 2008; Zhouet al., 2008;Hodgkisset al., 2010; Burnset al., 2011; Harding,2012; Kimet al., 2012; Fanet al., 2013):长臂猿婴猿约2岁时断奶,4 ~ 5岁开始进入第二性征的发育期,7 ~ 9 岁左右完全性成熟;体型和第二性征均达成熟的个体约在8 ~ 10岁离开出生群,寻找领域和配偶;野外的雌性长臂猿约9 ~ 13 岁生育第一胎 (孕期6 ~ 7 个月),少数个体能存活40 岁并停止繁殖 (老年)。因此,当前主流的观点将长臂猿划分为5个年龄组 (Brockelmanet al., 1998),即0 ~ 2 岁为婴猿 (infant),2 ~ 5 岁为少年期 (juvenile 1),5 ~8 岁为青年期 (juvenile 2 or adolescent),8 岁至迁离出生群为亚成体 (sub-adult),而成年猿 (adult) 则指已组成新家庭群可繁育、参与二重唱的雌性或雄性 。详细描述冠长臂猿的毛色变化过程,有助于科研和保护工作者在复杂环境下 (野外条件、非法贸易、低质量的影像资料等) 进行物种识别和年龄估计,推算个体出生季节,以更加精确地推测种群的性比和年龄结构,服务这些濒危物种的种群保护和管理。
此外,研究物种毛色变化的过程可为种间系统发育关系的讨论提供宏观证据 (Groves, 1972;Roos, 2016)。现生的冠长臂猿属包含7 个物种:东黑冠长臂猿 (N.nasutus)、海南长臂猿 (N.hainanus)、西黑冠长臂猿 (N.concolor)、北白颊长臂猿(N.leucogenys)、南白颊长臂猿 (N.siki)、北黄颊长臂猿 (N.annamensis) 和南黄颊长臂猿 (N.gabriellae) (Yanget al., 2023)。东黑冠长臂猿与海南长臂猿互为姐妹群,西黑冠长臂猿与另外4种组成姐妹群 (Thinhet al., 2010b;范朋飞,2012)。东黑冠长臂猿新生婴猿均为黑色,雄性终身保持深色,有少量棕色毛发集中在胸口,甚至延伸至喉部和腹部。成年雌性东黑冠长臂猿个体周身以浅黄或橘黄色的毛发为主;由眉骨上方经颅顶变宽,直至延伸到颈后和肩膀的黑色冠纹沿脊柱逐渐变窄,在肩胛骨附近收成V 形;面部的黑毛外周环有一圈白色的长毛,使个体看起来脸型圆润,遂得“玉面金丝”的美誉 (Fanet al., 2010; Mootnick and Fan, 2011)。西黑冠长臂猿和北白颊长臂猿两性出生时均被米白色毛发,约1 ~ 3月龄逐渐变成金黄色,自半岁开始向深色过渡,至1.5 ~ 2 岁完全变为黑色,其中北白颊长臂猿出现下颌延伸到耳上缘的白颊;成年雄性不再变色,雌性在接近性成熟时会再次转为浅色,颅顶黑色冠纹的面积和形状因物种和个体的差异而不同 (Mootnick and Fan,2011; Harding, 2012; Huet al., 2018)。
海南长臂猿野外种群数量约40 只,是世界上最濒危的灵长类动物 (Geissmann and Bleisch,2020; Liuet al., 2022)。关于其毛色变化的过程还未见报道。本文以海南长臂猿为例,通过10 只个体的实际观察记录与影像资料,描述婴猿、雌性亚成体至成年、成年至老年等关键时期的毛色变化,为海南长臂猿的个体发育过程提供资料,为种群监测提供操作性建议。
1 研究方法
本研究对象为栖息在海南热带雨林国家公园霸王岭分局 (北纬18°48′ ~ 19°12′,东经108°55′ ~109°17′) 内的25 只海南长臂猿个体,来自4 个家庭群和1 只雌性独猿个体。其中涉及毛色变化的有10 只,分别为5 只雌性和5 只新生婴猿,其相关种群信息见表1。
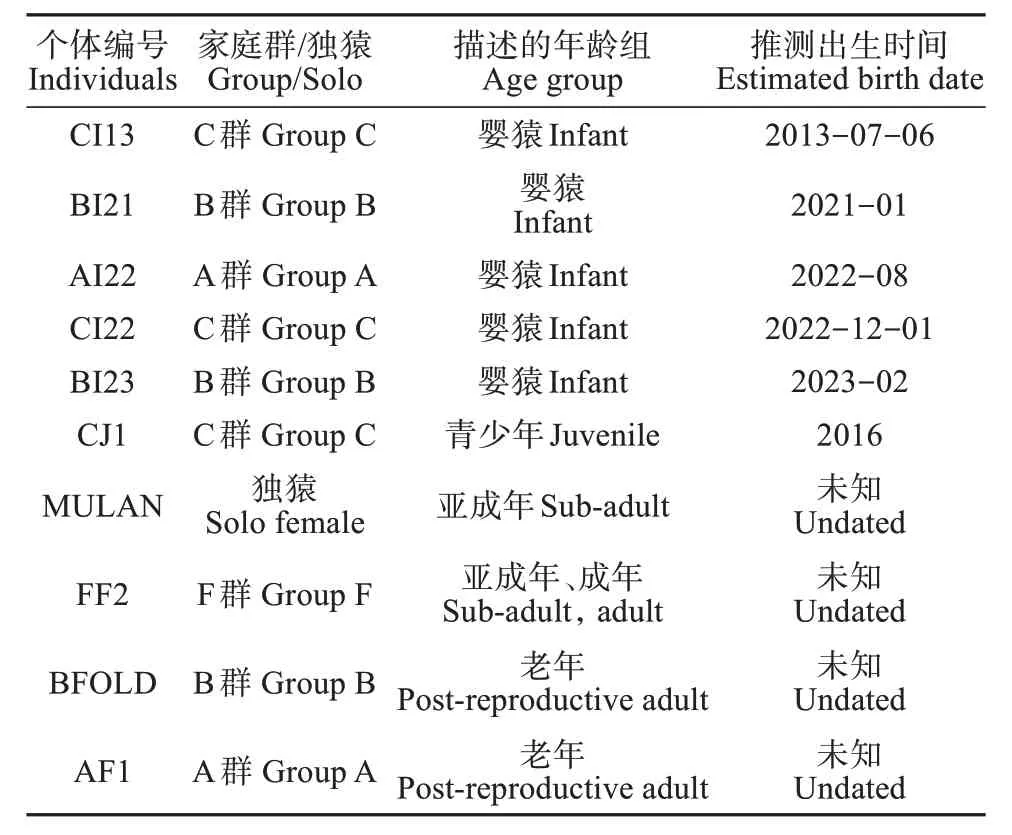
表1 涉及毛色变化描述的10只海南长臂猿个体信息Table 1 The ten N. hainanus individuals involved in the descriptions of the pelage colour change
本研究综合既有观点 (Liuet al., 1989; Geissmann, 1991; Brockelmanet al., 1998; Fanet al.,2010;韦绍干等,2017) 与具体观察,提出海南长臂猿野外观察和记录时建议使用的年龄组划分:出生至离开母亲独立活动为婴猿 (0 ~ 3 岁);本研究无法准确判断个体第二性征开始发育的时间,因此不区分少年和青年期,而是将独立活动至雌性开始变色、雄性开始练习鸣叫的这段时期为青少年 (3 ~ 8 岁);雌性毛色由黑转黄、雄性练习鸣叫至迁出前为亚成年 (通常8 岁以上);雌性毛色由黄转黑,并且停止繁殖为老年。由于青少年时期不存在毛色变化,本研究重点对海南长臂猿婴猿、亚成年至成年、成年至老年等关键发育阶段的毛色变化进行描述。
2021 年9 月至2022 年12 月,在霸王岭国家级自然保护区对已习惯化的长臂猿个体,每月进行至少30 h 的连续跟踪,在此期间观察到两只婴猿出生和一个新家庭形成。跟踪时使用8倍手持双筒望远镜进行观察。雌性个体识别主要基于整体毛发颜色、面周白毛的分布及冠纹形状等特征,未成年猿个体识别主要基于体型的相对大小及同成年雌性的近距与互动关系。对于未习惯化的个体,在每次目击时使用长焦数码相机进行影像记录。由于亚成年雌性的变色过程历时较长且个体可能在变色前就离开出生的家庭群而不易连续地观察。为避免臆测个体的年龄,借鉴Margulis 和Hálfdanardóttir (2021) 的评分制,根据个体外观黑、黄毛色占比将变色期雌性亚成年个体再细分为5 个阶段。
2 结果
2.1 婴猿的毛色变化
2.1.1 出生至一月龄婴猿的毛色变化
海南长臂猿婴猿出生后7日内,躯干及四肢大部分不被毛,略带粉红色的皮肤外露 (图1b、d 蓝色箭头)。后背靠近颈部处略覆浅黄色稀疏毛,并延至整个头部,且浓密程度随靠近颅顶而增加 (图1c、d黄色箭头)。颅顶及靠近前额的位置有小面积的黑色毛,构成类似雌性头顶冠纹的小小“黑斑”(图1a、c红色箭头)。

图1 出生后7日内的海南长臂猿婴猿及其母亲. a:出生后第2日的CI22与母亲CF1 (拍摄日期:2022-12-02);b:出生后第3日的CI22与母亲CF1 (拍摄日期:2022-12-03);c:出生后第6日的CI22与母亲CF1 (拍摄日期:2022-12-06);d:出生后第7日的CI22与母亲CF1 (拍摄日期:2022-12-07).(祝常悦摄)Fig. 1 Infant N. hainanus within seven days after birth and its mother. a: CI22 and its mother CF1, two days after birth, photographed on December 2, 2022; b: CI22 and its mother CF1, three days after birth, photographed on December 3, 2022; c: CI22 and its mother CF1, six days after birth,photographed on December 6, 2022; d: CI22 and its mother CF1, seven days after birth, photographed on December 7, 2022. (Photographed by Zhu Changyue)
出生后第3日,除了耳廓仍保持粉红色,其余不被毛的皮肤均开始微微地向暗色过渡,四肢的末端 (手指、脚趾) 变色最早,前肢又较后肢更早。第4 日起,脸周及耳后开始生出黑毛且逐渐增密,与仍偏浅色的面部及耳廓对比明显 (图2)。第5 日起观察到背部的毛由肩胛骨沿脊旁侧至盆骨逐渐增厚,呈浅黄色。第6日起,上肢开始微微覆浅黄色毛,十分稀疏,所有外露皮肤继续变暗,但仍呈灰粉色。颅顶稍显出非常短且稀疏的直立的簇毛,但还不具“冠”的形状 (图2b红色箭头)。

图2 出生后10日内的海南长臂猿婴猿和它们的母亲.a:出生后第4日的CI22与母亲CF1 (拍摄日期:2022-12-04,祝常悦摄);b:出生后第6日的CI13与母亲CF1 (拍摄日期:2013-07-12,李文永摄);c:出生后约第10日的AI22与母亲AF2 (拍摄日期:2022-09-02,李文永摄)Fig. 2 Infant N. hainanus within ten days after birth and their mothers. a: CI22 and its mother CF1, four days after birth, photographed on December 4, 2022, by Zhu Changyue; b: CI13 and its mother CF1, six days after birth, photographed on July 12, 2013, by Li Wenyong; c: AI22 and its mother AF2, ten days after birth, photographed on September 2, 2022, by Li Wenyong
出生后约2周,婴猿头部浅黄色的毛发逐渐增厚,背部已逐渐全部为浅黄色毛所覆盖,前肢毛发稀疏,后肢仍罕有毛发长出 (图3a)。耳廓和面部皮肤颜色继续变暗,四肢皮肤的颜色仍是前肢比后肢更早变黑,当手指已几乎完全变为黑色时,脚趾仍呈暗粉色 (图3b 蓝色箭头)。出生约3 周时,婴猿的躯干被浅黄色毛,腹侧疏背侧密,四肢毛发增量仍不明显。此时所有外露的皮肤已偏深色,与母亲的毛色对比十分明显 (图3c)。

图3 出生后2 ~ 3 周的海南长臂猿婴猿及其母亲.a、b:出生后第14日的CI22与母亲CF1 (拍摄日期:2022-12-14);c:出生后第20 日的CI22与母亲CF1 (拍摄日期:2022-12-20). (祝常悦摄)Fig. 3 Two to three weeks old infant N. hainanus and its mother. a, b: CI22 and its mother CF1, 14 days after birth, photographed on December 14, 2022; c: CI22 and its mother CF1, 20 days after birth, photographed on December 20, 2022.(Photographed by Zhu Changyue)
约1月龄的婴猿脑后、躯干及四肢均被浅黄色毛 (图4a、c),但四肢末梢的毛仍稀疏,皮肤的颜色已接近黑色 (图4b、d 蓝色箭头)。前额与耳后的黑毛继续增密 (图4c)。有些个体颅顶的“黑斑”微微向颅后延伸,似成一条“黑线”(图4a、c 红色箭头)。

图4 约1月龄的海南长臂猿婴猿和它们的母亲.a:出生后第31日的CI13 和母亲CF1 (拍摄日期:2013-08-06,李文永摄);b ~ d:约1月龄的BI23和母亲BF1 (拍摄日期:2023-03-26,钟旭凯摄)Fig. 4 One-month-old (circa) infant N. hainanus and their mothers.a: CI13 and its mother CF1, 31 days after birth, photographed on August 6, 2013, by Li Wenyong; b-d: One-month-old (circa) BI23 and its mother BF1, photographed on March 26, 2023, by Zhong Xukai
2.1.2 二月龄至周岁的毛色变化
2月龄的婴猿,从背侧望,以浅色为主 (图5a,c);面周的黑色毛逐渐延伸到颈前侧,且裸露的皮肤已完全呈黑色 (图5b、e 蓝色箭头),婴猿的头从正面看呈黑色 (图5d红色箭头)。

图5 约2 月龄的海南长臂猿婴猿和它们的母亲. a、b:约2 月龄的BI21 和母亲BF2 (拍摄日期:2021-03-31,钟旭凯摄);c ~ e:出生后第55、59 和65 日的CI13 和母亲CF1 (拍摄日期分别为:2013-08-30、2013-09-03、2013-09-09,李文永摄)Fig. 5 Two-month-old (circa) infant N. hainanus and their mothers.a, b: Two-month-old (circa) BI21 and its mother BF2, photographed on March 31, 2021, by Zhong Xukai; c-e: CI13 and its mother CF1, 55,59 and 65 days after birth sequentially, photographed on August 30,2013, September 3, 2013 and September 9, 2013, by Li Wenyong
3 ~ 5 月龄的婴猿,整体黑、黄毛色相杂 (图6a ~ e)。黑色毛比例逐渐增加的过程为腹侧先于背侧,四肢先于躯干。4 月龄婴猿的正面已几乎呈黑色 (图6c),而颅后、颈后、躯干背侧仍呈灰黄色(图6d),四肢外侧也零星地杂有浅色毛 (图6e)。5 月龄的婴猿颅顶开始出现短小的“冠”型,且背侧远观已接近黑色,仅“冠”两侧、颈后、肩胛骨附近和腰部有少面积的浅色毛 (图6f)。

图6 3 ~ 5月龄的海南长臂猿婴猿和它们的母亲.a:出生后第80日的CI13和母亲CF1 (拍摄日期:2013-09-24,李文永摄);b ~ f:约4 ~ 5 月龄的BI21 和母亲BF2 (拍摄日期分别为:2021-05-13、2021-05-26、2021-05-30、2021-06-07,范朋飞、钟旭凯摄)Fig. 6 Three- to five-month-old infant N. hainanus and their mothers. a: CI13 and its mother CF1, 80 days after birth, photographed on September 24, 2013, by Li Wenyong; b-f: Four- to five-month-old BI21 and its mother BF2, photographed on May 13, 2021, May 26,2021, May 30, 2021, and June 7, 2021 sequentially, by Fan Pengfei and Zhong Xukai respectively
10 月龄至1 周岁的婴猿几乎全身覆黑色毛 (图7a),“冠”型更为明显。从背侧望,仅在颈后到上背部有零星不易察觉的灰黄色;而从正面观,“冠”两侧有小面积的灰白色延至耳后 (图7b)。

图7 接近1 周岁的海南长臂猿婴猿及其母亲. a:约10 月龄的BI21 和母亲BF2 (拍摄日期:2021-10-23);b:约11 月龄的BI21和母亲BF2 (拍摄日期:2021-12-07).(祝常悦摄)Fig.7 Nearly one-year old infant N. hainanus and its mother. a: Tenmonth-old (circa) BI21 and its mother CF2, photographed on October 23, 2021; b: Eleven-month-old (circa) BI21 and its mother CF2, photographed on December 7, 2021. (Photographed by Zhu Changyue)
2.2 成年雄性的毛色变化
1 周岁后,无论雌雄,海南长臂猿个体将保持周身黑色至青少年末期,直至雌性第二次开始变色;而雄性终生为黑色,不再发生毛色变化。成年雄性覆较短的黑色毛,头顶直立的簇毛形成冠,成年雄性的冠毛较亚成年雄性更浓密且稍向额前倾 (图8)。

图8 海南长臂猿亚成年雄性与成年雄性. a:约9岁的原B群亚成年雄性BSA,于2022年5月迁离出生群 (拍摄日期:2022-04-21);b:年龄不详的成年雄性CM1,2011 年发现时已成为C 群主雄 (拍摄日期:2022-11-01). (祝常悦摄)Fig. 8 Sub-adult and adult male N. hainanus. a: Nine-year-old (circa) sub-adult male BSA who was born in Group B and left its natal family in May 2022, photographed on April 21, 2022; b: Adult male CM1 who has been the dominant male in Group C since 2011 with unspecified age, photographed on November 1, 2022. (Photographed by Zhu Changyue)
2.3 亚成年雌性的毛色变化
海南长臂猿雌性个体在亚成年阶段毛色会逐渐从黑色变为成年雌性的橙黄或亮黄色,该阶段持续约20 个月,根据黑、黄两色在整体外观中所占的比例,又细分为5个阶段。
第一阶段:当雌性个体的体型明显大于同群的青少年个体时,下颌至颈前侧开始微微显出棕红色,“冠”两侧毛色开始变浅 (图9a)。变色较早出现在下颌、颈前侧、腋下和下肢内侧。伴随下颌浅灰色毛面积逐渐增大,腋下和四肢内侧的毛开始逐渐由棕红色向灰黄色过渡,且逐渐扩展至胸口 (图9b ~ d)。此阶段的个体从背侧观察,毛色仍以黑色为主,且保有青少年期“冠”的形状。

图9 亚成年雌性毛色变化第一阶段举例. a:约6岁的雌性青少年CJ1 (拍摄日期:2022-11-06,祝常悦摄);b ~ d:亚成年雌性独猿MULAN (拍摄日期:2022-08-13、2022-08-23、2022-09-04,祝常悦、李文永摄)Fig. 9 Examples of sub-adult female N. hainanus who were undergoing the first stage of pelage colour change during sexual maturity. a:Six-year-old (circa) female CJ1, photographed on November 6, 2022,by Zhu Changyue; b-d: Sub-adult female MULAN, photographed on August 13, 2022, August 23, 2022 and September 4, 2022 sequentially,by Zhu Changyue and Li Wenyong respectively
第二阶段:黄灰色的面积逐渐扩大到四肢的外侧 (上肢先于下肢) 和腹部 (图10a)。此时躯干的背侧及臀部开始出现零星灰黄色的毛。约3 个月内,个体的上肢将完全替换为灰黄色的毛,下肢仍黑黄相杂且越靠近足部颜色越深。此阶段,个体从背侧观察已整体呈灰色,黑色毛仅集中在胸前、脑后、除下颌外的面周和“冠”的位置 (图10b、c)。

图10 亚成年雌性毛色变化第二阶段举例. a:亚成年雌性独猿MULAN (拍摄日期:2022-11-18);b、c:F 群成立前结伴生活的2 只雌性中较年轻的FF2 (拍摄日期:2021-08-06). (祝常悦摄)Fig. 10 Examples of sub-adult female N. hainanus who were undergoing the second stage of pelage colour change during sexual maturity. a: Sub-adult female MULAN, photographed on November 18,2022; b, c: Sub-adult female FF2 who used to accompany with an adult female FF1 before the formation of Group F, photographed on August 6, 2021. (Photographed by Zhu Changyue)
第三阶段:开始变色后的半年内,躯干及四肢的灰黄色毛逐渐向橙黄色过渡,上肢比下肢更早变得色泽鲜亮。此阶段个体周身远观已呈黄色,只有颅顶、前额和耳周颜色较暗,且“冠”的形状开始逐渐变圆 (图11)。

图11 亚成年雌性毛色变化第三阶段举例. F群成立前结伴生活的FF2 (左) 和FF1 (右) (拍摄日期:2022-01-21,祝常悦拍摄)Fig. 11 Example of sub-adult female N. hainanus who was undergoing the third stage of pelage colour change during sexual maturity.Sub-adult female FF2 (left) and its partner adult female FF1 (right) before the formation of Group F, photographed on January 21, 2022.(Photographed by Zhu Changyue)
第四阶段:开始变色约一年后,个体的“冠”形已完全消失,从正面无法观察到耳朵,颅顶的黑色冠纹较成年雌性面积更大但颜色偏灰。此阶段,个体全身毛色已接近橙黄色,上肢及肩胛部颜色更深,面周、颈部、胸口、腰臀部和下肢颜色较浅 (图12)。

图12 亚成年雌性毛色变化第四阶段举例. a、b:FF2 (拍摄日期:2022-04-29、2022-05-24);c:FF2 (左) 和FF1 (右) (拍摄日期:2022-06-12). (祝常悦摄)Fig. 12 Example of sub-adult female N. hainanus who was undergoing the fourth stage of pelage colour change during sexual maturity. a,b: Sub-adult female FF2, photographed on April 29, 2022 and May 24,2022; c: Sub-adult female FF2 (left) and adult female FF1 (right), photographed on June 12, 2022. (Photographed by Zhu Changyue)
第五阶段 (图13a,b):毛色为鲜艳的橙黄色,除因尚未哺乳而乳头短于已生育个体外 (图13c ~e),外观上已与成年雌性无异。

图13 亚成年雌性毛色变化第五阶段举例与已哺乳雌性举例. a:未哺乳雌性FF2 (拍摄日期:2022-10-09,祝常悦摄);b:未哺乳雌性FF1 (拍摄日期:2022-09-26,祝常悦摄);c:CF2,2020 年末次哺乳 (拍摄日期:2021-11-05,祝常悦摄);d:BF2,2022 年末次哺乳 (拍摄日期:2023-03-26,钟旭凯摄);e:CF1,拍摄时正在哺乳期 (拍摄日期:2022-12-06,祝常悦摄)Fig. 13 Examples of female N. hainanus who were undergoing the fifth stage of pelage colour change during sexual maturity and the compared examples of adult female N. hainanus who have had offspring and experiences of breastfeeding. a: FF2 who has not had offspring yet, photographed on October 9, 2022, by Zhu Changyue; b: FF1 who has not had offspring yet, photographed on September 26, 2022, by Zhu Changyue; c: CF2, the last lactating time was in 2020, photographed on November 5, 2021, by Zhu Changyue; d: BF2, the last lactating time was in 2022, photographed on March 26, 2023, by Zhong Xukai; e: Lactating female CF1, photographed on December 6, 2022,by Zhu Changyue
2.4 老年雌性的毛色变化
伴随着年龄增长,老年雌性的毛色逐渐变暗,全身再次转为以灰黄色为主,颅顶及躯干背侧甚至完全变为黑色。A 群的老年雌性 (AF1) 背侧远观呈暗棕色 (图14d ~ f),从颅顶至后颈沿着脊椎到腰臀部已大量覆黑毛 (图14f),但头顶仍保持雌性的圆形而不具“冠” (图14a ~ c)。它的四肢内侧和躯干腹侧仍偏黄,但胸口和颈前侧已呈黑色 (图14b)。成年雌性眼眶、口周的白毛在此阶段已完全变黑,只有面周和耳周的毛呈灰黄色 (图14a、e)。B 群曾有一只21 世纪以来未观测到繁殖的老年雌性 (BFOLD),约于2015 年消失 (韩玲,2019)。从影像资料中可以看出,与AF1 相似,它的头、后背和颈前侧毛色变暗 (图15a),四肢外侧,尤其是肘和膝关节处的毛呈黑色 (图15b)。

图14 已接近14年未繁殖的AF1. a、e、f:AF1 (拍摄日期:2022-03-02,祝常悦摄);b ~ d:AF1 (拍摄日期:2022-09-02,李文永摄)Fig. 14 Post-reproductive adult female N. hainanus AF1 who has not been observed being breeding since 2009. a, e, f: AF1, photographed on March 2, 2022, by Zhu Changyue; b-d: AF1, photographed on September 2, 2022, by Li Wenyong

图15 于2015年已消失的BFOLD (拍摄日期:2008-08-12,赵超摄)Fig. 15 Post-reproductive adult female N. hainanus BFOLD who has disappeared in 2015 (photographed on August 12, 2008, by Zhao Chao)
3 讨论
3.1 婴猿出生季节推断
野外观察长臂猿个体难度极大。对于习惯化的个体,监测人员可在约20 m 外安静地进行观察;而未习惯化的个体通常非常警觉,如果是有新生猿的家庭群,特别是怀抱婴猿的雌性,警惕度会更高。详细描述新生婴猿的毛色变化过程有利于在野外观察时反推婴猿的出生日期,是了解家庭群内成员年龄结构和监测个体成长发育的关键信息。根据野外观察的经验,孕末期的待产雌性可能远离家庭群数日,直至后代出生。此过程若为未习惯化程度很高的群体,几乎不可能观察到。新生婴猿被监测人员首次目击的时间通常在出生后一周至数月 (Brockelmanet al., 1998; Bryantet al., 2016;韦绍干等,2017)。此时,通过毛发的疏密、肤色及毛色反推大致的出生日期就成为唯一的手段。在一夫二妻的长臂猿家庭中,可能出现两只可繁殖雌性在较短的时段内先后生产的情况 (Fanet al., 2015; Huet al., 2018;韩玲,2019),分辨新生婴猿外观形态所处的不同阶段亦有利于进行个体识别。
3.2 雌性毛色与年龄判断
雌性个体由青少年时的黑色完全变色至与成年雌性外观上无差异需2年的时间,且可能在开始变色之前就离开出生群。在进行年龄判断时,结合个体的毛色变化与行为及个体生活史是必要的。如6 岁的CJ1 在表现出最初的亚成年变色特征时,即“冠”两侧毛色开始变浅之前已持续参与C群内雌性的激动鸣叫约1年,且响度及熟练程度明显高于群内年龄更小的青少年个体。雌性独猿MULAN在亚成年雌性毛色变化的第一阶段已开始练习雌性邀配的“程式化舞蹈” (Fanet al., 2016)。新形成家庭群 (F 群) 中较年轻的雌性FF2 的下颌边缘和踝部的毛色仍为明显发白的变色末期,已与群内雄性发生交配行为。
雌性个体进入老年阶段,繁殖能力降低或全无,已不再同群内任何雄性交配,但仍被观测到留在家庭群内,与其它成员一同活动 (Zhouet al.,2008; Denget al., 2017; Huet al., 2018)。掌握种群中有繁殖潜力雌性的数量和分布对于分析和评估该种群的生存力及可能面临的灭绝风险至关重要(Fanet al., 2013; Liuet al., 2022)。Mootnick 和Fan(2011) 指出,西黑冠长臂猿滇西亚种的模式标本(KIZ 830353) 较其他成年雌性偏黑的毛色可能反映出老年个体因雌激素水平降低而无法维持金黄、橙黄、浅黄或浅褐等冠长臂猿雌性的典型毛色。相似的例子还有美国德克萨斯州格拉迪斯波特动物园 (Gladys Porter Zoo, Texas, USA) 1 只在34 岁时被切除子宫的雌性北白颊长臂猿,术后1年内,它的背部最先开始出现小面积的黑色;术后第6 年,除手、腕、足、上肢内侧、前臂外侧及下颌与锁骨处可见少量灰白色毛发,周身已几乎完全呈黑色 (Mootnick and Fan, 2011)。广西靖西邦亮的东黑冠长臂猿G2 群内的雌性个体F2 亦观察到毛色偏暗,其年龄可能也接近繁殖的末期 (马长勇和范朋飞,未发表数据)。本研究中AF1和BFOLD的例子为分辨老年雌性变黑与亚成年雌性变黄这两个逆向的过程提供了2 个技巧:(1) 亚成年雌性头部和腹侧的毛色最早开始变浅,而老年雌性头部和背侧的毛色最早开始变暗;(2) 亚成年雌性在变色过程中,青少年时期的“冠”形逐渐消失,而老年雌性毛色变暗的过程中颅顶保持成年雌性的圆形。如果有条件观察到个体的面部细节,可注意到老年个体裸露的皮肤上有较多皱纹。
3.3 毛色变化模式与系统发育关系
海南长臂猿新生儿毛色较西黑冠长臂猿和其它4种浅颊长臂猿偏黄,且出生即在前额可见冠纹样的黑毛。除了东黑冠长臂猿新生儿毛色为黑色外,海南长臂猿婴猿由浅至深的毛色变化发生时间最早,且接近周岁时已完全变黑。就婴猿的毛色和变色模式而言,海南长臂猿处于东黑冠长臂猿与西黑冠长臂猿之间,这与基于分子证据构建的系统发育树是吻合的 (Thinhet al., 2010b; Roos,2016)。婴猿出生皮肤颜色迅速变暗可能是抵御紫外线辐射的黑色素沉积,以避免细胞和DNA 结构的损伤 (章誉兴等,2021)。黑色毛发亦有保暖作用 (Mootnick and Fan, 2011)。分子钟证据表明,冠长臂猿属被认为由北方较高纬度地区在距今约1 050 万年扩散至印支半岛东部,又在距今150 ~30 万年间完成了最后的属内分化,形成今天分布在中南半岛最南端和海南岛的类群 (Chatterjee,2006; Thinhet al., 2010a)。作为属内最古老的物种,东黑冠长臂猿的新生儿可能保有了适应高纬度、高海拔、低温、强紫外线辐射的毛色特征。长臂猿新生儿的毛色受到系统发育关系的影响,但是产生属内变异机制仍不得而知,或可作为研究灵长类新生儿毛色演化的一个重要案例。灵长类动物中,头部和四肢的毛色被认为相对保守(Bellet al., 2021),新生儿身体各部位毛色的变化顺序也应是值得注意收集的信息。
在灵长类动物中,美狐猴属 (Eulemur) 的12个物种皆为雌性较雄性拥有相对更浅或更亮的毛色(Mittermeieret al., 2013)。其中婴猴变色模式与东黑冠长臂猿相似的例子有红额美狐猴 (Eulemur rufus) 和黑美狐猴 (E.macaco)。红额美狐猴婴猴出生时毛色与成年雄性相同,雌性个体于出生后7 ~17 周逐渐变色并过渡到与母亲一样的毛色,变色与婴猴离开母亲逐渐独立活动的时期相重合 (Bartholdet al., 2009)。Barthold 等 (2009) 认为,出生时与雄性同色可以帮助雌婴度过最脆弱、最易被攻击的阶段,这种模仿雄性而混淆性别的毛色伪装让雌婴得以在激烈的雌性攻击和竞争中存活下来。针对新生儿与成年个体强对比色的肤、毛,已提出吸引照护、预防杀婴和父权混淆 (paternity cloak) 等假说 (Caroet al., 2022),但不同类群的具体表现可能受到系统发育关系的制约 (Larson,1985; Treves, 1997)。如在山魈属、狒狒属、狮尾狒狒属 (Theropithecus) 和猕猴属 (Macaca) 等睾丸与体重比大的多雄社会中,新生儿的毛色偏灰暗,在视觉上不显眼;而睾丸与体重比较小的疣猴亚科 (Colobinae) 通常为单雄群,新生婴猴的毛色为夺目的浅色。冠长臂猿属新生的婴猿大多与母亲颜色相近,可能是一种保护策略,被母亲携带时融入背景色,离开母亲时有警戒作用 (Caroet al.,2021)。而随着年龄增长离开母亲独立活动后,外貌上不具明显亲缘特征的毛色可能更有助于幼猿和青少年个体免受群内雄性的攻击。
未成年雌性个体在青少年阶段保持与雄性相同的深色,已有通过混淆性别以避免近交并提高出生群对未迁出雌性容忍度的假说 (Geissmann,1993; Harding, 2012)。而雌性从亚成年到成年,由暗至亮的毛色变化模式仍值得讨论。不同于雌性选择的性选择假说,即雄性通过更为明亮、华丽的毛色展现优异的健康状况并暗示可繁殖出高质量的后代 (Bradley and Mundy, 2008);长臂猿科动物中存在性二色的物种,雌雄虽然外观在色彩上差异巨大,但没有任何一方更显眼而另一方更隐蔽 (Bartlett and Sheldon, 2022)。这可能引出另一种假设:成年个体雌雄的二色性可能表现出对复杂热带、亚热带森林环境的适应。在植被密度高而种群密度相对低的环境中,视觉上的颜色差异类似于具有性别指示作用的鸣声,能够远距离判断个体的性别,利于防御领地、震慑同性竞争者、寻找配偶。毛色的明亮程度变化受雌激素水平和个体所处社会环境的影响 (Geissmann and Anzenberger, 2009; Rafaczet al., 2013; Bolechováet al.,2019; Margulis and Hálfdanardóttir, 2021),对于性红肿不明显的冠长臂猿,通过毛色可以有效地估计雌性的年龄和繁殖状态。本研究中处于孕期和刚经历生产的雌性个体,毛色明显较非孕期更为鲜亮,为明亮的橙黄色或浅黄色 (图16 b,d),而非孕期的毛色偏白且暗淡 (图16 a,c)。

图16 非孕期与孕期或刚生产雌性海南长臂猿毛色对比举例.a:非孕期的可繁殖雌性BF1 (拍摄日期:2021-05-30,钟旭凯摄);b:孕期的BF1 (左) 和成年雄性BM1 (右),BF1 推测于2023年2 月生产 (拍摄日期:2022-10-12,王昱心摄);c:非孕期的可繁殖雌性CF1 (拍摄日期:2022-04-10,祝常悦摄);d:产后2 周的CF1 (拍摄日期:2022-12-14,祝常悦摄)Fig. 16 Examples of the non-pregnant fertile female N. hainanus and their compared images that has been pregnant or just has delivery.a: Non-pregnant fertile female BF1, photographed on May 30, 2021,by Zhong Xukai; b: Pregnant female BF1 (left) who has probably had the delivery in February 2023 and adult male BM1 (right), photographed on October 12, 2022, by Wang Yuxin; c: Non-pregnant fertile female CF1, photographed on April 10, 2022, by Zhu Changyue; d:CF1, two weeks after delivery, photographed on December 14, 2022,by Zhu Changyue
3.4 海南长臂猿种群监测的展望
不同于笼养个体可频繁、规律、近距离地被观察和记录毛色及其他形态特征的变化 (Barelli and Heistermann, 2009),本研究中对“冠”形、整体毛色、身体各部位变色顺序的描述可以在野外观察中帮助监测人员快速判断个体所处的大致年龄组,甚至推断婴猿的出生季节、预估亚成年迁离出生群时间、识别有繁殖潜力的雌性。近年来,随着监测力度增大,海南长臂猿观测到个体迁出、形成新家庭群及婴猿出生的频次上升 (Bryantet al., 2016; Denget al., 2017; Chanet al., 2020a; Liuet al., 2022),既是保护工作初具成效的好消息,也对一线监测工作提出了更高的要求和新的挑战。
尽管本研究强调了人力跟踪观察对了解海南长臂猿种群动态和个体发育的重要性,积极应用新技术,尤其是自动化的数据收集方法是野外监测的必然趋势。嘉道理农场暨植物园与海南大学等团队的工作已证明在重要的食物斑块、树冠层通道和过夜地附近的林冠层安装红外相机,能够拍摄到已习惯化海南长臂猿的清晰影像,并实现雨林环境中的自动识别 (Chanet al., 2020b; Wanget al., 2022)。对于未习惯化的家庭群和独猿个体,无人机搭载热成像 (Zhanget al., 2020) 与地面人工观测相结合、5G 实时传输网络 (陈耀亮,2022) 与红外相机相结合等方式可提升海南长臂猿野外监测的时效性和准确度,得以尽早监测到婴猿出生或亚成体离群等重要种群结构变化,甚至有可能发现新家庭群的形成,为濒危动物的保护决策提供严谨、精确的数据。
致谢:感谢海南热带雨林国家公园霸王岭分局、海南热带雨林国家公园研究院对本研究的支持。海南热带雨林国家公园霸王岭分局的冯业兴、符和能、黄卢标、李全金、刘辉勤、杨先珊,中山大学生命科学学院的阿合博塔·哈孜太、王子荻、杨与参与野外调查,特此感谢。
猜你喜欢
小哥白尼(野生动物)(2022年7期)2022-09-20
世界科学技术-中医药现代化(2021年5期)2021-11-05
养猪(2021年4期)2021-08-26
云南画报(2021年7期)2021-08-06
杂文月刊(选刊版)(2021年5期)2021-06-02
现代园艺(2018年2期)2018-03-15
畜牧兽医科学(2018年14期)2018-02-14
湖南中医药大学学报(2015年1期)2016-01-06
畜牧兽医学报(2015年3期)2015-07-05
小学生作文·小学中高年级适用(2014年3期)2014-08-18
