
天行长臂猿鸣唱的声谱特征与性别差异
2024-02-26 10:36郭亭妍马海港韩普王子荻祝常悦楚原梦冉张利祥李如雪戚嘉儒李家华范朋飞
兽类学报 2024年1期
郭亭妍 马海港 韩普 王子荻 祝常悦, 楚原梦冉 张利祥李如雪 戚嘉儒 李家华 范朋飞*
(1 中山大学生态学院,深圳 518000)(2 中山大学生命科学学院,广州 510275)(3 云南高黎贡山国家级自然保护区保山管护局,保山 678000)(4 云南铜壁关省级自然保护区管护局,德宏 678400)(5 大理白族自治州云山生物多样性保护与研究中心,大理 671000)(6 云南高黎贡山国家级自然保护区保山管护局隆阳分局,保山 678000)
声音通讯对许多动物的生存和繁殖至关重要,能够编码物种、身份 (Terlephet al., 2015, 2016)、性别、质量等关键信息 (Charltonet al., 2009;Mathevonet al., 2010),并以此实现吸引配偶(Moos-Heilen and Sossinka, 2010)、 躲避天敌(Clarkeet al., 2006) 、防御领域 (Kinzey and Robinson, 1983)、维持社会关系 (King and Sueur, 2011)等功能。描述声音信号的基本特征是理解灵长类声音通讯行为与开展保护研究的基础 (Bernsteinet al., 2016; Fanet al., 2022),不仅有助于理解信号编码信息的类别、实现功能的过程以及人类语言的起源与进化,且有助于在保护区通过声学技术监测濒危灵长类种群动态等保护实践 (Yanget al.,2020; Vu and Doherty, 2021; Lwinet al., 2022)。
长臂猿科 (Hylobatidae) 动物是一类小型树栖类人猿,主要分布于东南亚热带雨林与季雨林中,包括长臂猿属 (Hylobates)、白眉长臂猿属 (Hoolock)、冠长臂猿属 (Nomascus) 和合趾猿属 (Symphalangus) (Liebalet al., 2014; Reichardet al.,2016)。其配偶制主要为一夫一妻制 (Jianget al.,1999; Fuentes, 2000)。近年的研究发现,在冠长臂猿属的几个物种中可以形成稳定的一夫二妻制(Guanet al., 2018),在长臂猿属和合趾猿属的少数种群中可以形成稳定的一妻二夫群 (Reichardet al., 2016)。鸣唱行为 (morning song) 是长臂猿最典型的行为之一,鸣唱声音洪亮且传播距离远 (Marshall and Marshall, 1976)。在鸣唱中,长臂猿可以发出由多种音节 (在吸气或呼气时发出的一个单独连续的不同频率或调频的声音) 组成的复杂音句(phrase) 或序列 (sequence) (指一个大的或松散的音节的集合,可能一起发出或单独发出) (Haimoff,1983a),由多个序列进一步组合成鸣唱回合(bout) ,这种复杂的鸣唱结构在所有类人猿中绝无仅有 (Geissmann, 2000, 2002)。除了克氏长臂猿(Hylobates klossii) 和爪哇长臂猿 (Hylobates moloch) 外 (Haimoff, 1985; Dooleyet al., 2012),所有长臂猿都具有雌雄精确配合发出的二重唱或三重唱行为 (Geissmann, 2002; Maet al., 2021),且雌性都会发出激动鸣叫 (great call,仅由雌性发出的固定刻板而又易于识别的声音) (Reichardet al.,2016)。研究发现鸣唱能够传递鸣叫者的位置、性别、身份等信息 (Fenget al., 2014; Hamet al.,2016; Hradecet al., 2023),经野外行为观察或声学回放实验证实具有防御资源 (配偶或食物) (Fanet al., 2009; Niuet al., 2023)、凝聚家庭关系等功能(Maet al., 2021)。
现生白眉长臂猿属包括东白眉长臂猿 (Hoolock leuconedys)、西白眉长臂猿 (Hoolock hoolock) 以及天行长臂猿 (Hoolock tianxing) 3 个物种,仅分布于中国云南西部和藏南地区,以及缅甸、孟加拉和印度东北部,均形成稳定的一夫一妻制家庭群 (Choudhury, 1991; Yuanet al., 2013)。研究表明该属雌雄叫声差异小,两性具有相同的音节类型,没有性别特异性 (Marler and Tenaza,1976; Marshall and Marshall, 1976),音节类型主要有eek、oo、wa 和ow 音节 (Marler and Tenaza,1976; Haimoff, 1983a, 1983b; Gittins and Tilson,1984)。白眉长臂猿属的雌雄二重唱被认为由引入序列 (introductory sequence)、间奏序列 (interlude sequence) 以及激动鸣叫组成 (Haimoff, 1983a),其中音节组合等特点可能在不同物种、不同家庭群之间存在高度变异 (Marler and Tenaza, 1976; Gittins and Tilson, 1984)。然而,现有的大多数对白眉长臂猿属鸣唱的研究仅基于少数圈养个体,且缺少对声谱特征的量化分析。
天行长臂猿于2017 年由我国科学家命名 (Fanet al., 2017),是我国一级重点保护野生动物,被IUCN 红色名录列为濒危物种 (Fanet al., 2020)。天行长臂猿雌雄叫声十分相似,且二重唱中两性的叫声高度重叠,目前尚无系统的研究揭示该物种叫声的基本特征,包括雌雄独猿与家庭群基本的音节、音句类型以及雌雄叫声的差异等。这阻碍了对天行长臂猿的日常监测、保护与行为学研究。本研究使用手持录音机与被动录音机录制音频,测量音节的时间与频率指标,利用无监督机器学习的方法对音节聚类,确定天行长臂猿的音节类型,对比雌性和雄性音节的频率特征差异;进一步结合音句的音节组成以及音句的长度划分出音句类型,对比雌性独猿、雄性独猿及合唱中雌性与雄性音句使用频率的差异,以期为日常巡护监测中对天行长臂猿个体的性别识别提供科学依据,并为未来研究天行长臂猿鸣唱的功能和信息编码打下基础。
1 研究方法
1.1 研究地点
研究地点位于云南省高黎贡山国家级自然保护区 (北纬24°56′ ~ 28°23′,东经98°08′~ 98°53′),以及云南省德宏傣族景颇族自治州盈江县苏典傈僳族乡 (北纬25°00′35″ ~ 25°16′25″,东经97°45′31″ ~ 98°02′21″)、卡场镇 (北纬24°51′11″ ~ 25°06′01″,东经97°42′57″ ~ 97°53′59″) 以及芒章乡(北纬24°51′38″ ~ 25°02′08″,东经98°02′43″ ~ 98°14′26″)。
1.2 数据收集
本研究于2018—2023 年收集了天行长臂猿鸣唱回合。使用便携式手持录音机SONY PCM-D100与ZOOM H6 录音机,及Sennheiser ME 66 指向性话筒近距离对不同家庭群和独猿进行录音,同时记录鸣唱类型 (独唱或二重唱)、频次 (数量 / 天)、鸣唱开始和结束时间、鸣唱树位置以及群体特征(个体数量和形态) 等信息。此外,根据现有天行长臂猿家庭群或独猿已知分布和潜在分布的范围,在卡场、芒章乡分别布设了6 台、33 台自动录音机 (Song Meter SM4),且尽可能保证仅录制一个家庭群或者独猿的鸣声,避免后期无法确定同一段声音数据中是否包含多个家庭群。前期研究发现天行长臂猿主要在清晨鸣叫,极少数在12: 00后鸣叫 (Yinet al., 2016; Chanet al., 2017),因此所有自动录音机设置为仅在每天06: 00—13: 00 连续录音。手持录音机设置采样率为48 kHz,音频采样精度为24 bit,自动录音机设置采样率为24 kHz,音频采样精度为16 bit。我们还结合保护区的种群调查结果来确认声音数据对应的个体信息 (表1)。
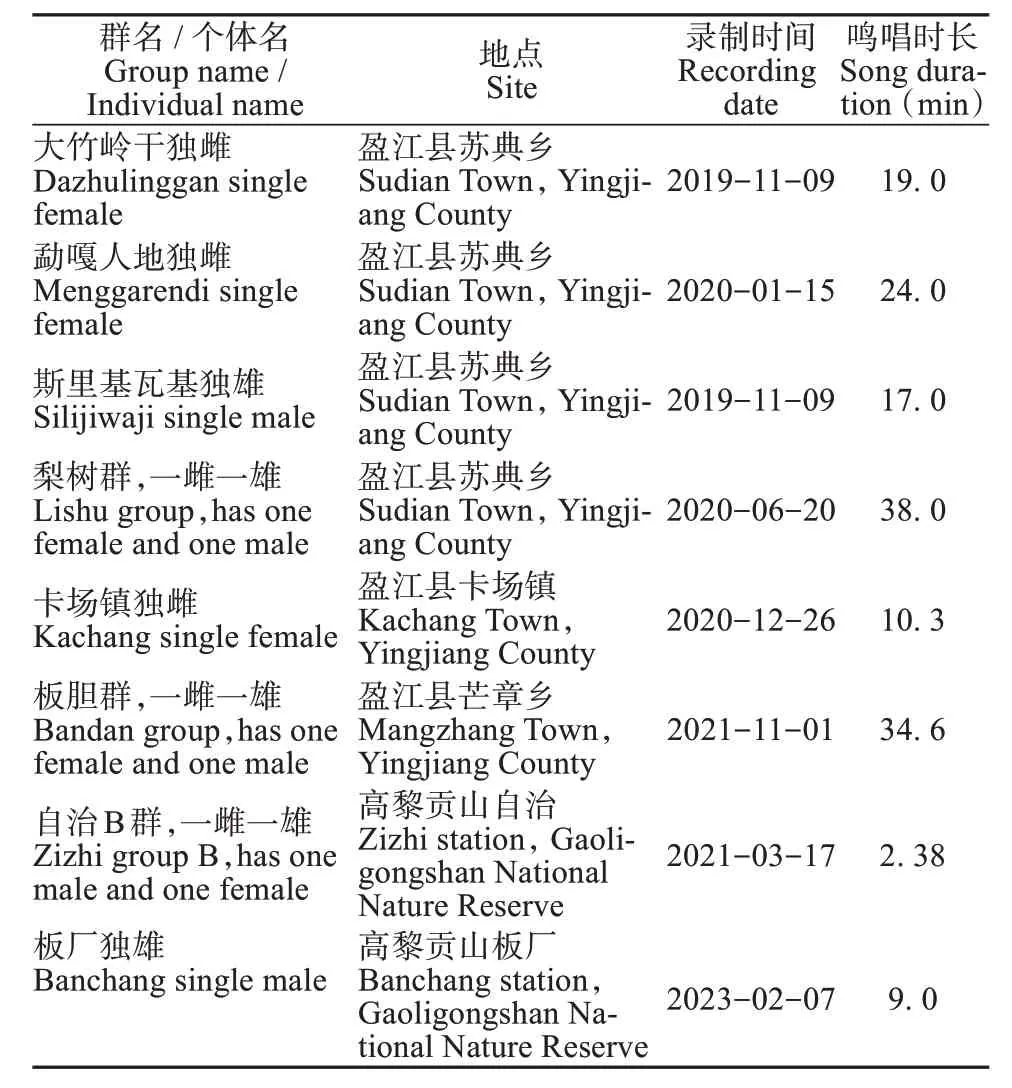
表1 云南高黎贡山和盈江天行长臂猿鸣唱数据收集信息表Table1 Song bout information of Hoolock tianxing recorded in Mt.Gaoligong and Yingjiang County, Yunnan, China
1.3 音节的时间标注和频率测量
依据激动鸣叫完整、背景噪声低、尽可能覆盖多只个体的原则,使用Praat (× 64) 6.2.12 测量了3个家庭群、3只雌性独猿、2只雄性独猿的8段高质量鸣唱录音。对于合唱回合,基于前人文献总结的雌雄配合规律 (Haimoff, 1983a; Gittins and Tilson, 1984),标注能够识别雌雄个体的音节。首先,使用Adobe Audition 针对鸣唱回合提取左声道并标准化,随后使用Praat (× 64) 6.2.12 的语音标注功能 (annotation function) 生成每个回合的声谱图 (参数设置:快速傅里叶变换时加海宁窗,窗长为1 024 个点,50%重叠,带宽为0.03 s) 并标注,以此获得每个音节的持续时间;其次,使用Praat(× 64) 6.2.12 的查看和编辑功能 (View & Edit) 针对每个音节测量起始频率、结束频率、最小频率和最大频率4 个基频 (f0) 指标 (参数设置:频率范围为0 ~ 2 000 Hz,动态范围为70 db,带宽为0.03 s)。另外,于2021 年使用录像设备Nikon P900、SONY FDR-AX700 4K 近距离拍摄9 段板厂家庭群的清晰视频,观察个体在发出各个音节时是否存在呼吸状态的切换。
1.4 音节的分类
1.4.1 时间、频率信息的标注
基于测量的频率数据,计算音节频率的极差R (音节的最高频率与最低频率的差值)、频率的落差start_end (起始频率与结束频率的差值)、斜率dfslope [(结束频率 - 起始频率) / 时间] 以及频率变化速率dfrange [(最大频率 - 最小频率) / 时间],最终构建包含10个天行长臂猿鸣唱指标的数据集,并以此进行分析。
1.4.2 k-中心点聚类 (k-medoids 聚类) 及其效果验证
k-medoids 聚类是一种无监督机器学习方法,该方法在迭代中通过最小化每个聚类簇中的对象与该簇的中心点的距离来优化新的聚类中心(Schubert and Rousseeuw, 2021),对于噪声和孤立点不敏感。为了降低量纲不同带来的影响,对数据集中心化与标准化处理 [(数值 - 均值) / 标准差] (Kassambara, 2011)。之后使用k-medoids 聚类对标准化后的数据进行聚类,然后使用R 包cluster中的pam函数完成音节类型的聚类。
基于人工标注的时间和频率信息,在R包中完成数据分析步骤,使用clValid 包中的连接度 (connectivity)、轮廓系数 (silhouette coefficient)、Dunn指数、非重叠的平均比例 (the average proportion of non-overlap, APN)、平均距离 (the average distance,AD)、均值间的平均距离 (the average distance between means, ADM)、品质因数 (the figure of merit,FOM) 来衡量聚类效果 (表2),连接度、APN、AD、FOM 值越小聚类效果越好,轮廓系数、Dunn指数值越大聚类效果越好。

表2 本研究中使用的聚类验证指标含义Table 2 The meaning of cluster validation metrics used in this study
将聚类数目从2 (最小分类数目) 每隔一个取至10 (超过了可能存在的所有过渡类型) (2, 3, 4……10),将轮廓系数、Dunn 指数从小到大排列,连接度、APN、AD、ADM、FOM 指数从大到小排列,利用综合评价中的秩和比的方法,计算每一组合在每一指标中排序的均值 (Sun and Tian,1994),选出最佳聚类数。基于划分出的音节类型,计算声谱图特征的均值与标准差,声谱图的绘制基于R 包seewave 的spectro 函数实现。
1.4.3 识别音节层面上的性别差异
对每种音节的声谱图特征分别做wilcoxon 秩和检验,为了降低由假设检验次数带来的假阳性率,使用ggpubr 包的compare_means 函数计算经Bonferroni 校正后的P值,比较雌性和雄性之间是否有音节上的差异。
1.5 音句的分类及性别差异
根据音句中音节的组合、音句的长度以及前人的文献将天行长臂猿的音句划分为7种类型。统计每一种音句的平均音节个数,使用R 包igraph绘制网络图展示音句中音节的转换,并以R 包seewave的spectro函数绘制音句声谱图。
在音句的使用方面,将所有的回合 (bout) 划分为雌性独唱、雄性独唱、合唱中雌性鸣唱部分、合唱中雄性鸣唱部分4个类别,计算每一种音句在不同类别中出现的平均频次 (出现总次数 / 该类型的回合数量),由于类别内的样本量不同,对每一个类别做标准化处理,用R 包pheatmap 的pheatmap函数绘制热图,并使用UPGMA法对类别进行层级聚类,从而比较在合唱、独唱中雌雄两性音句使用的差异。为展示合唱中雌性与雄性鸣唱的差异,使用Adobe Photoshop 2020 以不同的颜色标示出合唱声谱图中雌性和雄性分别贡献的部分。
2 结果
2.1 音节类型的识别
共测量了8 段高质量鸣唱录音中3 812 个音节的特征 (表3),用于k-medoids 聚类。k-medoids 聚类在识别天行长臂猿的声音类型中有较好的效果,综合7个聚类验证的指标,将天行长臂猿的音节类型分为5 类 (表4),且5 种音句类型在发声时不连续或存在呼吸状态的切换。5 类音节中wa 音节为1 022个,ow音节为933个,whoop音节为910个,oo 音节为792 个,eek 音节为155 个。雌性和雄性鸣唱包含的音节类型相同,且合唱与独唱时的音节类型也相同 (表3)。基于划分出的5类音节类型,计算声谱图特征的均值与标准差 (表5)。
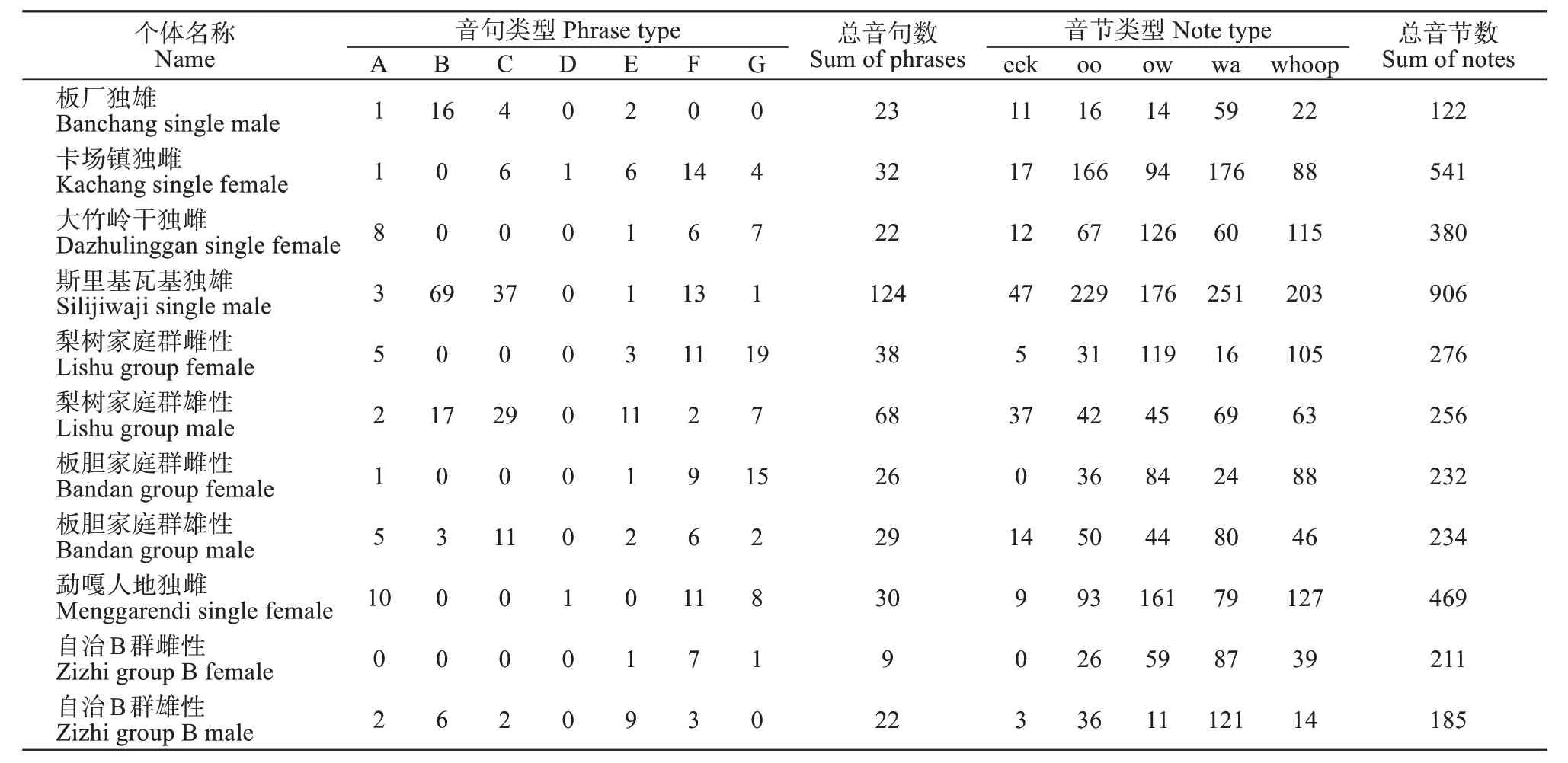
表3 天行长臂猿不同个体音节、音句标注信息统计Table 3 Statistic for the number of notes and phrases of each individual of Hoolock tianxing

表4 不同簇数的k-medoids聚类结果的验证指标Table 4 Validation index of k-medoids clustering with different cluster numbers
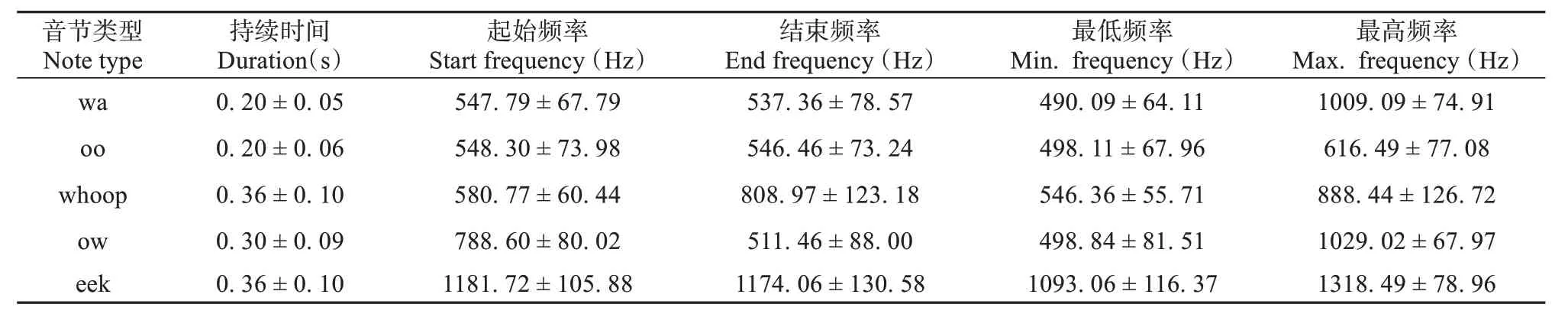
表5 声音类型的声谱图特征 (仅列举持续时间、起始频率、结束频率、最低频率、最高频率)Table 5 The spectrogram features of notes (only list duration, start frequency, end frequency, minimum frequency, maximum frequency)
wa 音节 (图1a、f):频率先上升再下降,持续时间为 (0.20 ± 0.05) s,是雌雄性使用最频繁的音节类型。oo 音节 (图1b、g):起始频率、结束频率和最低频率几乎没有差别,最高频率稍高,通常与wa 音节组合出现在音句中,但在音句加速 (音节的持续时间缩短) 时通常省去oo 音节。whoop 音节 (图1c、h):在音句中,该音节常由oo音节转变而来,或逐渐过渡为oo 音节,通常与wa 音节组合使用。ow 音节 (图1d、i):在音句中ow 音节通常与whoop 音节组合,音节频率略微上升,频率稳定一段时间后下调。节奏加快时常过渡为wa音节,起始频率相比于wa音节更高。eek 音节 (图1e、j):频率最高,持续时间为 (0.36 ± 0.10) s,eek 音节的起始频率、结束频率、最低频率近似,在雌性和雄性的独唱与合唱中均有出现,通常出现在独唱的一个句子的开头,或者在合唱时雌性或雄性某方开始鸣唱的开头。

图1 天行长臂猿雌雄性各音节的声谱图. a、f:雄性与雌性的wa 音节;b、g:雄性与雌性的oo 音节;c、h:雄性与雌性的whoop 音节;d、i:雄性与雌性的ow音节;e、j:雄性与雌性的eek音节Fig. 1 Spectrograms of notes produced by male and female of Hoolock tianxing. a, f: spectrograms of ‘wa’ note produced by male and female; b,g: spectrograms of ‘oo’ note produced by male and female; c, h: spectrograms of ‘whoop’ note produced by male and female; d, i: spectrograms of‘ow’ note produced by male and female; e, j: spectrograms of ‘eek’ note produced by male and female
2.2 雌雄音节类型的声谱特征对比
在ow 音节、wa 音节中雌性频率变化的极差、频率落差、频率变化速率都显著大于雄性个体(P< 0.01),而雌性的whoop 音节的频率变化幅度与速率显著低于雄性个体 (P< 0.01)。除极差外,雌性的oo 音节各个频率指标都略低于雄性 (P<0.02),雌性与雄性的eek 音节的起始频率、极差、最大频率无显著差异,其他频率指标具有显著差异 (P< 0.05) (图2)。
2.3 天行长臂猿独唱时的音句类型与雌雄差异
依据对独猿音频声谱图的观察、音节类型与音句长度,结合前人文献,将天行长臂猿的音句类型整理分类为7 种 (图3,图4),分别为音句A:由wa-oo/ow oo或连续的wa组成,能单独出现或连续叫2 ~ 3声;音句B:wa-whoop或ow-whoop,常常紧接着雄性的eek音节 (图5);音句C:由1 ~ 5个wa-oo 与wa-whoop 组合而成,通常会和音句B 以多样的方式组合;音句D:由连续的oo 音节组成;音句E:完全由wa 音节以及wa-oo 组成;音句F:由whoop-ow 与wa-oo 组合而成,平均音节个数最多;音句G:主要由2 ~ 10个连续的whoop-ow/owwhoop 组成。在雌性独唱仅未出现音句B,雄性独唱仅未出现音句D (图5)。

图3 天行长臂猿音句的声谱图Fig.3 Spectrograms of Hoolock tianxing’s phrases

图4 天行长臂猿音句的句法网络图Fig. 4 Network of notes in Hoolock tianxing’s phrases

图5 各类型的音句平均出现频次 (出现次数/回合数) 热图Fig.5 Heatmap of the mean occurence frequency of phrases (occurrence frequency/number of bouts)
2.4 天行长臂猿的合唱模式
天行长臂猿的合唱由引入序列、激动鸣叫序列和间奏序列组成,图6 是一个天行长臂猿二重唱的典型例子。引入序列通常由离散的音节以及音句A 与音句D 组成。天行长臂猿合唱有时会省略引入序列、直接进入激动鸣叫。激动鸣叫序列通常由雌性发起,雄性配合,主要由音句E、F 组成,激动鸣叫中的音句F 音节数量多且变化大 (音节数为23.34 ± 12.91)。而间奏序列中,雄性鸣唱的部分由音句B 与音句C 组合而成,雌性鸣唱的部分为较短的音句G (音节数为7.32 ± 3.43)。
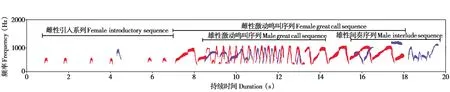
图6 天行长臂猿二重唱的声谱图,红色代表雌性贡献的部分,蓝色代表雄性贡献的部分Fig. 6 The spectrogram of Hoolock tianxing’s duet,the female contribution is shown in red and the male contribution is shown in blue
合唱与独唱、雌性鸣唱与雄性鸣唱都具有音句A、E、F、G,合唱中雌性的部分未出现音句B、C、D。层次聚类的结果表明,在音句平均出现频率上,雄性独唱和二重唱中雄性鸣唱的部分相似,雌性独唱和二重唱中雌性鸣唱的部分相似 (图5)。
3 讨论
本文分析了天行长臂猿鸣唱的声谱特征与性别差异。天行长臂猿的鸣唱模式与已发表的同属物种西白眉长臂猿、东白眉长臂猿的鸣唱模式类似,鸣叫回合均由引入序列、激动鸣叫序列、间奏序列组成 (Haimoff, 1983b; Geissmann, 2002)。结果表明,通过量化与无监督机器学习的方式,可以很好地将容易被忽视与混淆的音节类型区分开。从音节的声谱特征上看,本研究识别出天行长臂猿具有5 种基本的音节类型,其中4 种在已发表的西白眉长臂猿与东白眉长臂猿的声谱图中都曾出现 (Marler and Tenaza, 1976; Marshall and Marshall,1976; Haimoff, 1983b),但前人的研究没有识别whoop 音节。whoop 音节经常逐渐转换成oo 音节,缺少量化分析时,这一音节可能容易与oo 音节混淆,因此从未得到分类与描述。根据近距离拍摄的清晰视频,whoop 音节与ow 音节之间虽然缺少时间间隔,但是在发声时存在换气,因此whoop应被划分为一个独立的音节。
天行长臂猿声音的性二型较弱,雌雄使用同样的声音曲目,这可能是一夫一妻的婚配制度、种群小、资源竞争较弱的副产物 (Aunget al.,2023)。在音节特征上,雌性的oo 音节、eek 音节和whoop 音节的频率指标都低于雄性,这可能是由雌雄体型的略微差异造成,白眉长臂猿雌性个体体重略高于雄性 (Reichardet al., 2016),声带略长于雄性,基频更低 (Taylor and Reby, 2010)。在雌雄性wa音节与ow音节的频率指标中,仅雌性的最大频率高于雄性,可能在听觉上能够避免在激动鸣叫中与雄性的声音混淆。
过去的研究普遍认为白眉长臂猿属下的物种雌雄两性间完全没有声音上的性二型,因而怀疑在家庭群合唱时雌雄性可能无法分别向雌雄性别的独猿传递自身适合度的信息 (Haimoff, 1983b),且因难以识别异性声音信号导致独猿个体无法根据鸣声寻找潜在配偶。但本研究结果表明,尽管具有相同的声音类型,但雌雄两性间在音节特征和音句使用上存在差异。
在音句的使用上,天行长臂猿雌雄两性可能都能鸣唱同样的音句,尽管音句B在本研究的雌性鸣唱中并未出现,但曾出现在同属的西白眉长臂猿的雌性独猿的鸣唱中 (Haimoff, 1983b),天行长臂猿雌性可能也能够鸣唱该类型的音句。雌性和雄性在音句的鸣唱频次上存在差别,在合唱的间奏序列中,与前人研究相似,雄性更多鸣唱音句B (wa-whoop) 与音句C (wa-oo-wa-whoop),而雌性更多鸣唱音句G (whoop-ow-whoop-ow-whoop-ow)。本研究对雌雄独猿鸣唱的描述补充了前人研究的空白,在雌雄二重唱中,雄性和雌性都更倾向于独唱时的音句类型,即雄性极少唱音句G,雌性极少唱音句B 和C。而音句B、C、G 在二重唱中多出现在雌雄重叠程度较弱的间奏序列中,可能合唱的间奏序列能够分别编码并传递该家庭群中雌雄个体的身份、质量信息。而在激动鸣叫部分,雌雄两性的音句E、F 高度重叠,可能能够作为整体实现传递雌雄配合强度的功能。
以往的研究仅基于雌雄之间是否存在特异性的音节来判定白眉长臂猿属声音二型性的强弱,这可能阻碍了对白眉长臂猿属鸣叫的功能与信息编码的理解。研究证明灵长类动物能够识别到音节上的差异与音节组合的不同 (Waneliket al.,2013),从而识别自己的亲属 (Rendallet al., 1996)、提取捕猎相关的信息 (Clarkeet al., 2006; Waneliket al., 2013)。天行长臂猿在音节特征和音句使用上的差异能否作为一种个体身份信息同样被信号接收者解译,在未来还需要通过回放实验验证。
历史上,天行长臂猿曾广泛分布于我国云南省的多个市县,包括腾冲、保山等 (李致祥和林正玉,1983;马世来和王应祥,1986,1988)。2017年的调查发现天行长臂猿仅分布在云南省腾冲市、保山市和盈江县15 个片区 ,约32 ~ 34 群和11 个成年独猿,总数不足150 只 (Yanget al., 2020;Zhanget al., 2020)。天行长臂猿的生存面临着巨大的挑战,很多长臂猿生活在片段化的森林斑块中,有些斑块甚至仅有1群长臂猿。这些孤立群体中的成年后代无法找到配偶进行繁殖,限制了长臂猿种群恢复 (Fanet al., 2011)。由于独猿能通过鸣唱吸引配偶 (Clarkeet al., 2006),因而回放天行长臂猿独猿个体的鸣唱可能能够促进其在片段化森林斑块找到异性并组成家庭、繁殖后代。而识别雌雄声音差异能够保证日常巡护监测中对天行长臂猿个体性别识别的时效性与准确性,从而为后续的种群调查、定向回放吸引和个体识别工作建立基础,助力对物种的保护与监测。未来的研究需侧重建立天行长臂猿的个体识别技术体系,而天行长臂猿声音的信息编码机制与功能则需要通过回放实验等手段进一步研究。
致谢:感谢云南高黎贡山国家级自然保护区保山管护局、云南高黎贡山国家级自然保护区保山管护局隆阳分局、云南高黎贡山国家级自然保护区保山管护局腾冲分局、德宏州林业和草原局、云南铜壁关省级自然保护区、盈江县林业和草原局、苏典傈僳族乡人民政府、大理白族自治州云山生物多样性保护与研究中心。感谢赵超先生提供的视频资料。
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23
小哥白尼(野生动物)(2022年7期)2022-09-20
云南画报(2021年7期)2021-08-06
杂文月刊(选刊版)(2021年5期)2021-06-02
杂文月刊(选刊版)(2020年9期)2020-10-28
杂文月刊(2019年18期)2019-12-04
杂文月刊(2019年19期)2019-12-04
青年歌声(2017年11期)2017-03-15
Coco薇(2015年12期)2015-12-10
食品科学(2013年17期)2013-03-11
