
饲料中添加发酵豆粕对湘云鲫生长性能、肠道及肝脏健康的影响
2024-02-05 08:04周良星刘佳裕冯麒凤周桂莲刘天骥张遨然王光花顾夕章王向荣李洪琴
中国饲料 2024年3期
周良星, 赵 敏, 刘佳裕, 冯麒凤 , 周桂莲, 李 勇,5, 刘天骥,刘 匆, 张遨然, 王光花, 罗 浩, 顾夕章, 王向荣, 罗 莉 , 李洪琴
(1.新希望六和股份有限公司农业农村部饲料及畜禽产品质量安全控制重点实验室,北京 100102;2.西南大学水产学院重庆市水产科学重点实验室淡水鱼类资源与生殖发育教育部重点实验室,重庆 400715;3.射阳六和饲料有限公司江苏省水产生物环保饲料工程研究中心,江苏 盐城 224000;4.广州希望饲料有限公司,广东 广州 510000;5.山东新希望六和集团有限公司青岛市动物饲料安全重点实验室,山东 青岛 266061;6. 中国海洋大学,山东 青岛 266072)
鱼粉供应不足,价格居高不下,鱼粉替代技术研究已成为水产养殖领域热点问题 (Dossou 等,2018;Van Doan 等,2018;Hoseinifar 等,2018)。 豆粕蛋白质含量高、价格低且氨基酸分布相对平衡,是鱼粉主要替代蛋白源之一(Gatlin III 等,2007),但豆粕中的抗营养因子会损伤水生动物肠道健康, 降低水生动物的生长性能 (Li 等,2017 ,Gemede 和Ratta,2014), 限制了其在水产饲料中的应用。 微生物发酵可最大限度消除豆粕中的抗营养因子,增加豆粕中粗蛋白质、小肽和游离氨基酸含量,并产生寡肽、谷氨酸、益生菌、维生素、乳酸、 未知生长因子等生物活性物质 (张蓓莉等,2017;Mukherjee 等,2016), 显示出比豆粕更好的鱼粉替代效应(李宁宇等,2020;Wang 等,2016)。近年来,在大口黑鲈(何明等,2020)、石斑鱼(罗智等,2004)、凡纳滨对虾(刘韬等,2022)和异育银鲫(陈萱等,2005)等水产养殖品种上的研究表明,以适宜比例发酵豆粕替代饲料中的鱼粉, 可以提高动物消化酶活性、抗氧化能力以及免疫功能,改善肌肉品质, 且不会对养殖动物生长性能产生不利影响。
湘云鲫是由二倍体母本和异源四倍体父本杂交培育而来的三配体淡水新品种,具有食性广、生长迅速、抗逆性强、耐低氧、耐低温等优点,是目前国内个体最大的优质鲫鱼,市场潜力巨大,然而目前鲜有发酵豆粕在湘云鲫饲料中的应用效果报道。基于此,本试验以发酵豆粕替代湘云鲫饲料中的部分鱼粉和豆粕,研究其对湘云鲫生长性能、肠道及肝脏健康的影响, 为发酵豆粕在湘云鲫饲料中的应用提供理论依据。
1 材料和方法
1.1 试验饲料 以鱼粉、肉粉、发酵豆粕、豆粕等为蛋白源, 大豆油为脂肪源配制成2 组等氮(37%)等脂(7.8%)的试验饲料,对照组为鱼粉组(FM 组), 添加发酵豆粕的试验饲料为发酵豆粕替代组(FSM 组)。 为避免限制性氨基酸差异对试验结果的影响,FSM 组添加0.02%的蛋氨酸和0.10%的赖氨酸,使其蛋氨酸及赖氨酸含量与FM组近似相等。饲料组成及主要营养成分见表1。饲料原料均购于重庆新希望饲料有限公司, 粉碎后过60 目筛,用转鼓式混合机混合均匀,制成直径2.5 mm 的饲料,风干后-20 ℃保存。

表1 试验饲料组成及主要营养成分(风干基础) %
1.2 试验鱼饲养与管理 试验鱼购于重庆市北碚歇马渔场,用1%食盐消毒后,以商品饲料暂养1 周。 选择体质健康、个体均匀、体重为(37.26±0.16)g 的试验鱼160 尾, 随机置于8 个圆柱形水族缸(容积500 L)中,分为2 组,每组4 个重复,每个缸20 尾。
每天8:30、12:30、17:00 饱食投喂3 次,养殖周期56 d。 养殖水源为曝气自来水,饲养期间水温22 ~26 ℃, 溶解氧>6.0 mg/L,pH 6.5 ~7.5,氨氮含量<0.50 mg/L,亚硝酸盐含量<0.05 mg/L。
1.3 样品采集 养殖试验结束后, 停食24 h,MS-222(0.01%)麻醉,分别计数、称重。 每重复随机取5 尾鱼测定体重、体长,从尾静脉抽取静脉血于EDTA-Na 抗凝管中,静置后4000 g 离心10 min,取上层血浆存于-80 ℃冰箱中保存待用。 每重复随机取5 尾鱼,于冰盘中解剖,分离肠道、肝脏、肌肉,液氮速冻后于-80 ℃保存备用。 此外,每重复随机取3 尾鱼,取前肠、肝脏于4%多聚甲醛中固定备用,用于石蜡切片的制备。
1.4 指标测定
1.4.1 生长指标及形体指标测定 采用以下公式计算生长性能和生物学性状:
增重率(WG)/%=(W2-W1)/W1×100;
特定生长率 (SGR)/(%/d)=(LnW2-LnW1)/t×100;
饵料系数(FCR)=D/(W2-W1);
干物质沉积率(DRR)/%=100×(W2×W2d-W1×W1d)/(Wd×Wdd);
蛋白质沉积率 (PRR)/%=100×(W2×W2p-W1×W1p)/(Wd×Wdp);
脂肪沉积率(LRR)/%=100×(W2×W2l-W1×W1l)/(Wd×Wdl);
肥满度(CF)/(g/cm3)=100×W2/L3;
脏体比(VSI)/%=内脏团重/W2×100;
肝体比(HSI)/%=肝脏重/W2×100。
式中:W1为起始均重,g;W2为终末均重,g;Wd为尾均摄食饲料总量,g;W2d为终末鱼全鱼干物质含量;W2p为终末鱼全鱼粗蛋白质含量;W2l为终末鱼全鱼粗脂肪含量;Wdd为饲料干物质含量;Wdp 为饲料粗蛋白质含量;Wdl为饲料粗脂肪含量;W1d为初始鱼干物质含量;W1p为初始鱼粗蛋白质含量;W1l为初始鱼粗脂肪含量;L 为体长,cm;D 为尾均摄食量,g;t 为饲喂天数。
1.4.2 试验饲料、全鱼、肌肉常规营养成分的测定水分、 粗蛋白质、 粗脂肪、 粗灰分含量分别采用105 ℃烘箱干燥法(GB/T 6435-2006)、凯氏定氮法(GB/T 6432-1994)、索氏抽提法(GB/T 6433-1994)、550℃灼烧法(GB/T 6438-1992)进行测定。
1.4.3 石蜡组织切片制样 多聚甲醛固定后,乙醇梯度脱水,置于二甲苯中透明,用低熔点的石蜡包埋成蜡块, 用Leica UC7 RT 切片机连续切片,厚度为4 μm,展片后HE 染色,中性树胶封片。
1.4.4 酶活及生化指标的测定 消化酶: 淀粉酶、胰蛋白酶、脂肪酶、Na+-K+-ATPase 活性,抗氧化指标:总抗氧化能力(T-AOC),总超氧化物歧化酶(T-SOD)活性、丙二醛(MDA)含量,血浆生化指标:谷丙转氨酶(ALT)活性、谷草转氨酶(AST)活性、甘油三酯(TG)含量、总胆固醇(TCHO)含量、高密度脂蛋白胆固醇(HDL-C)含量、低密度脂蛋白胆固醇(LDL-C)含量、溶菌酶(LMZ)活性,均采用南京建成生物工程研究所试剂盒测定,免疫球蛋白M(IgM)含量、二胺氧化酶(DAO)活性、内毒素(LPS)含量指标采用上海优选Elisa 试剂盒测定。
1.5 数据分析 采用SPSS 22.0 对数据采用独立样本T 检验比较分析组间差异, 显著水平为P<0.05。 试验结果以“平均值±标准差”(Mean±SD)形式表示。
2 结果
2.1 发酵豆粕对湘云鲫生长性能的影响 由表2可知,FM 组与FSM 组间湘云鲫的终末均重、增重率、特定生长率、尾均摄食量、饵料系数、肥满度、脏体比、肝体比、干物质沉积率、蛋白质沉积率和脂肪沉积率均无显著差异(P>0.05)。

表2 发酵豆粕对湘云鲫生长性能的影响
2.2 发酵豆粕对湘云鲫体组成的影响 由表3可知,与FM 组相比,FSM 组湘云鲫肌肉粗蛋白质含量显著提高(P<0.05), 粗灰分含量显著降低(P<0.05)。FM 组与FSM 组间湘云鲫全鱼粗蛋白质、粗脂肪、粗灰分以及肌肉粗脂肪含量均无显著差异(P>0.05)。
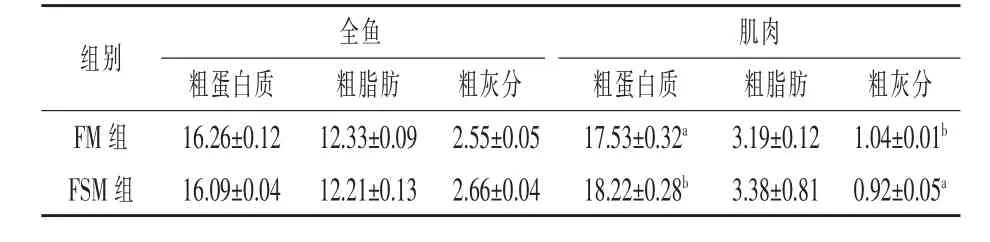
表3 发酵豆粕对湘云鲫体组成的影响(鲜样) %
2.3 发酵豆粕对湘云鲫肠道健康的影响
2.3.1 发酵豆粕对湘云鲫肠道形态结构的影响由图1 可观察到两组肠道绒毛结构均完整, 无绒毛脱落现象。由表4 可知,FM 组与FSM 组肠道绒毛数目与高度均无显著差异(P>0.05)。

图1 发酵豆粕替代鱼粉对湘云鲫肠道形态结构的影响

表4 发酵豆粕对湘云鲫肠道形态结构的影响
2.3.2 发酵豆粕对湘云鲫肠道黏膜通透性的影响发酵豆粕对湘云鲫肠道形态结构的影响 由表5可知, 与FM 组相比,FSM 组血浆D-乳酸含量显著降低了21.5%(P<0.05),而二胺氧化酶活性与内毒素含量无显著差异(P>0.05)。

表5 发酵豆粕对湘云鲫肠道黏膜通透性的影响
2.3.3 发酵豆粕对湘云鲫肠道消化吸收功能的影响 由表6 可知,与FM 组相比,FSM 组肠道脂肪酶活性显著提高了10.2%(P<0.05),然而胰蛋白酶、淀粉酶和Na+-K+-ATP 酶活性与FM 组间无显著差异(P>0.05)。
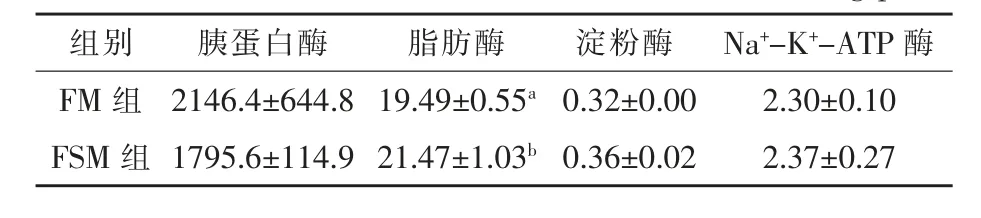
表6 发酵豆粕对湘云鲫肠道消化吸收功能的影响 U/mg prot
2.3.4 发酵豆粕对湘云鲫肠道抗氧化能力的影响由表7 可知,FSM 组肠道总超氧化物歧化酶活性和丙二醛含量与FM 组间无显著差异(P>0.05)。

表7 发酵豆粕对湘云鲫肠道抗氧化能力的影响
2.4 发酵豆粕替代鱼粉对湘云鲫肝脏结构功能的影响
2.4.1 发酵豆粕替代鱼粉对湘云鲫肝脏形态结构的影响 发酵豆粕对湘云鲫肝脏结构影响见肝脏图2。 FM 组和FSM 组肝细胞索明显, 排列较整齐,肝细胞结构完整。
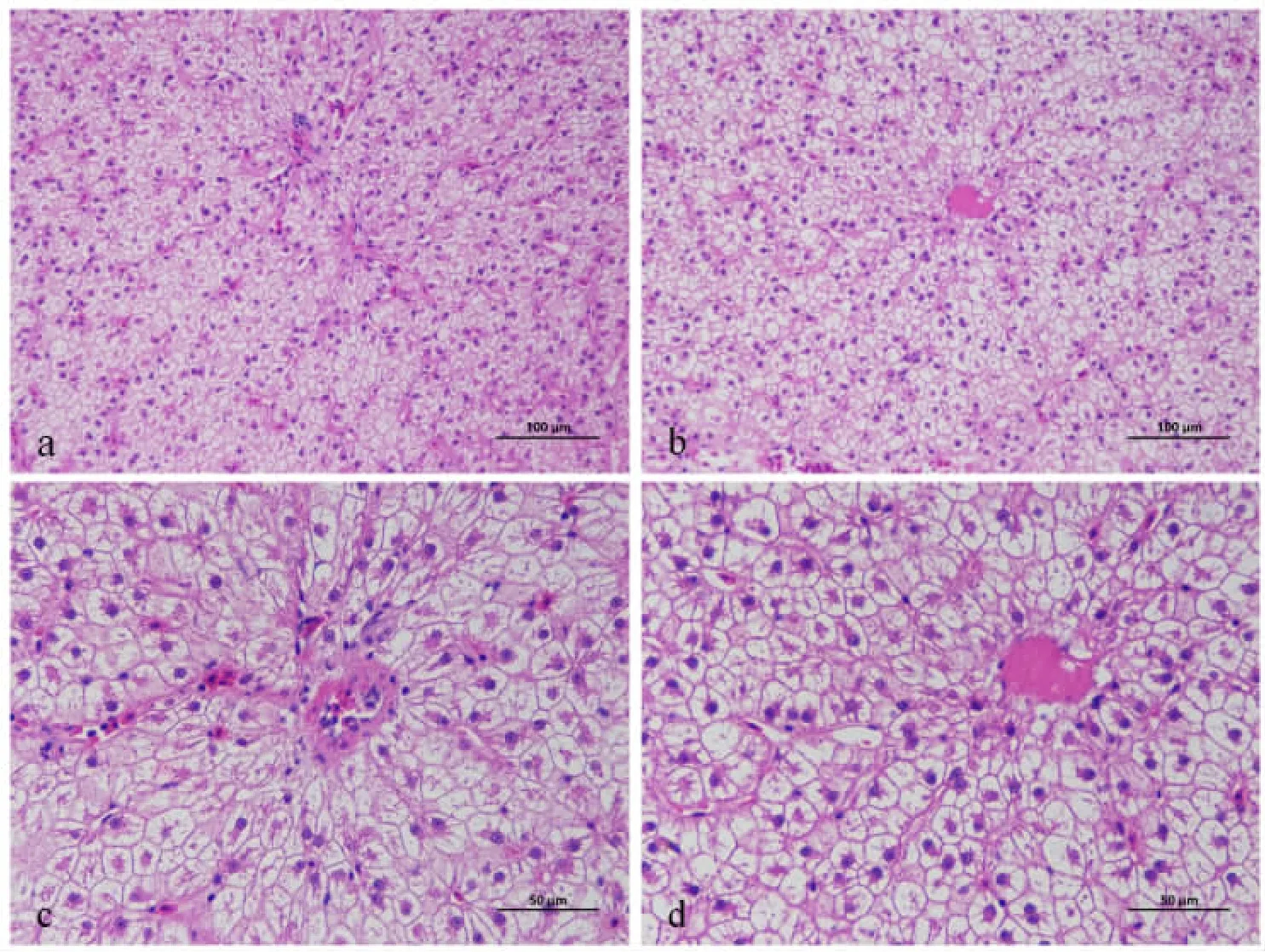
图2 发酵豆粕替代鱼粉对湘云鲫肝脏形态结构的影响
2.4.2 发酵豆粕对湘云鲫血浆生化指标的影响由表8 可知, 与FM 组相比,FSM 组血浆谷丙转氨酶活性、总胆固醇含量显著降低(P<0.05),其中谷丙转氨酶活性降低了11.3%,总胆固醇含量降低了23.1%,溶菌酶活性、免疫球蛋白M 含量显著提高(P<0.05)。FM 组与FSM 组间谷草转氨酶活性、甘油三酯、高密度脂蛋白胆固醇、低密度脂蛋白胆固醇含量、HDL-C/LDL-C 无显著差异(P>0.05)。

表8 发酵豆粕对湘云鲫血浆生化指标的影响
2.4.3 发酵豆粕对湘云鲫肝脏抗氧化能力的影响由表9 可知,FM 组与FSM 组肝脏总抗氧化能力、总超氧化物歧化酶活性、 丙二醛含量均无显著差异(P>0.05)。

表9 发酵豆粕对湘云鲫肝脏抗氧化能力的影响
3 讨论
经微生物发酵后,由于消除了抗营养因子,并增加了粗蛋白质、小肽和氨基酸等含量,发酵豆粕表现出更好的鱼粉替代效应(Wang 等,2016)。 近年来, 发酵豆粕在水产动物饲料中引起了越来越多的关注, 尤其是在鱼粉替代方面(Rahimnejad等,2019;Liang 等,2017;冯建等,2016)。发酵豆粕替代鱼粉在鱼类饲料中研究广泛, 但不同物种间其添加量存在很大差异 (祝焱彬等,2018;Barnes等,2015),这可能与养殖品种、饲料组成以及微生物降解抗营养因子成功率等差异有关。 在大口黑鲈(何明等,2020)饲料中以豆粕、发酵豆粕替代鱼粉的比较研究中发现,发酵豆粕可以替代30%鱼粉而不影响其生长性能,而豆粕仅能替代15%的鱼粉,豆粕经发酵后,干物质、粗蛋白质、总氨基酸和磷的表观消化率显著提高。 此外,发酵豆粕替代暗纹东方鲀饲料中的豆粕可提高其生长性能以及饲料利用能力(孔纯等,2017)。 本研究发现,添加12%发酵豆粕部分替代饲料中的鱼粉和豆粕对湘云鲫的生长性能及饲料利用无影响。 研究表明,发酵豆粕替代饲料中适量的鱼粉不会影响水产动物的生长性能(Choi 等,2020;Rahimnejad 等,2019)。Rahimnejad 等(2021) 在以花鲈为模型的试验发现, 低剂量发酵豆粕替代鱼粉不影响花鲈生长性能,替代量过高时其增重率、摄食量、蛋白质效率随替代水平的增加而下降, 最适鱼粉替代量为26.9%~37.1%。Choi 等(Choi 等,2020)在虹鳟上的研究发现, 当发酵豆粕替代鱼粉水平低于40%时,不影响虹鳟的增重率以及饲料转化率,当替代率达到60%时,则会降低其增重率以及饲料效率。
肠道是营养物质消化、吸收的主要场所,其消化和吸收相关酶的活性直接影响机体对外界营养物质的消化和吸收速率。本试验中,发酵豆粕提高了湘云鲫肠道脂肪酶活性, 而对其他消化吸收酶活性无影响。 湘云鲫肠道消化能力的提高与生长性能部分的结果趋势一致。除此以外,肠道是机体抵御病原微生物入侵的第一道屏障, 该屏障允许吸收养分,阻止病原体入侵,其结构和功能完整性是维持养殖动物肠道健康的基础 (Liu 等,2021;Lauriano 等,2016)。 血清D-乳酸和DAO 酶活性是用作评估肠黏膜屏障功能的间接指标(Zhang等,2018),当屏障功能受损时,细胞内的D-乳酸会释放出来,进入血液循环系统,从而对机体产生毒害作用。在日本鲈鱼(Rahimnejad 等,2019)和花鲈(Rahimnejad 等,2021)上的研究表明,血清D-乳酸含量随发酵豆粕替代量增加而增加, 低发酵豆粕组DAO 酶活性与对照组无显著差异。在仔猪(冯江鑫等,2020)上的研究表明,发酵饲料组肠道ZO-1、CLDN-1 基因相对表达量上调, 增强了肠道屏障功能,本试验结果与之相似。 有研究表明,在小鼠(Li 等,2020)酒精性肝病模型中使用乳杆菌混合物后, 肠上皮通透性改善以及肠道菌群中革兰氏阴性细菌减少导致进入门静脉的LPS 的减少,提高了肠道屏障功能。 故在本试验中,猜测可能是发酵饲料中的有机酸发挥了作用, 提高了湘云鲫肠道屏障功能。
血液学指标通常被用作评估鱼类生理和健康状况的关键诊断指标,并受季节、温度和营养变化的影响(Van Doan 等,2017;Dawood 等,2016)。 本试验中, 发酵豆粕替代部分鱼粉和豆粕降低了血浆中总胆固醇含量, 这与花鲈 (Rahimnejad 等,2021)、虹鳟(Matsunari 等,2010)、大黄鱼(冯建等,2016)中得到的结果相一致。研究表明,大豆蛋白替代动物性蛋白源可以降低养殖动物血液中的胆固醇水平, 这主要与大豆中所含异黄酮有关(Tsai 和Huang,1999),此外,大豆蛋白水解产物也通过增强肝细胞中的LDL-R 转录来降低血液中的胆固醇水平(Cho 等,2007)。血浆谷丙转氨酶和谷草转氨酶是肝细胞损伤的敏感指标(Boone等,2005)。 在日本鳗鲡中(李宁宇等,2020),以发酵豆粕替代鱼粉, 其血清谷丙转氨酶活性显著降低。 在卵形鲳鲹(李秀玲等,2019)、花鲈(Rahimnejad 等,2021)、梭鱼(祝焱彬等,2018)中发现,血清谷丙转氨酶活性随发酵豆粕添加量的增加而上升,但低剂量组与对照组无显著差异。 本研究中,添加发酵豆粕降低了湘云鲫血浆谷丙转氨酶活性, 与上述研究结果相一致, 表明饲料中添加12%发酵豆粕不会对湘云鲫肝脏造成损伤, 肝脏H&E 染色切片的结果也说明了这一点。
溶菌酶和免疫球蛋白在非特异性免疫和特异性免疫方面发挥了重要作用 (Yu 等,2018;Zhao等,2014)。 溶菌酶可以降解革兰氏阴性菌的细胞壁,是先天免疫的重要组成部分(Ghanei-Motlagh等,2021)。 硬骨鱼体液中的免疫球蛋白M 是特异性免疫系统的主要组成部分,可以作为鱼类免疫应答的重要标志物 (Chen 等,2013;Dominguez 等,2004)。有研究表明,豆粕经发酵产生的低分子量肽和氨基酸可以增强机体的免疫功能 (Li 等,2021;Zhuo 等,2014)。 本试验中,添加发酵豆粕提高了湘云鲫血浆溶菌酶活性以及IgM 含量,表明发酵豆粕可以提高湘云鲫的免疫能力。 这与在日本鲈鱼(Rahimnejad 等,2019)、 凡纳滨对虾 (刘韬等,2022)、异育银鲫(陈萱等,2005)、黑鲷(彭翔等,2012)中的结果一致。另外,发酵豆粕中包含的微生物可以作为益生菌来增强鱼类的免疫反应(Zhou等,2010)。
4 小结
综上,在等氮等脂等赖氨酸等蛋氨酸前提下,以12%发酵豆粕替代饲料中部分鱼粉和豆粕,可在不影响湘云鲫幼鱼生长性能及饲料利用的前提下,提高湘云鲫肌肉蛋白质含量,促进脂质代谢,增强免疫机能。
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2022年6期)2022-06-29
当代水产(2019年11期)2019-12-23
当代水产(2019年7期)2019-09-03
当代水产(2019年6期)2019-07-25
当代水产(2019年6期)2019-07-25
当代水产(2019年4期)2019-05-16
当代水产(2019年1期)2019-05-16
东坡赤壁诗词(2018年3期)2018-07-16
生命科学研究(2018年1期)2018-05-29
