
气候对不同地区川金丝猴出生模式的影响
2024-02-26 10:36陆展滔兰官伟李瑞源司旭蕊刘星宇莫诚姚辉齐敦武向左甫
兽类学报 2024年1期
陆展滔 兰官伟 李瑞源 司旭蕊 刘星宇莫诚 姚辉 齐敦武 向左甫
(1 中南林业科技大学生命科学与技术学院,长沙 410004)(2 成都大熊猫繁育研究基地,四川省濒危野生动物保护生物学重点实验室,成都 610086)(3 西华师范大学,西南野生动植物资源保护教育部重点实验室,南充 637002)(4 四川白河国家级保护区,九寨沟 623400)(5 湖北神农架金丝猴保育生物学湖北省重点实验室,神农架 442411)(6 中南林业科技大学林学院,长沙 410004)
关于非人灵长类动物出生模式的描述首次出现在猴和猿类的年生殖周期研究中 (Lancaster and Lee, 1965),被划分为3 种,即显著的季节性出生、高峰性出生、无规律性出生(Struhsaker and Leland,1986)。季节性出生被定义为出生事件只发生在明确划定的时期内 (van Schaiket al., 1999),如云南拉沙山滇金丝猴 (Rhinopithecus bieti) 社群的出生事件只发生在2—4 月,表现出严格的出生季节性(Huanget al., 2012;王双金等,2012);猕猴 (Macaca mulatta) 74%的出生事件集中在5—6 月,表现为高峰性出生 (Vandenbergh and Vessey, 1968);某些物种则几乎全年都存在出生情况,如野生鬃毛吼猴 (Alouatta palliata) 全年的大部分月份均有新生婴猴被记录,出生季节分布没有显著差异,表现为无规律性出生 (Glander, 1980);有些物种则表现出出生高峰期不明显或者较弱,如郁乌叶猴(Trachypithecus obscurus);而有些物种则表现出非季节性的繁殖模式,如红吼猴 (Alouatta seniculus)每月都有婴猴出生,但是存在一个出生低谷期(Crockett and Rudran, 1987)。
影响非人灵长类动物出生模式的因素有很多,栖息地环境温度可以影响动物出生模式,因为温度与动物的能量消耗和体温调节有关。对于恒温动物而言,在过高或过低的温度条件下,需要消耗能量来维持体温的恒定,从而减少自身在繁殖上所花费的能量。随着温度上升,圈养松鼠猴(Saimiri sciureus) 每月出生婴猴数量增加 (Trevino,2007);阿根廷查科地区的夜猴 (Aotus azarai) 每年4—5 月进行交配 (Fernandez-Duqueet al., 2002),以避免极端高温给受孕个体带来的能量代谢挑战;降水量也会对动物的繁殖产生影响,如栖息于热带地区的北绒毛蛛猴 (Brachyteles hypoxanthus),每年的气温变化幅度小,其出生季节受降水影响大,雨季延长会导致出生季节的跨度延长 (Wiederholt and Post, 2011);黑猩猩 (Pan troglodytes) 在每年雨季 (食物供应高峰) 开始发情,降水量的季节性变化通过影响食物供应对其繁殖起间接作用 (Brockman and van Schaik, 2005; Andersonet al., 2005)。
动物繁殖期消耗能量巨大,其将繁殖周期中最需要能量的时期与每年食物供应的高峰时期相匹配,因此繁殖与栖息环境食物供应的波动关联很大 (Di Bitetti and Janson, 2000; Bronson, 2009)。基于食物供应与动物季节性繁殖之间的联系,可以划分为收入繁殖者 (income breeder) 与资本繁殖者(capital breeder) 两种不同的繁殖类型 (Di Bitetti and Janson, 2000; Brockman and van Schaik, 2005)。资本繁殖者为了生殖活动而提前储存能量,为随后的怀孕、分娩、育幼做准备,如马达加斯加西北部狐猴 (Lemuridae) 会在雨季提高食物摄入量、增加体重和怀孕,而出生季节则集中在食物缺乏的旱季 (Pichon and Simmen, 2015);收入繁殖者则根据当前的食物摄入量开始繁殖活动,如栖息在刚果盆地的山魈 (Mandrillus sphinx) 在食物匮乏的季节 (旱季) 受孕,其出生高峰期与食物丰富的季节 (雨季) 相符,在哺乳期的前半段,雌性山魈摄入食物以满足婴猴的营养需求,在哺乳期的后半段,婴猴可以独立进食 (Dezeureet al., 2022)。美国洛杉矶动物园中的皇柽柳猴 (Saguinus imperator) 几乎在所有月份都有婴猴出生,园内有超过七成的雌性在3—7 月分娩,而野生的皇柽柳猴种群的出生高峰却出现在4—8 月,出生季节比圈养种群晚,可能的原因是圈养种群拥有稳定的食物供应 (Baker and Woods, 1992)。
海拔梯度的增加导致气温降低,甚至高海拔地区的最低气温可能会达到寒温带地区常见的水平,而降水量在一定范围内随着海拔上升而增加,通常在连续海拔梯度的中等高度位置达到高峰(van Schaik and Brockman, 2005),导致植被类型沿海拔梯度变化垂直分布,一般来说,高海拔地区与低海拔或中等海拔地区相比,其植物丰富度和初级生产力相对较低 (Rahbek, 1995),因此海拔梯度引起的极端气候与食物供应的变化影响灵长类动物的繁殖,这一观点在对藏酋猴 (Macaca thibetana)(Zhao and Deng, 1988; Zhao, 1994)、黑猩猩 (Pan troglodytes schweinfurthii) (Matthewset al., 2021),以及大型食草动物如马鹿 (Cervus canadensis)(Moyeset al., 2011)、 狍 (Capreolus pygargus)(Peláezet al., 2020) 和大角羊 (Ovis canadensis)(Festa-Bianchet, 1988) 的研究中得到证实。
纬度位置也可以用于解释灵长类动物出生模式的变化。通常纬度越高其出生季节性越强,因为与靠近赤道的地区相比,高纬度地区的气候与食物供应呈明显的季节性变化 (Janson and Verdolin, 2005; Heldstabet al., 2020)。例如西藏小昌都(北纬29°15′,东经98°37′) 和云南拉沙山 (北纬26°20′,东经99°15′) 的野生滇金丝猴群出生同步模式不同,前者出生季节第2周出生数量最多,后者第1、4周出生数量最多 (Xiang and Sayers, 2009;王双金等,2012)。Trebouet 等(2021) 基于13 个野生猕猴属 (Macaca) 种群出生模式的调查发现,随着纬度的上升,猴群出生模式趋向于严格的季节性,其中北纬12°以上的种群呈现严格的季节性繁殖,北纬3° ~ 14°和南纬5°范围内存在中度季节性繁殖种,南纬1° ~ 北纬3°之间为非季节性繁殖种。
灵长类的繁殖行为也受到光周期的影响,光照诱导褪黑激素释放,进而调节下丘脑-垂体-性腺 (Hypothalamic-pituitary-gonadal axis) 轴系统复杂的神经内分泌过程,来调控动物的繁殖 (Bradshaw and Holzapfel, 2007; Dardente, 2012)。例如Fernandez-Duque等(2002) 研究发现,每年4—5月,光照时间缩短刺激阿根廷夜猴 (Aotus azarai) 交配活动的开始。光周期通常由纬度决定,Rasmussen(1985) 发现生活在不同纬度区域的狐猴 (Lemuridae),其交配季节开始日期不同,高纬度种群的交配与出生季节开始日期晚,因此通过比较分布于不同纬度的灵长类种群的出生模式可以探究光周期对繁殖的影响。
川金丝猴 (Rhinopithecus roxellana) 属于我国一级重点保护野生动物,分类上隶属于灵长目 (Primates) 猴科 (Cercopithecidae) 疣猴亚科 (Colobinae)仰鼻猴属 (Rhinopithecus),是我国特有的珍稀濒危灵长类物种,分布于四川、甘肃、陕西及湖北,栖息于海拔1 000 ~ 4 100 m的温带高山、亚高山森林 (向左甫,2020)。川金丝猴是季节性繁殖的灵长类动物 (Qiet al., 1995; Xianget al., 2017),有研究发现圈养川金丝猴出生高峰期在3—6 月 (Zhanget al., 2000; Renet al., 2003),但野生川金丝猴多数集中于3 月下旬至5 月上旬生育 (Xianget al.,2017),目前对川金丝猴出生季节性的研究较多,但是鲜有将两地猴群的出生情况进行对比,以探讨气候等因素对川金丝猴出生季节的影响。
本文报道了2023 年1—8 月四川白河国家级保护区王家山地区和湖北神农架大龙潭区域川金丝猴投食群的婴猴出生情况,分析了两个种群出生季节性以及气候对婴猴出生事件的潜在影响。
1 研究方法
1.1 研究地点与对象
研究地点为四川白河国家级自然保护区和湖北神农架国家公园。白河国家级自然保护区 (北纬33°10′ ~ 33°22′,东经104°01′ ~ 104°12′) 位于四川省阿坝藏族羌族自治州九寨沟县,保护区地处岷山山系北段,海拔1 240 ~ 4 453 m,属温带半湿润气候 (李艳忠等,2016;金贵祥等,2020),猴群位于王家山区域 (北纬104°08′,东经33°15′,海拔1 700 ~ 2 200 m),主要植被类型包括常绿阔叶林、落叶阔叶林、针叶阔叶林和高山针叶林 (彭振中等,2022)。
神农架国家公园大龙潭川金丝猴野外科研基地 (北纬31°29′,东经110°117′) 位于湖北省西北部的神农架国家公园大龙潭区域内 (北纬31°21′ ~ 31°36′,东经110°03′ ~ 110°33′),该保护区地处大巴山系,海拔约2 200 m (Yaoet al., 2011),地跨中、北亚热带,春季短暂 (4—5 月底),而冬季漫长(11 至翌年3 月底),全年气候温暖湿润,季节性变化明显,属于亚热带季风气候 (朱兆泉和宋朝枢,1999)。
研究对象为白河王家山及神农架大龙潭区域的两个习惯化了的川金丝猴种群。2023年8月,白河种群个体总数约130 只,由7 个一雄多雌社会单元 (one-male muit, OMU) 和1 个全雄社会单元 (allmale muit, AMU) 组成;神农架种群个体总数约90 只,由6 个OMU 和1 个AMU 组成;白河种群成年雌性个体数量为45 只,神农架种群成年雌性个体数量为32 只,所有的成年雌性个体均被观察到参与交配活动;OMU 单元成年雄雌比,白河种群约为1∶6.4,神农架种群约为1∶5.3 (表1)。两地工人每日在固定地点投喂食物2次,猴群中的所有个体,包括成年、亚成年、青少年和婴幼猴,均可通过面部特征 (五官形状、皱纹多寡)、受伤情况 (伤疤形状、身体残疾) 和毛发特征等进行个体识别。两猴群已经习惯研究人员的存在,可在离猴群5 ~ 20 m的距离内观察猴群。

表1 2023年白河种群、神农架种群一雄多雌单元基本结构Table 1 Basic structure of one-male unit in Baihe and Shennongjia groups in 2023
1.2 数据收集
2023 年1—8 月,两名研究人员在白河种群活动范围内进行跟踪观察,当发现刚出生的婴猴时,采用焦点动物取样法 (focal animal sampling method) 记录婴猴出生日期、婴猴性别、雌性个体信息,并用单反相机 (日本,佳能700D) 进行拍照记录,在整个观察期间,对出生婴猴数量及成年雌性数量进行统计;神农架猴群,相同的信息由大龙潭基地研究人员提供。并对白河保护区王家山区域及神农架大龙潭区域气候信息 (气温、降水)进行统计,资料来源于欧洲中期天气中心 (http//www.ecmwf.int)。
1.3 数据分析
根据Eisenberg 等 (1981) 的方法,以观察期内第一个婴猴出生日期为起始,将猴群的出生季节每7 天作为一个周期进行划分,计算出生季节的婴猴平均出生日期、出生日期中值和标准差,以及周婴猴 / 成雌值 (I / F),其中I 为当前婴猴出生总数,F为观察期内成年雌性总数。根据van Schaik 等(1999) 的定义,3个月内发生2/3的出生事件,可以认为该物种具有出生季节性。按照Caughley (1977)的方法分析猴群出生季节性是否符合脉冲模型,即如果一个种群出生季节标准差小于30 d,那么此物种可被视作季节性出生的物种。计算公式为:
公式中f为每周期婴猴出生数量,x为周期序号,L为观察期内婴猴出生总数达到一半时所在周期的上限日期前的累积天数,g为观察期内婴猴出生总数的一半与观察期内婴猴出生总数达到一半时所在周期前的婴猴出生个体数的差,C 为周期天数(7天),d为中值所在周期的婴猴出生数量。
将2023 年1—7 月的气候数据 (日平均气温、日降水量),按照上述方法进行划分,计算每个周期的平均气温和累积降水量,使用SPSS 26.0 数据分析,使用Circular statistics 和Rayleigh test (Z =nr2) 分析出生季节性和出生模式 (王双金等,2012),采用Hierarchical cluster analysis分析出生高峰期,通过Pearson相关性检验 (PCCs) 分析周平均气温、降水量与周I / F之间的相关性,显著性水平设置在α = 0.05。
2 结果
2.1 出生季节性
白河种群出生日期为1月3日至8月20日,跨度为232 d,除6月外,其他月均有婴猴出生,其首例婴猴出生事件于2023 年1 月3 日被研究人员记录。2月28日至3月20日、3月28日至4月25日、5月23日至6月6日,周I / F斜率增加,说明婴猴出生数量增加,婴猴数在8月20日达到最大值 (17个) (图1a)。
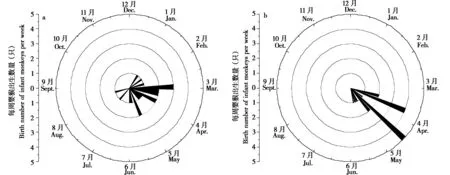
图1 白河种群 (a)、神农架种群 (b) 猴群婴猴出生分布Fig 1 Birth distribution of monkeys in Baihe group (a) and Shennongjia group (b)
神农架种群出生日期为3 月17 日至5 月23 日,跨度为75 d。猴群中新生婴猴于3 月、4 月和5 月被记录,在3月17日发现首个新生婴猴。3月10—30 日、4 月6—17 日,周I / F 斜率增加,说明婴猴出生数量增加,婴猴数在5 月10 日达到最大值(14个) (图1b)。
白河种群每周出生婴猴数量较为平均,第33周后出生事件结束,两个组的出生呈同步 (图2a);神农架种群第2 周 (3 月24—30 日) 与第5 周 (4 月14—20 日) 出生婴猴较多,占52%,其他周出生婴猴较少,第10周后出生事件结束 (图2b)。
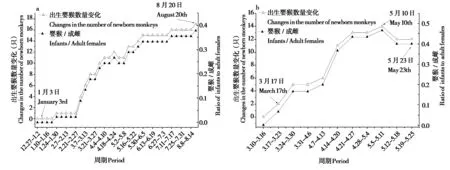
图2 白河种群 (a)、神农架种群 (b) 猴群婴猴 / 成雌比 (I / F) 和出生婴猴数量变化Fig. 2 Changes of infants / adult females ratio (I / F) and number of newborn monkeys in Baihe group (a) and Shennongjia group (b)
白河种群猴群出生季节跨度大,平均出生日期为5 月6 日 (SD = 141 d),出生日期中值为4 月3 日,不符合脉冲模型,出生季节性不严格,但是有75%的出生事件发生在3—5 月,有2 例发生在1月,各有1例发生在2月、7月和8月,因此白河种群猴群出生模式也为季节性繁殖 (r= 0.681,Z=9.749,P< 0.01),存在出生高峰期 (高峰:3 月1—16 日,次高峰:4 月2—18 日,末高峰:5 月10—30 日) (图2a);神农架种群的平均出生日期为4 月28 日 (SD = 19 d),出生日期中值为4 月14 日,符合脉冲模型,存在两个出生高峰期 (高峰:3 月17—27 日,次高峰:4 月5—26 日),且该组猴群所有的出生事件发生在3—5 月,该组猴群出生模式表现出严格的季节性 (r= 0.975,Z=16.168,P< 0.01) (图2b)。
2.2 气候季节性变化与婴猴出生
2020年8月至2023年7月,白河种群区域与神农架种群区域月平均气温有显著差异 (t= -2.96,P< 0.01),总体上白河种群区域月平均气温低于神农架种群区域 (图3),但月降水量差异不明显(t= -0.98,P= 0.37)。白河种群区域年平均气温为(5.38 ± 0.51) ℃,最低月平均气温为 (-8.16 ±1.16) ℃ (1 月),最高月平均气温为 (16.57 ±0.28) ℃ (7月),年降水量 (1259.71 ± 173.36) mm,4—10 月总降水量占全年的85% ~ 89%;神农架种群区域年平均气温为 (11.17 ± 0.13) ℃,最低月平均气温为 (-0.96 ± 0.42) ℃ (1 月),最高月平均气温为 (21.07 ± 0.68) ℃ (7 月),年降水量为(1470.71 ± 60.46) mm,4—10月总降水量占全年的81% ~ 84%。
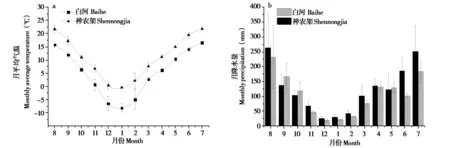
图3 白河和神农架月平均温度及月降水量比较Fig. 3 Comparison of monthly average temperature and monthly precipitation in Baihe and Shennongjia
婴猴出生事件受气温、降水量的影响,婴猴出生事件发生在平均气温上升与降水量增加的月份 (表2),分析表明白河种群区域周平均气温 (rs=0.97,P< 0.01)、周降水量 (rs= 0.65,P< 0.01) 与I / F变化呈显著正相关;神农架种群区域周平均气温与I / F变化呈正相关 (rs= 0.71,P< 0.05),但周降水量与I / F 变化相关性不明显 (rs= 0.31,P=0.35) (图4)。

图4 白河种群 (a、b) 及神农架种群猴群 (c、d) 周平均气温、周降水量与婴猴 / 成雌值 (I / F) 的关系Fig. 4 The relationship between the weekly average temperature, weekly precipitation and infant / adult female values (I / F) in the Baihe group (a,b) and Shennongjia group (c, d)
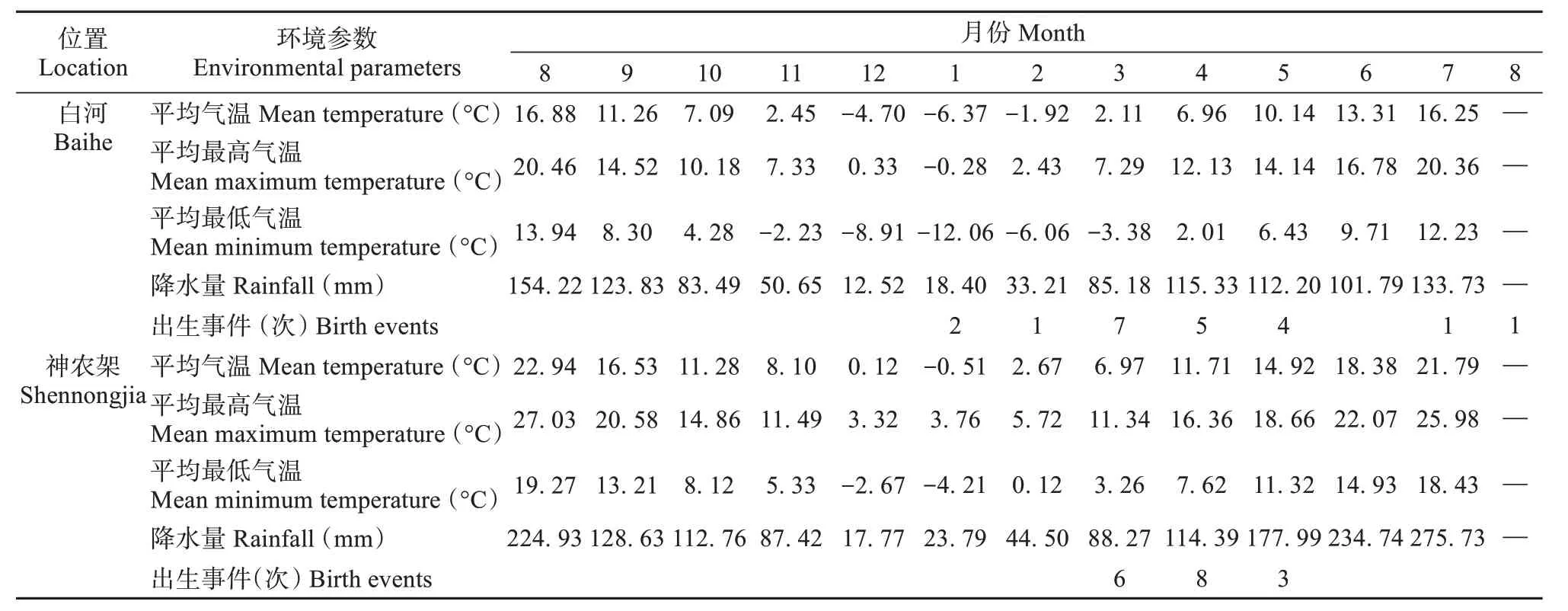
表2 2022年8月至2023年8月白河种群、神农架种群栖息地气候季节性变化与出生事件Table 2 Seasonal changes of habitat climate and birth events in Baihe and Shennongjia groups from August 2022 to August 2023
3 讨论
根据2023 年1—8 月的白河种群与神农架种群婴猴出生数据特征,我们发现神农架种群猴群的出生日期标准差小于30 d,所有的出生事件发生在3—5 月,表现出严格的出生季节性;而白河种群猴群出生日期标准差大于30 d,75%的出生事件发生在3—5月,1月有2例,2月、7月和8月各有1例,因此白河种群猴群出生模式为高峰性出生,不符合脉冲模型,并没有表现出像神农架种群的高度季节性。
虽然两个种群出生分布格局存在差异,但是其出生事件发生于气温与降水量逐渐上升的月份,并表现出相同的出生高峰期 (3—5 月)。研究发现白河种群与神农架种群周I/F 变化和周平均气温均呈正相关,表明气温是灵长类动物开启季节性繁殖的机制,其主要决定了动物4 个能量消耗的过程,即基础代谢率、主动代谢、生长和繁殖,当环境温度远低于动物正常体温水平时,需要消耗一定的能量来维持体温,以保证其他生命活动的热需求 (Dunbar, 1988),因此栖息地温度变化与动物的体温调节成本息息相关。本研究中两地猴群所在栖息地环境温度变化幅度大,神农架种群猴群栖息地7 月平均温度为21.87 ℃,1 月平均温度为-0.51 ℃;白河种群猴群栖息地7 月平均温度为16.57 ℃,1 月平均温度为-6.37 ℃,均相差约23 ℃,温度变化大使怀孕的雌猴面临更大的能量压力与体温调节压力 (Xianget al., 2017)。同时环境温度也跟分娩和婴猴的生长密切相关,猴类独特盆骨结构使得产道比猿类狭窄,分娩时间更长,此时雌猴与婴猴面临的体温调节压力大 (Trevathan, 2015)。相关研究表明,南非萨马拉雌性青腹绿猴 (Chlorocebus pygerythrus) 选择在不活动的夜间进行分娩,避免了分娩后婴猴由于羊水蒸发而导致的体表温度降低的情况 (McFarlandet al.,2022)。也有研究指出,刚出生的豚尾猴 (Macaca nemestrina) 幼崽易受较低环境温度的影响 (Ruppenthalet al., 1983),长期的低温条件使婴猴将更多的能量用来调节体温,从而发育延缓 (Lubachet al., 1992),因此怀孕雌猴会选择于分娩时间和婴猴生长有利的环境温度条件,减少低温带来的风险。
在周I/F值变化和每周降水量的相关性检验中,白河种群呈正相关,神农架种群相关性不明显,因此本研究的数据不能验证每周降水量与猴群每周I/F 值变化之间的联系。在过去关于热带灵长类动物出生季节性的研究中会使用出生时间与降水量之间的相关性 (Andersonet al., 2005; Wiederholt and Post, 2011),因为热带地区每年的月平均气温变化幅度小,而月降水总量表现出季节性的变化,这样的变化对热带灵长类动物的繁殖影响显著。川金丝猴属于温带物种,温带地区月平均气温变化幅度大,尽管本研究中两组猴群栖息地的降水模式也呈现出季节性的变化,但极端温度带来的热应激对温带物种的繁殖影响可能更大。
除了要面对环境温度带来的体温调节压力以外,栖息环境的食物供应季节性变化也会对动物的繁殖周期产生影响 (Dunbar, 1988; Di Bitetti and Janson, 2000),因为许多灵长类动物具有较长的妊娠期和哺乳期,它们在繁殖上要耗费巨大的能量成本,如苏门答腊岛上的长尾猕猴 (Macaca fascicularis),需要通过协调哺乳期的能量需求高峰与食物供应的季节性高峰,或者利用食物供应高峰期来开启新一轮的繁殖活动 (van Schaik and van Noordwijk, 1985)。本研究中两地猴群出生高峰期均为旱季末期和雨季早期 (3—5 月),在每年的3 月,降水量逐渐上升,此时栖息地植物嫩芽与嫩叶开始出现,这为已分娩的雌猴提供了丰富蛋白补充,满足婴猴在食物匮乏时期的营养与生长需求;5 月嫩叶开始长出,6—10 月是食物供应的高峰期,此时婴猴可以开始进食水分高、易消化的嫩叶 (Li, 2006;铁军等,2010,2011;李沂韦,2016)。一些研究表明,在笼养与野生条件下,成年雌性川金丝猴个体会选择食物供应较高的季节开始交配行为 (任宝平,2002;李文俊,2010;魏力,2016),由此可以看出川金丝猴符合资本繁殖者的特点,即成年雌性在食物供应高的季节通过摄入食物、积累营养来繁殖后代 (Xianget al.,2017)。
白河种群的出生模式与已知的野生川金丝猴种群出生模式有所差异 (任宝平,2002;Xianget al., 2017),野生种群的生育事件集中在每年的3月底至6 月初发生,本研究发现白河种群除3—5 月外,在温度较低的1—2 月及温度较高的7—8 月仍有婴猴出生,因此环境温度不太可能是影响猴群出生时间跨度的因素。圈养的川金丝猴种群 (1—6 月) 和滇金丝猴种群 (12 月至翌年6 月) 的繁殖季节及出生事件与白河的川金丝猴种群具有类似表现 (Cuiet al., 2006; Xianget al., 2017)。考虑到白河国家级自然保护区于2020 年开始投食,投食历史很短 (投食起始时间由保护区提供),人工投食可能是白河种群出生时间跨度长 (232 d) 的原因,因为与自然生态条件下的可获取的食物相比,稳定的人工投食为雌猴在食物匮乏时期提供丰富的营养条件,促进了繁殖活动的进行 (Altman and Alberts, 2003)。
神农架国家公园于2006年开始投食计划 (投食起始时间由保护区提供),投食历史比白河种群悠久,相比于栖息地食物供应,神农架种群更依赖于稳定的人工投食,且两地猴群出生高峰期 (3—5 月) 与食物供应高峰期 (6—10 月) 一致,由此推测在食物供应条件有差异的情况下,两地猴群依然表现出一样的出生高峰期。川金丝猴可能通过感知环境因素的变化 (气温、降水),来调节繁殖活动的时间。这与生活在巴西的普通狨猴 (Callithrix jacchus) 的研究结果相似,即人工圈养的普通狨猴种群在有恒定的食物供应的条件下,依然表现出与野生种群相似的繁殖模式 (Sousaet al., 1999)。
白河种群平均出生日期为5 月6 日,出生日期中值为4 月3 日,而神农架种群平均出生日期为4月28日,出生日期中值为4月14日。白河种群栖息地纬度比神农架种群高2° ~ 3°,海拔相差不大,然而高纬度的白河种群比神农架种群提前73 d 出生,平均出生日期推迟9 d,出生日期中值提前11 d,出生时间跨度相差116 d,且白河种群和神农架种群每周出生的婴猴数量不同,同步性模式不同,白河种群每周出生数量较为均匀,存在3个出生高峰期,而神农架种群在第2、5周出生较多,存在两个出生高峰期。可以看出两地猴群的出生模式趋异适应,即随着栖息地纬度的升高,猴群出生时间提前,但出生时间跨度变长,出生季节性减弱,这一现象与前人的研究结果相比存在差异(Fooden and Aimi, 2003; Bicca-Marques and Gomes, 2010;王双金等,2012; Trebouetet al.,2021)。西藏小昌都滇金丝猴群与云南拉沙山滇金丝猴群栖息地纬度相差3°,前者比后者更早开始进入出生季节,同时也表现出更显著的出生季节性 (王双金等,2012)。位于不同纬度地带的两群日本猴 (Macaca fuscata),高纬度 (北纬38°17′) 的猴群出生季节比低纬度 (北纬31°22′) 提前,因为高纬度地区冬季食物供应较差,出生季节的提前可以让婴猴在冬季来临前达到良好的发育水平,以适应恶劣的环境条件 (Fooden and Aimi, 2003)。巴西黑角悬猴 (Cebus apella) 出生季节性研究发现,分布于热带与亚热带地区的3个猴群的出生分布格局没有差异,可能的原因是栖息地环境温度与光周期差异变化不大 (Bicca-Marques and Gomes, 2010)。秘鲁野生皇柽柳猴种群,雌性大多数在10 月和11 月产仔,而北半球的同种野生种群出生高峰期为4—8 月,其出生模式发生了季节性逆转 (Terborgh, 1984; Baker and Woods, 1992)。因此纬度变化给猴群出生季节性带来的影响仍存在许多复杂的问题,需要更系统的研究来解释其调控机制。
本研究支持川金丝猴属于季节性繁殖物种 (出生高峰期为3—5 月),神农架种群出生模式属于季节性出生而白河种群更偏向于高峰性出生。野外观察发现的白河川金丝猴种群出生季节数据比其他地方报道的更早,可能原因是白河种群猴群栖息地纬度比神农架种群高,冬季来临时间提前,环境温度更低,选择更早时间的繁殖策略可能是为了让婴猴更早地过渡到可以独立进食的阶段,以减少怀孕母猴在食物供应较低的冬季的能量挑战,这体现了川金丝猴对极端环境的适应性,同时在人工投食带来营养丰富的食物条件下,雌性个体更容易达到合适的繁殖状态,投食历史短暂的白河种群表现出更长的出生时间跨度。
猜你喜欢
人民周刊(2020年15期)2020-08-31
环球时报(2020-07-22)2020-07-22
参花·青春文学(2019年4期)2019-09-10
小学阅读指南·低年级版(2019年3期)2019-05-13
当代陕西(2018年20期)2018-10-31
中国周刊(2018年6期)2018-06-15
当代陕西(2018年6期)2018-05-22
中国水土保持(2018年5期)2018-05-10
语文教学与研究(读写天地)(2015年4期)2015-05-25
小猕猴智力画刊(2013年1期)2013-03-15
