
太行山猕猴的性打搅行为
2024-02-26 10:36韩梦亚田军东周言言王钰炜罗彤彤路纪琪
兽类学报 2024年1期
韩梦亚 田军东 周言言 王钰炜 罗彤彤 路纪琪*
(1 郑州大学生命科学学院,郑州 450001)(2 郑州大学生物多样性与生态学研究所,郑州 450001)
性打搅 (Sexual interference) 是指动物在交配期间受到同种其他个体化学信息素、面部表情、声音、攻击等干扰的现象 (Arnold, 1976)。性打搅行为在非人灵长类动物普遍存在 (Gouzoules,1974; Niemeyer and Anderson, 1983; Thierry, 1986;Qiet al., 2011; Dixson, 2012)。迄今,已对30 多种灵长类动物的性打搅行为作了研究 (杨斌等,2013)。中国的非人灵长类物种丰富 (Fan and Ma,2018),但有关性打搅的研究报道仅见于川金丝猴(Rhinopithecus roxellana) (Li and Zhao, 2007;杨斌,2009; Qiet al., 2011) 和藏酋猴 (Macaca thibetana)(Xiong, 1993;庞奎海,2019;庞奎海等,2020;Panget al., 2021) 等少数物种。
根据打搅强度,可将性打搅行为分为性干扰(Sexual harassment) 和性打断 (Sexual interruption)(Bruce and Estep, 1992; Dixson, 2012)。性干扰的强度相对温和,打搅者通过趋近交配对、瞪眼、龇牙咧嘴、尖叫或拍打等行为,干扰交配活动 (Thierry, 1986; Dixson, 2012),通常并不造成交配行为的中止 (Qiet al., 2011)。性打断强度则较为激烈,打搅者对交配中的个体实施撕咬、抓打、拉扯或追赶等攻击性干扰行为,往往导致交配行为中止(Dixson, 2012)。对非人灵长类物种性打搅的诸多研究表明,性打搅行为的频次、方式和强度等表现出种间和种内差异。种间差异受社会结构和婚配制度的影响,而种内差异主要受性别、年龄和社会等级等因素的影响 (Niemeyer and Anderson,1983; Dixson, 2012;杨斌等,2013)。如川金丝猴成年和亚成年雌性的性打搅频次最高,可打断正在进行的交配;成年雄性的打搅频次较低,但均指向交配中的雄性,大多导致交配行为的终止;青年猴多打搅雌性个体的交配,并不导致交配活动的终止;幼猴仅打搅其母亲的交配,通常会打断交配 (杨斌,2009)。藏酋猴成年雄性的性打搅频次最高,性打断一般由高等级雄性发起;成年雌性多打搅亲缘个体的交配,并不打断交配;少年猴打搅频次较低,多打搅其母亲的交配,有交配行为打断的可能 (庞奎海,2019)。黑猩猩 (Pan troglodytes) 未成熟个体的性打搅频次最高且均为性干扰,雌性幼猴仅打搅其母亲和亲缘个体的交配,而雄性幼猴还会打搅非亲缘个体的交配;成年雄性通过直接的攻击行为打搅较低等级雄性的交配尝试;成年雌性则很少发起性打搅,其动机似乎为争夺交配中雄性的注意力 (Tutin, 1979)。
针对不同性别-年龄组个体的性打搅特征,学者们迄今已提出多种假说,试图解释性打搅行为的动机和功能。对成年雄性性打搅的解释主要是雄性性竞争假说 (The male sexual competition hypothesis),即成年雄性的性打搅是为减少群内其他雄性的交配成功率,或者为得到与被打搅雌性未来的交配机会而采取的一种性竞争策略 (Niemeyer and Anderson, 1983)。对成年雌性性打搅的解释主要是雌性性竞争假说 (The female sexual competition hypothesis) (Hrdy, 1977; Niemeyer and Anderson, 1983) 和占有假说 (The possessive hypothesis)(Niemeyer and Chamove, 1983; Brereton, 1994) 等。雌性性竞争假说认为,雌性个体会对群内高质量的精子资源展开竞争,通过打搅交配活动,减少其他雌性的生殖机会,从而使自己获得更多的交配机会。该假说可再分为直接的性竞争假说 (Niemeyer and Anderson, 1983) 和间接的性竞争假说(Hrdy, 1977),前者是指雌性打搅者通过减少交配雌性的生殖成功率而增加自身的成功率;后者是指雌性打搅者通过打搅交配活动,以引起交配雄性的注意,从而使自己随后有可能与该雄性交配。占有假说认为,当成年雌性交配时,与其关系亲密的成年雌性担心自身社会关系被打断而实施性打搅。对未成熟个体性打搅的解释主要基于母婴冲突假说 (The parent-offspring conflict hypothesis)(Tutin, 1979)、保护假说 (The protective hypothesis)(Gouzoules, 1974)、学习假说 (The learning hypothesis) (Dixson, 2012) 以及分离焦虑假说 (The separation anxiety hypothesis) (Tutin, 1979; Niemeyer and Anderson, 1983) 等。母婴冲突假说认为,未成熟个体通过打搅其母亲的交配,以减少或推迟下一个同胞后代的出现,从而使自己获得母亲更多的抚育。基于对红面猴 (Macacaarctoides) 的研究,Gouzoules (1974) 提出了保护假说:雄性红面猴在与雌性交配或者刚射精完毕时会咬交配伙伴,而与交配雌性关系密切的个体 (尤其是交配雌性的后代) 则试图通过打搅使交配雌性免受交配雄性的攻击。学习假说认为,未成熟个体的性打搅行为是其学习成熟社会行为的一部分。分离焦虑假说认为,性打搅是打搅者担心其与交配中的一方的社会关系被打断的一种情绪焦虑反应。
就已有的研究报道来看,尚无任何一种假说适用于所有的物种、性别-年龄组等 (Dixson,2012)。关于非人灵长类性打搅行为性别-年龄组差异的研究尚缺乏充分的案例支持。为此,系统地研究性打搅行为的性别-年龄组间差异,对阐明各性别-年龄组性打搅行为的表型差异,进而探讨其功能差异具有重要意义。
猕猴华北亚种 (Macaca mulatta tcheliensis) 为中国所特有 (蒋学龙等,1991; Brandon-Joneset al., 2004),其野生种群目前仅分布于河南、山西两省交界的太行山南端地区,故又称太行山猕猴(Luet al., 2007;路纪琪,2020,2022)。迄今为止,已有关于太行山猕猴的繁殖生态 (侯进怀等,1998)、社会结构 (田军东等,2011b)、行为谱 (田军东等,2011a)、觅食行为生态 (吕九全等,2002;郭相保等,2011)、社会联属 (鹿鹤等,2013)、营养生态 (Cuiet al., 2018, 2019, 2020) 以及遗传结构(Zhouet al., 2023) 等方面的研究报道,但尚缺乏对太行山猕猴性打搅行为的深入探讨。本研究旨在分析太行山猕猴的性打搅行为的特征及其性别-年龄组间差异,从而进一步揭示其动机和功能差异,并对相关假说进行验证,以期丰富和补充对非人灵长类动物性打搅行为的表型差异及其功能差异的认识。
1 研究方法
1.1 研究地点
本研究地点位于太行山猕猴国家级自然保护区 (北纬34°54′ ~ 35°42′,东经112°02′ ~ 113°45′)所辖的五龙口地区 (北纬35°12′49″,东经112°41′25″)。该保护区以猕猴、林麝 (Moschus berezovskii)、金钱豹 (Panthera pardus) 和金雕 (Aquila chrysaetos) 等物种为主要保护对象。研究地区处于暖温带大陆性季风气候区,夏季湿热 (最高气温43.4 ℃),冬季干冷 (最低气温-20.0 ℃),年平均气温14.3 ℃,平均降雨量695 mm (胡玉梅等,2004; Luet al., 2007)。
五龙口地区位于河南省济源市东北约15 km处,该地区山势陡峭,最高海拔1 058 m (剑过顶),最低海拔132 m,森林植被较为退化,以野皂荚(Gleditsia microphylla)、黄刺玫 (Rosa xanthina) 和山桃 (Prunus persica) 等灌丛为主,仅在山顶残存小面积天然次生林 (宋朝枢和瞿文元,1996)。
1.2 研究对象
栖息于五龙口地区的猕猴主要有5个群,基于我们的长期观察记录,依其群体大小及活动区域等,分别命名为WLK (Wulongkou,五龙口;下同) -1A 群、WLK-1B 群、WLK-2 群、WLK-3 群和WLK-4 群。其中WLK-1A、WLK-1B、WLK-2 和WLK-3群主要活动于研究地区的核心区域,WLK-4群则主要活动于后山区域。该地区太行山猕猴可自由取食自然食物 (植物嫩叶、芽、野果等),并有少量补充食物,包括玉米、小麦和花生,并辅以胡萝卜等 (Guoet al., 2020)。
本研究以太行山猕猴WLK-1A 群为目标群,根据形态 (体表被毛的长度、颜色、身体疤痕等)和行为特征 (攻击性、活跃程度) 等,对群内所有个体进行识别并命名,同时依据田军东等 (2011a)个体识别的标准,对群内的个体进行性别-年龄组划分。观察对象为群内所有个体,包括成年雄性10 只,成年雌性36 只,亚成年雄性2 只,亚成年雌性5只,青少年21只,婴幼猴19只。
1.3 行为定义
在本研究中,交配指1 只雄性个体爬跨1 只雌性个体,雄性生殖器插入雌性阴道内,进行抽插运动,交配往往伴随着射精或无射精 (侯进怀等,1998;何民强,2020)。雄性射精时,常表现出身体痉挛,并有张开嘴、露齿及发生鸣叫等现象,排出的精液迅速凝固,呈乳白色;雌性则回望雄性,并出现咧嘴、呜鸣等现象,或以前肢抓拽雄性 (何民强,2020)。本研究依此判断交配过程有无射精发生。
参考田军东等 (2011a) 的方法,本研究将太行山猕猴的性打搅行为划分为挨坐 (contact sitting)、追赶 (chasing)、趋近 (approaching)、理毛 (grooming)、带有威胁的注视 (staring with threaten)、抓打(seizing)、撕咬 (bite) 和钻挤 (squeezing),并对各种行为分别予以定义 (表1),其中钻挤为本研究新增行为。

表1 行为参数定义Table 1 Definitions of behavior parameters
依据打搅者与被打搅者是否存在物理性接触,将性打搅行为分为接触式性打搅 (contact harassment) 和非接触式性打搅 (non-contact harassment)。接触式性打搅指在交配发生时,打搅者与被打搅者发生身体接触 (挨坐、理毛、抓打、撕咬、钻挤等);非接触式性打搅指打搅者与被打搅者无任何身体接触 (追赶、趋近、带有威胁的注视等) (杨斌,2009; Qiet al., 2011;庞奎海,2019)。
被打搅个体对打搅者的打搅行为的反应主要包括:(1) 无反应和继续:被打搅个体忽略打搅者的性打搅并继续性行为;(2) 逃跑后继续:被打搅个体受到性打搅时,先逃跑避让,待性打搅停止后与原交配对象继续交配;(3) 停止交配:被打搅个体停止交配,且在观察结束前未见再次交配;(4) 选择打搅者:被打搅个体选择与打搅者交配;(5) 攻击打搅者:被打搅个体攻击打搅者 (杨斌等,2013)。
1.4 数据采集与统计分析
2021 年9 月至2022 年8 月,于每日08: 00—17: 00 (11 月至翌年3 月) 或18: 00 (4—10 月),采用焦点动物取样法 (Focal animal sampling) (Altmann,1974),以WLK-1A 群内的亚成年和成年雌性个体为目标个体,每4 d 对群内所有目标个体进行至少1 次观察、取样,每次取样10 ~ 20 min。使用SONY 数码摄像机 (HDR-CX405) 辅助记录。采用连续记录法 (continuous recording),记录目标个体交配时来自第三方的性打搅行为,包括行为发起者,行为接受者,行为发生的时间,性打搅的模式和强度,打搅者的性别、年龄和等级以及被打搅者的反应等信息。采用随机取样法 (Ad libitumsampling) 记录群内成年个体的竞争行为过程,包括冲突的时间、行为发起者、行为接受者、行为类型及行为结果等 (李保国等,2006)。
利用Microsoft Office Excel (2010) 对所采集的性打搅行为和竞争行为数据进行整理。利用IBMSPSS Statistics 26.0 对数据进行统计分析,用Origin 2021b (OriginLab Corporation) 作图。根据采集的竞争行为数据,采用David 得分法 (David´s score, DS) 分别计算成年雄性和成年雌性个体的社会等级 (Gammellet al., 2003)。采用Wilcoxon Signed Rank Test,分析太行山猕猴各性别-年龄组个体性打搅模式间和打搅对象间的差异。数据均为平均值 ± 标准误 (mean ± SE);所有检验均为双尾,显著性水平设为α = 0.05。
2 结果
2.1 性打搅行为的特点
太行山猕猴的性打搅行为仅发生在交配期(2021 年9 月至2022 年3 月),11 月是交配行为和性打搅行为的高峰期。在交配期,共记录交配行为325 次 (射精类交配占66.2%,215/325),其中34.2% (111/325) 的交配行为受到性打搅。在受打搅的交配中,记有134次性打搅行为 (表2)。其中,接触式和非接触式性打搅分别占59.7% (80/134) 和40.3% (54/134)。在非交配期 (2022 年4—8月),共观察记录交配行为10 次 (无射精),无性打搅行为发生。

表2 太行山猕猴的性打搅行为Table 2 Sexual interference behaviors in Taihangshan macaques
太行山猕猴的性打搅行为发生于交配过程的3 个阶段,以射精前阶段最多 (91.8%, 123/134),而射精阶段 (3.7%, 5/134) 和射精后阶段 (4.5%, 6/134) 均较少。根据打搅的强度和结果,射精前阶段的性打搅可分为性干扰 (87.8%, 108/123) 和性打断 (12.2%, 15/123) (表2)。
2.2 性打搅行为的发起者
大多数性打搅由单一个体发起 (80.2%, 89/111),少数由多个个体相继或同时发起 (19.8%,22/111)。在134 次性打搅行为中,雄性 (成年和亚成年) 个体发起的性打搅占29.1% (39/134),雌性 (成年和亚成年) 个体占14.9% (20/134),青少年个体占7.5% (10/134),婴幼个体占48.5%(65/134) (图1)。
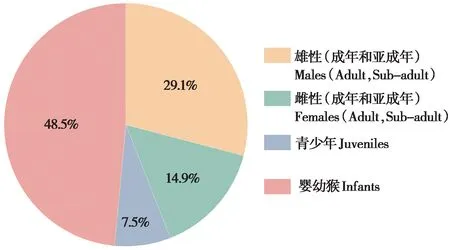
图1 太行山猕猴性打搅行为的频次分布Fig.1 Frequencies distribution of sexual interference behaviors in Taihangshan macaques
2.3 成年和亚成年雄性的性打搅
雄性 (成年和亚成年) 太行山猕猴的性打搅分为非接触式性打搅 (87.2%, 34/39) 和接触式性打搅 (12.8%, 5/39)。其中,非接触式性打搅包括追赶 (55.9%, 19/34)、趋近 (29.4%, 10/34) 和带有威胁的注视 (14.7%, 5/34);接触式性打搅包括理毛(40.0%, 2/5)、抓打 (40.0%, 2/5) 和撕咬 (20.0%,1/5)。雄性太行山猕猴的非接触式性打搅 (3.4 ±0.846,n= 10) 与接触式性打搅 (0.5 ± 0.224,n=10) 之间具有显著差异 (Z= -2.539,P= 0.011),以非接触式性打搅模式为主 (图2A)。
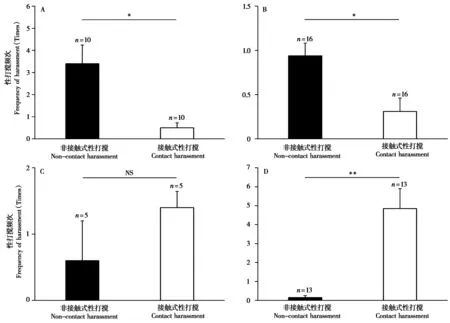
图2 太行山猕猴性别-年龄组间性打搅模式的比较. A:雄性 (成年和亚成年);B:雌性 (成年和亚成年);C:青少年;D:婴幼猴. *:P <0.05;**:P < 0.01;NS:无显著差异Fig. 2 The comparison of harassment patterns among all age-sex groups of Taihangshan macaques. A: Males (adult and sub-adult); B: Females(adult and sub-adult); C: Juveniles; D: Infants. *: P < 0.05; **: P < 0.01; NS: No significant difference
雄性太行山猕猴对交配中的雄性 (0.6 ±0.220,n= 10) 与雌性 (3.3 ± 1.000,n= 10) 的打搅频次间具有显著差异 (Z= -2.056,P= 0.040),倾向于打搅交配中的雌性 (图3A)。
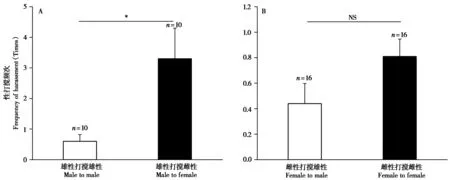
图3 雄性和雌性太行山猕猴性打搅行为的对象. A:雄性 (成年和亚成年);B:雌性 (成年和亚成年). *:P < 0.05;NS:无显著差异Fig. 3 The objects of sexual interference behaviors of male and female Taihangshan macaques. A: Males (adult and sub-adult); B: Females (adult and sub-adult). *: P < 0.05; NS: No significant difference
雄性太行山猕猴的性打搅多发生在射精前阶段 (79.5%, 31/39), 少部分发生在射精阶段(10.3%, 4/39) 和射精后阶段 (10.3%, 4/39) (表2)。发生在射精前阶段的雄性性打搅,有10 次(32.3%, 10/31) 属于性打断,并导致交配行为的终止,其余均为性干扰 (67.7%, 21/31)。这些性打断均由高等级雄性个体向雌性 (80.0%, 8/10) 或低等级雄性 (20.0%, 2/10) 发起。
对射精前阶段被打搅者反应的分析表明,被打搅者很少攻击打搅者。交配雄性对雄性打搅者的回应主要表现为逃跑后继续交配 (66.7%, 4/6) 和停止交配 (33.3%, 2/6);交配雌性对雄性打搅者的回应亦为逃跑后继续交配 (68.0%, 17/25) 和停止交配 (32.0%, 8/25)。
2.4 成年和亚成年雌性的性打搅
雌性 (成年和亚成年) 太行山猕猴的性打搅分为非接触式性打搅 (75.0%, 15/20) 和接触式性打搅 (25.0%, 5/20)。其中,非接触式性打搅包括趋近 (53.3%, 8/15)、追赶 (40.0%, 6/15) 和带有威胁的注视 (6.7%, 1/15);接触式性打搅包括理毛(80.0%, 4/5) 和挨坐 (20.0%, 1/5)。雌性打搅者的非接触式性打搅 (0.94 ± 0.143,n= 16) 与接触式性打搅 (0.31 ± 0.151,n= 16) 之间具有显著差异 (Z=-2.134,P= 0.033),以非接触式性打搅模式为主(图2B)。
雌性太行山猕猴对交配中的雄性 (0.44 ±0.157,n= 16) 与雌性 (0.81 ± 0.136,n= 16) 的打搅频次间无显著差异 (Z= -1.414,P= 0.157)(图3B)。
雌性太行山猕猴的性打搅多发生在射精前阶段 (85.0%, 17/20),少部分发生在射精阶段 (5.0%,1/20) 和射精后阶段 (10.0%, 2/20) (表2)。发生在射精前阶段的雌性性打搅,仅有4 次 (23.5%, 4/17)属于性打断,并导致交配行为的终止,其余均为性干扰 (76.5%, 13/17)。这些性打断有3 次为高等级雌性向低等级雌性发起,有1次为雌性向雄性发起,雄性的反应为选择该雌性交配。
对射精前阶段被打搅者反应的分析表明,交配雄性对雌性打搅者的回应包括无反应和继续(50.0%, 3/6)、攻击雌性打搅者 (33.3%, 2/6) 以及选择雌性打搅者 (16.7%, 1/6);交配雌性对雌性打搅者的回应包括逃跑后继续 (45.5%, 5/11)、停止交配 (27.3%, 3/11)、无反应和继续 (18.2%, 2/11)以及攻击雌性打搅者 (9.1%, 1/11)。
2.5 青少年个体的性打搅
本研究共记录到5只青少年个体的性打搅行为10 次 (表2)。其中,非接触式性打搅 (30.0%, 3/10)主要是青少年打搅者距交配对约1 m,观察交配对的交配活动 (100.0%, 3/3);接触式性打搅 (70.0%,7/10) 主要表现为挨坐并观察交配对的交配活动(85.7%, 6/7),以及钻挤 (14.3%, 1/7)。在5 只青少年打搅者 (雄性4 只,雌性1 只) 中,仅有1 只青少年雄性打搅者表现出非接触式性打搅,其余个体的性打搅均为接触式性打搅。青少年打搅者的非接触式性打搅 (0.60 ± 0.600,n= 5) 与接触式性打搅 (1.40 ± 0.245,n= 5) 之间无显著差异 (Z=-0.966,P= 0.334) (图2C)。
青少年个体的性打搅均发生在射精前阶段 (表2),打搅的对象均为交配雌性,其中有9 次性打搅为青少年个体打搅交配中的母亲,另1次为青少年个体打搅与其母亲具有友好关系的雌性。所有被打搅雌性的反应均为无反应和继续 (有时会为青少年打搅者理毛,但并不停止交配),并未发生性打断。
2.6 婴幼猴的性打搅
婴幼太行山猕猴的性打搅分为接触式性打搅(96.9%, 63/65) 和非接触式性打搅 (3.1%, 2/65)。其中,接触式性打搅主要是吮乳 (77.8%, 49/63),其次为挨坐并观察交配对的交配活动 (12.7%,8/63) 和钻挤 (9.5%, 6/63);非接触式性打搅主要是婴幼打搅者距交配对约1 m,在附近玩耍/觅食,同时观察交配对的交配活动 (100%, 2/2)。婴幼打搅者的非接触式性打搅 (0.15 ± 0.104,n= 13) 与接触式性打搅 (4.85 ± 1.055,n= 13) 之间具有显著差异 (Z= -3.192,P= 0.001),以接触式性打搅模式为主 (图2D)。此外,受婴幼猴打搅雌性 (53.9%)与未受打搅雌性怀孕率 (57.1%) 之间无显著差异(P> 0.05)。
所有婴幼猴的性打搅均发生在射精前阶段 (表2),且均为婴幼猴打搅其母亲的交配。婴幼猴的性打搅仅有1 次 (1.5%, 1/65) 表现为性打断,被打搅的母亲停止交配,并为其理毛;其余被打搅母亲的回应主要是无反应和继续 (有时会为婴幼猴打搅者理毛,但并不停止交配) (92.3%, 60/65),推开婴幼猴并继续交配 (1.5%, 1/65),有3 次是被打搅的母亲无反应,但交配雄性威胁婴幼打搅者 (4.6%,3/65)。
3 讨论
本文分析了太行山猕猴各性别-年龄组的性打搅行为的频次、方式和打搅对象等。研究发现,太行山猕猴的性打搅仅发生于交配期;各性别-年龄组的打搅频次、方式和打搅对象等均存在差异;而性打搅行为的表型差异可能源于其功能差异。
3.1 雄性的性打搅
成年和亚成年雄性太行山猕猴的性打搅多发生在射精前阶段,以非接触式性打搅模式为主,倾向于打搅交配中的雌性,且主要表现为性干扰。雄性太行山猕猴性打搅行为的动机和功能可用雄性性竞争假说来解释 (Niemeyer and Anderson,1983)。
雄性太行山猕猴以非接触式性打搅模式为主,与对藏酋猴的研究结果一致 (Panget al., 2021)。在藏酋猴中,雄性个体倾向于打搅交配中的雄性,反映出激烈的雄性性竞争 (Panget al., 2021)。雄性太行山猕猴则倾向于打搅交配中的雌性,与对猕猴 (Carpenter, 1942; Wilson, 1981; Chapais, 1983; de Elvira and Herndon, 1986; Manson, 1996) 和日本猴(Macaca fuscata) (Huffman, 1987) 的研究结果一致。研究表明,性打搅通常会破坏打搅者与被打搅者之间的关系,进而增加雄性间和雄雌间的攻击(Niemeyer and Chamove, 1983)。Chapais (1983) 认为,雄性猕猴更多地打搅交配中的雌性而非雄性是合理的,因为就雄性的能量消耗而言,打搅交配雌性成本较低。因此,雄性打搅者倾向于打搅交配中雌性,原因可能是为了减少与交配雄性的打斗冲突 (尽管交配雄性的等级通常较低),打搅交配雌性同样可以达到目的,且成本和风险较低。本研究认为,太行山猕猴雄性打搅者倾向于打搅交配中的雌性是一种低成本、低风险的性竞争策略。
本研究发现,在雄性太行山猕猴打搅的交配对中,几乎所有被打搅雄性个体的等级均低于雄性打搅者,仅观察到1次由低等级雄性个体对高等级雄性交配的打搅 (发生在交配雄性射精过程中),打搅的对象是与高等级雄性交配的雌性个体。对Cayo Santiago 猕猴的研究也有类似发现,即多数雄性打搅者的等级均高于交配雄性,而在少数由较低等级雄性发起的性打搅中,打搅对象均为交配中的雌性 (Chapais, 1983)。因此,低等级雄性太行山猕猴可能通过打搅交配中的雌性,以降低高等级雄性的交配成功率。低等级雄性藏酋猴通常通过不停地打搅高等级雄性的交配,影响其交配时间,从而降低其交配成功度 (庞奎海,2019)。
雄性太行山猕猴的性打搅多发生在射精前阶段,其结果可导致低等级雄性的交配中断,并阻止其射精。雄性的性打搅大多表现为性干扰(21/31),这至少会立即减少配偶对的共处时间,进而减少其交配频次 (Chapais, 1983)。此外,遭受持续的骚扰可能降低低等级雄性的生育能力,进而影响其繁殖成功率 (de Elvira and Herndon, 1986)。本研究还观察到1 例发生在射精后阶段的性打搅,在低等级雄性交配射精后,高等级雄性采用追赶的方式打搅交配雌性,该雌性随即选择雄性打搅者,与之交配并射精。Wilson (1981) 对猕猴的研究也发现,在少数情况下,雄性通过性打搅获得与交配雌性交配的机会。这些均为雄性性竞争假说提供了证据,类似结果亦见于藏酋猴、川金丝猴和日本猴 (Huffman, 1987, 1991; Qiet al., 2011;Panget al., 2021)。关于雄性太行山猕猴的性打搅是否能够增加其未来潜在的交配成功率,目前尚无数据支持,今后研究应予以关注。
3.2 雌性的性打搅
成年和亚成年雌性太行山猕猴的性打搅多发生在射精前阶段,以非接触式性打搅模式为主,打搅交配中的雌性与打搅交配中雄性的频次间无显著差异,且主要表现为性干扰。雌性太行山猕猴性打搅行为的动机和功能可用雌性性竞争假说解释 (Niemeyer and Anderson, 1983)。
雌性太行山猕猴以非接触式性打搅模式为主,与对藏酋猴和川金丝猴的研究结果一致 (Qiet al.,2011;庞奎海等,2020)。前人的研究发现,雌性猕猴的性打搅可以打断其他雌性的交配 (de Elvira and Herndon, 1986)。雌性太行山猕猴的性打搅多发生在射精前阶段,且可以打断正在进行的交配,从而减少交配雌性的交配成功率,这一结果支持直接的性竞争假说。太行山猕猴雌性打搅者更多地打搅交配中的雌性,但与打搅交配中雄性无显著差异。雌性针对交配雌性的性打搅均是由高等级雌性指向低等级雌性,亦有低等级雌性通过打搅交配雄性,以减少高等级雌性的交配成功率的现象。在本研究期间,观察到1次低等级雌性采用挨坐方式打搅正在与高等级雌性交配的雄性个体,导致该雄性个体停止交配,转而与打搅者交配并射精。对黑猩猩、川金丝猴和猕猴的研究也发现,在少数情况下,雌性可通过性打搅获得与交配雄性交配的机会 (Nishida, 1979; Wilson, 1981; Qiet al.,2011)。这些结果为间接的性竞争假说提供了证据。
雌性性竞争假说已在川金丝猴、狮尾狒狒(Theropithecus gelada) 和长尾叶猴 (Presbytis entellus) 等一雄多雌社会结构的灵长类物种得到验证(Dunbar, 1980; Sommer, 1989; Qiet al., 2011)。具有多雄多雌社会结构的倭黑猩猩 (Pan paniscus),其优势雌性攻击交配中的其他雌性,且雌性的性打搅几乎在所有情况下都导致交配的终止,支持雌性性竞争假说 (Vervaecke and Van Elsacker, 2000;Vervaeckeet al., 2003)。雌性猕猴的性打搅多指向交配雌性,并导致交配终止,为雌性性竞争假说提供了证据 (de Elvira and Herndon, 1986;本研究)。然而,与本研究结果不同,Wilson (1981) 对猕猴的研究发现,雌性的有效性打搅多表现为攻击交配雄性;进而认为,雌性个体间的性竞争 (可能导致繁殖成功率差异) 似乎并非猕猴所具有的特征。而且,该假说在其他具有相同社会结构 (多雄多雌型) 的猕猴属物种亦未得到验证 (Dixson,2012)。 例如红面猴 (Niemeyer and Chamove,1983)、藏酋猴 (庞奎海等,2020) 和汤基猕猴 (M.tonkeana) (Thierry, 1986),成年雌性的性打搅多发生在射精阶段和射精后阶段,一般不打断交配,与成年雌性个体间的性竞争无关,不支持雌性性竞争假说;成年雌性更倾向打搅与其具有亲缘关系或良好社会关系的雌性个体,符合占有假说。野生藏酋猴的雄雌有效性比 (性成熟雄性数量∶发情雌性数量) 接近1∶1,交配竞争对雌性来说显得不那么激烈 (庞奎海,2019),而本研究群的有效性比为0.32∶1,较少的性成熟雄性数量会引起较多发情雌性之间激烈的交配竞争。因此,雌性太行山猕猴的性打搅支持雌性性竞争假说,由此为该假说在多雄多雌社会结构的猕猴属物种提供了证据。
3.3 青少年个体的性打搅
青少年太行山猕猴的性打搅仅发生在射精前阶段,非接触式性打搅与接触式性打搅模式间无显著差异,打搅对象为其母亲或与母亲关系友好的其他雌性,且均表现为性干扰。青少年个体性打搅的动机和功能可用学习假说解释。
本研究发现,青少年太行山猕猴的性打搅行为均指向其母亲或与母亲关系友好的其他雌性,主要表现为在交配对附近观察交配,且不会打断交配,因而支持学习假说。青少年个体通过性打搅的方式欣赏和观察交配,是对社会性行为的学习,与对食蟹猴 (M.fascicularis) 和川金丝猴性打搅的研究结果一致 (de Benedictis, 1973;杨斌,2009)。对日本猴的研究也发现,雄性在整个幼年时期 (直到3岁) 都在学习并积累经验,从而逐渐获得成年个体的性行为模式 (Gunstet al., 2013)。
3.4 婴幼猴的性打搅
婴幼太行山猕猴的性打搅仅发生在射精前阶段,以接触式性打搅模式为主,打搅对象均为其母亲,且大多表现为性干扰。婴幼猴性打搅的动机和功能可用分离焦虑假说和学习假说解释。
Tutin (1979) 主张,婴幼猴的性打搅是其断乳之后缺乏安全感的一种“分离焦虑”表达。婴幼太行山猕猴的性打搅行为大多表现为钻进母亲怀里吮乳,且因可能受到交配雄性的威胁,常表现出恐惧和胆怯。本研究认为,婴幼太行山猕猴的性打搅可能是其担心与交配中的母亲的社会关系被打断,而表现出的一种情绪焦虑反应,故而支持分离焦虑假说。此外,婴幼猴也观察其母亲的交配行为。因此,婴幼猴的性打搅同样支持学习假说。
母婴冲突假说在食蟹猴得以验证 (Gore, 1986);婴幼猴仅打搅其母亲的交配,性打搅与其母亲的交配成正相关,且能够降低其母亲随后的怀孕率;而成年雌性为了避免婴幼猴的打搅,会在婴幼猴不在场的情况下交配,且在婴幼猴打搅时会对其产生负面反应。本研究发现,雌性面对婴幼打搅者的反应通常为无反应和继续 (有时会为婴幼打搅者理毛,但并不停止交配),并无负面反应;而且,受到婴幼猴打搅的雌性与未受到打搅雌性的怀孕率之间无显著差异。本研究认为,婴幼太行山猕猴的性打搅可能并不支持母婴冲突假说。此外,交配中的雄性太行山猕猴撕咬交配雌性现象很少发生,与对藏酋猴 (庞奎海,2019) 和川金丝猴(Qiet al., 2011) 的研究结果相似,均不支持保护假说。
综上所述,太行山猕猴性打搅行为的频次、方式和打搅对象等表现出性别-年龄组间差异;而各性别-年龄组性打搅行为的表型差异源于其功能差异。基于功能分析,本研究认为,雄性和雌性太行山猕猴的性打搅均支持性竞争假说,青少年个体的性打搅支持学习假说,婴幼猴的性打搅支持分离焦虑假说和学习假说。本研究结果可望增加对非人灵长类性打搅行为发育及其功能差异的认识。
致谢:感谢河南太行山猕猴国家级自然保护区济源管理局、五龙口风景管理局及匡三傲先生和匡振京先生对野外工作的支持与帮助。
猜你喜欢
保健与生活(2024年4期)2024-02-02
小猕猴学习画刊(2022年10期)2022-01-01
青年文学家(2021年29期)2021-11-19
趣味(语文)(2021年11期)2021-03-09
小猕猴智力画刊(2019年4期)2019-05-08
小猕猴智力画刊(2019年3期)2019-04-19
祝您健康(2018年1期)2018-01-10
天津诗人(2017年2期)2017-11-29
中国火炬(2014年8期)2014-07-24
中国火炬(2011年7期)2011-07-25
