
吉林蛟河针阔混交林生物与非生物因素对生产力的影响*
2024-01-20 10:02范秀华岳庆敏韩卓秀黄一鑫
林业科学 2023年12期
张 萌 范秀华 岳庆敏 韩卓秀 黄一鑫
(1.北京林业大学理学院 北京 100083;2.北京林业大学国家林业和草原局森林经营工程技术研究中心 北京 100083)
作为陆地生态系统的主体,森林植被生物量和初级生产力约占全球陆地生物量的85%和总初级生产力的60%,在调节全球碳循环中发挥着重要作用(Houghtonet al.,2009)。森林生物量净变化量是由保留木、进阶木带来的生物量增长以及死亡木造成的生物量损失3 个生物量动态过程构成的(Brienenet al.,2015),探究生物与非生物因素对不同生物量动态过程的影响,对理解森林生态系统功能、制定森林管理计划具有重要意义(Yuanet al., 2019;Yueet al.,2020)。当前,已有学者提出用植被数量、林木分化程度、生物多样性、地形等因素来解释生产力变异(Zhanget al.,2015;Haoet al.,2018)。
植被数量假说提出驱动生态系统功能的是植被数量(即断面积或生物量)而不是质量(即生物多样性或组成)(Lohbecket al.,2015)。森林演替早期,单位面积胸高断面积增加会使叶面积指数上升(Coomeset al.,2012),植被数量与生产力的正相关关系更紧密;随着演替进行,单位面积胸高断面积增加可能导致光可利用性降低,从而引起生产力下降(Prado-Junioret al.,2016)。另外,由于林下光照限制、大树死亡概率增加,单位面积胸高断面积大的群落常具有高死亡量(Prado-Junioret al.,2016)。
林木分化程度高可促进光捕获,提高光资源利用效率(Zhanget al.,2015);同时,林木分化程度也反映群落生态位分化(Yuanet al.,2018),较高的林木分化程度使个体间竞争减少,增强资源利用效率。多项研究表明,林木分化程度对生产力有促进作用(谭凌照等,2017;吴兆飞等,2019);但也有研究显示,林木分化程度与生产力呈负相关,其相关关系受造林密度等因素影响(杨桂娟等,2019)。
提高物种多样性,可使群落生态位差异增大,提升资源利用效率,从而提高生产力(Zhanget al.,2012);然而,共存物种数量增加也可能导致生态位重叠,使物种多样性与生产力的关系不显著甚至呈负相关关系(Prado-Junioret al.,2016)。系统发育多样性反映共存物种生态特征的差异,相比物种多样性更能准确表示生物多样性与生产力的关系(Haoet al.,2018)。
地形因子通过影响资源可利用性进而影响森林生产力(Ouyanget al.,2019),其效应可能与林木存活与生长的限制因子有关,但对生产力的具体作用仍存在争议(Haoet al.,2018;Yueet al.,2020)。
本研究以吉林蛟河针阔混交林固定样地为研究对象,利用多元回归分析,探究生物(单位面积胸高断面积、林木分化程度、生物多样性等)与非生物(地形因子)因素对保留木生产力、进阶木生产力和死亡量的影响;采用变差分解法分析3 种生物量变化对生物量净变化量的相对重要性,旨在通过量化不同因子对生产力的影响,阐明生物与非生物因素对林分生产力的影响和维持作用,为东北针阔混交林可持续经营提供科学依据和理论指导。
1 研究区概况
研究区位于吉林省蛟河市林业实验区管理局林场(43°57′75″—43°57′91″N,127°43′62″—127°44′05″E),地处长白山系张广才岭山脉,海拔468.5~486.49 m。属温带大陆季风性气候,年均气温3.8 ℃,最热月7 月平均气温21.7 ℃,最冷月1 月平均气温-18.6 ℃。年均降水量700~800 mm,主要集中在夏季。土壤类型为暗棕色森林土,土层厚度20~90 cm。植被类型属针阔混交林。主要乔木树种包括红松(Pinus koraiensis)、紫椴(Tilia amurensis)、水曲柳(Fraxinus mandshurica)、胡桃楸(Juglans mandshurica)、 蒙古栎(Quercus mongolica)、白牛槭(Acer mandshuricum)、色木槭(Acer pictumsubsp.mono)和春榆(Ulmus davidianavar.japonica)等。 主要灌木包括暴马丁香(Syringa reticulatasubsp.amurensis)、毛榛(Corylus mandshurica)和簇毛槭(Acer barbinerve)等。
2 研究方法
2.1 样地调查与数据获取
2010 年布设20.48 hm2(320 m×640 m)固定样地,划分为512 个20 m×20 m 连续样方。同年初选择样地内所有胸径(DBH)≥1 cm 的木本植物,记录物种名、胸径、树高、冠幅、枝下高及坐标,并挂牌标记,2015、2020 年分别进行第1 次和第2 次复测。以样方4 个顶点海拔的平均值作为样方海拔。样方任意3 个顶点组成4 个平面,将4 个平面相对于水平面产生的夹角平均值作为样方坡度,相对于垂直平面产生的夹角平均值作为坡向。相关统计结果见表1。
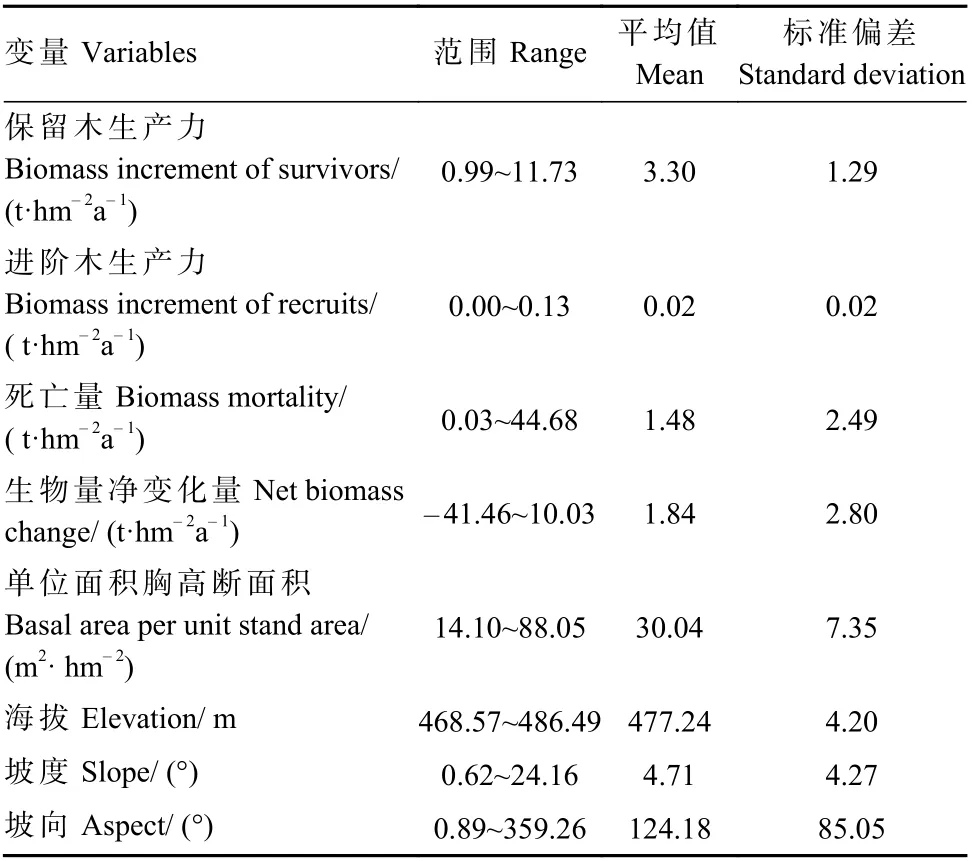
表1 样地内基础变量信息统计Tab.1 The statistical information of basic variables in the sampling plot
2.2 生产力计算
基于2010 年初和2020 年测定数据,利用异速生长方程(Haoet al.,2020)计算胸径(DBH)≥1 cm 个体的地上生物量。保留木代表初测与复测均存活的个体,保留木生产力为各样方中保留木年均生物量增量之和。进阶木表示初测未达起测标准(1 cm)而复测达到起测标准的个体,进阶木生产力为各样方中进阶木年均生物量增量之和。死亡木代表初测时存活而复测时死亡的个体,死亡量为各样方中死亡木年均生物量损失量之和。用进阶木生产力与保留木生产力之和减去死亡量得到样方生物量净变化量。
2.3 生物多样性及林木分化程度指标计算
以稀释物种丰富度、物种辛普森指数、系统发育多样性指数代表各样方生物多样性。因稀释物种丰富度修正了样方中个体数量对物种丰富度的正效应(Prado-Junioret al., 2016),故可代替物种丰富度。在样方中随机抽取一定数量个体,其物种数量即为稀释物种丰富度。本研究中,由于样方树木个体数量≥20,因此将随机抽取的个体数设置为20。物种辛普森指数代表在样方中随机取样的2 个个体不属于同一物种的概率,同时考虑了丰富度和均匀度。系统发育多样性指数即样方内个体的系统发育树总枝长(Faith,1992)。通过获取样地内个体科、属、种的标准拉丁名,利用Phylomatic(http://phylodiversity.net/phylomatic/)生成系统发育树,使用picante 包pd 函数计算获得系统发育多样性指数。
利用胸径辛普森指数、胸径变异系数代表各样方的林木分化程度。胸径以1 cm 为单位划分等级,统计每个样方内胸径等级数及每个胸径等级内个体数,根据辛普森指数公式,用胸径等级数替换物种数,即可计算胸径辛普森指数。胸径变异系数代表样方内树木胸径大小不等的程度(Yuanet al.,2018)。相关计算公式见表2,计算结果见表3。

表2 物种多样性和林木分化程度计算公式①Tab.2 The formulas of species diversity and degree of tree differentiation in the studied forest
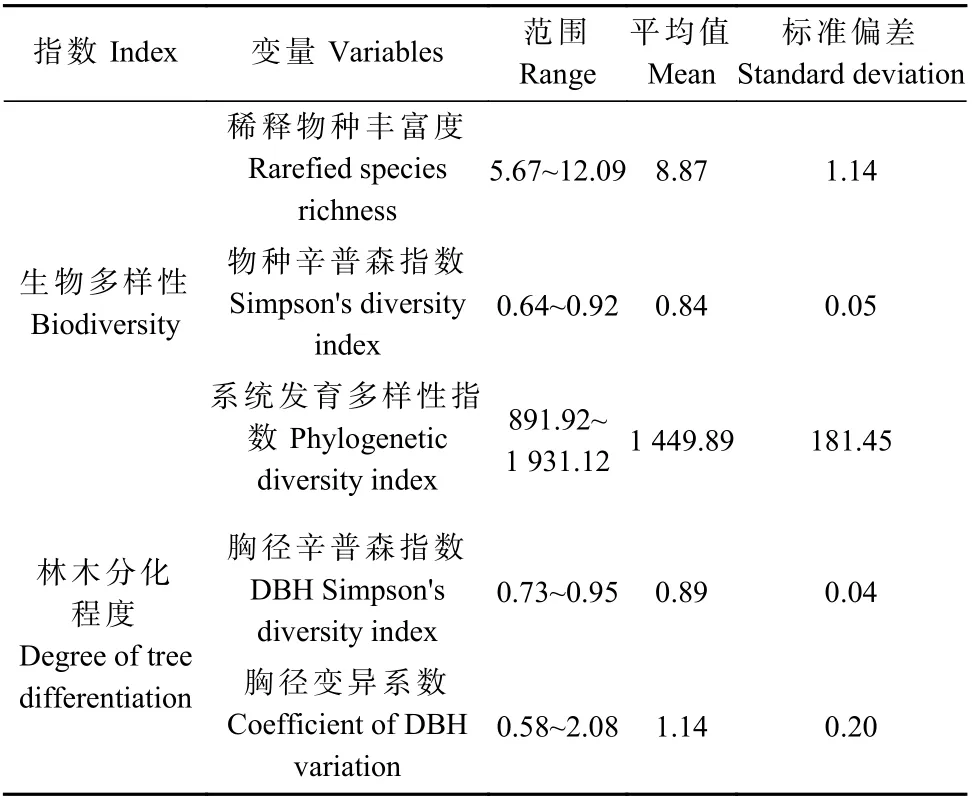
表3 生物多样性和林木分化程度统计结果Tab.3 The statistical results of biodiversity and degree of tree differentiation
2.4 统计分析
对生产力、死亡量进行自然对数转换以提高正态性,采用Z-score 方法对解释变量进行标准化处理以消除解释变量单位不同对研究结果的影响。多重共线性会影响解释变量对响应变量的解释和预测能力,利用方差膨胀因子(VIF)检验变量间多重共线性。当VIF<10 时,认为变量间没有多重共线性(Foxet al.,2008)。结果显示,胸径辛普森指数VIF>10,故删去该变量。使用多元线性回归评估所有解释变量对生产力和死亡量的效应,全模型包括单位面积胸高断面积、地形因子(海拔、坡度、坡向)、生物多样性指标(稀释物种丰富度、物种辛普森指数、系统发育多样性指数)、林木分化程度指标(胸径变异系数)。基于AICc值进行模型筛选,选出与最小AICc 差值<2 的所有模型,并对其进行加权平均,模型间无显著差异。为探究保留木生产力、进阶木生产力、死亡量对生物量净变化量的相对影响,采用变差分解法计算3 种生物量动态过程对生物量净变化量方差的解释比例。所有计算均在R4.0.3 中运行。方差膨胀因子(VIF)检验用car 包vif 函数,模型筛选与加权平均用MuMIn 包dredge、model.avg 函数,变差分解用vegan 包的varpart函数。
3 结果与分析
3.1 各因素对生产力、死亡量的影响
各解释变量的多元回归模型结果见表4。
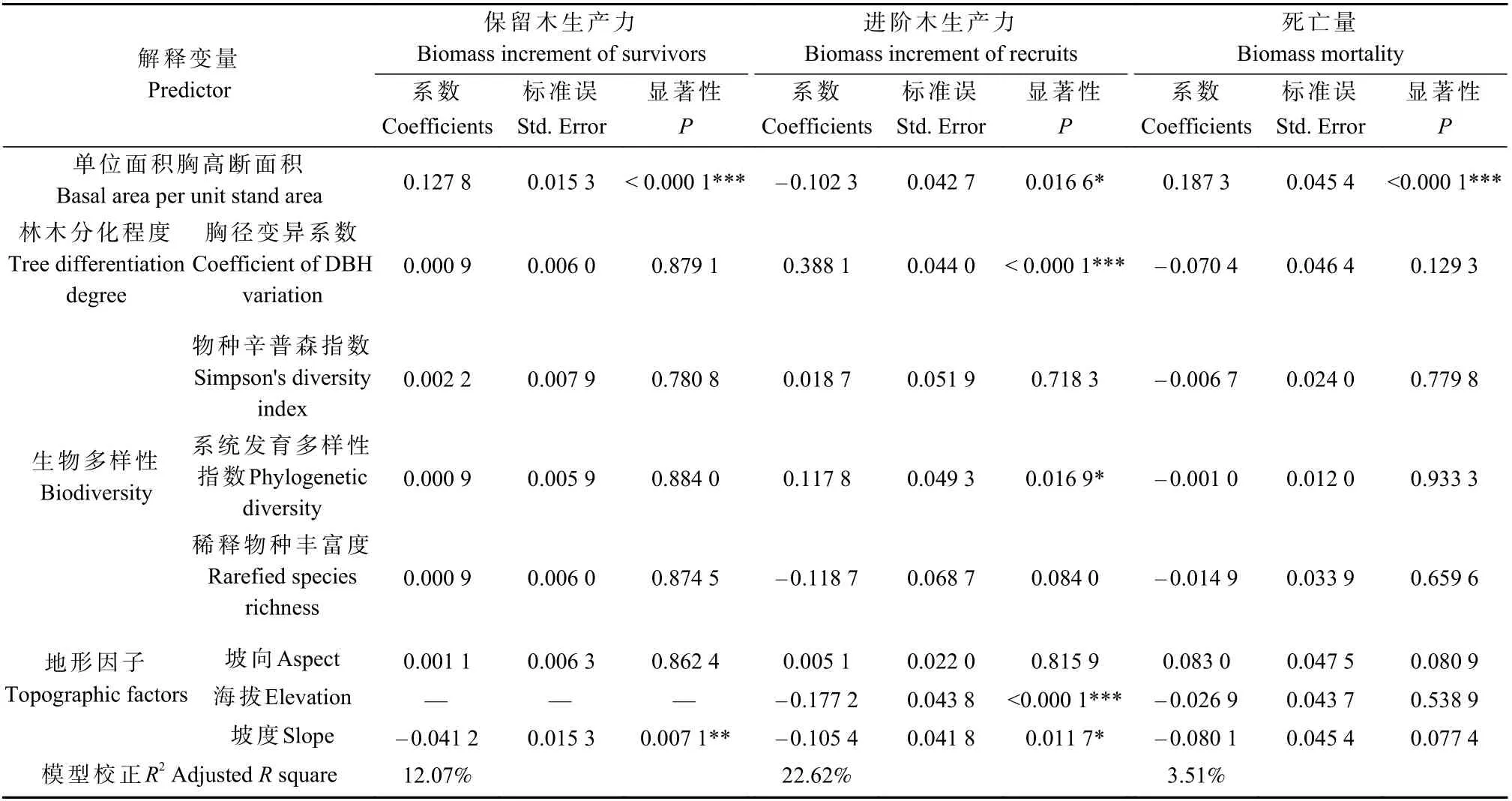
表4 多元回归模型结果Tab.4 Results of multiple regression model
对保留木生产力,所有解释变量可解释其总方差的12.07%,单位面积胸高断面积有显著正效应,坡度有显著负效应,其他解释变量无显著影响。
对进阶木生产力,所有解释变量可解释其总方差的22.62%,胸径变异系数和系统发育多样性指数有显著正效应,单位面积胸高断面积、海拔和坡度有显著负效应,其他解释变量无显著影响 。
对死亡量,所有解释变量可解释其总方差的3.51%,单位面积胸高断面积有显著正效应,其他解释变量无显著影响。
3.2 生物量动态过程对生物量净变化量的相对影响
死亡量单独解释生物量净变化量总方差变化的74.54%,保留木生产力单独解释20.87%,进阶木生产力单独解释0.01%;保留木生产力与进阶木生产力协同效应共同解释0.40%,保留木生产力与死亡量协同效应共同解释2.40%,进阶木生产力与死亡量协同效应共同解释4.44%。可见,生物量净变化量的主要决定因素为死亡量和保留木生产力,进阶木生产力贡献较小。
4 讨论
4.1 单位面积胸高断面积对不同生物量变化量的影响
单位面积胸高断面积对保留木生产力有显著正效应,是解释变量中最重要的预测因子。单位面积胸高断面积较高的群落通常拥有更多的大树,这些大树对生物量增长的贡献更大(Stephensonet al., 2014),且这些群落通常具有较大叶面积指数,提高了光截获效率,叶片可通过光合作用固定更多碳(Coomeset al.,2012;Michaletzet al.,2014)。然而,Yuan 等(2018)发现,植被数量对生产力的作用强度随群落生物量提高逐渐降低,这可能是竞争加剧或负密度制约效应造成的(Fineganet al., 2015)。单位面积胸高断面积对进阶木生产力有显著负效应,这是因为林下光能利用率低和水分竞争加剧导致进阶木生长缓慢(van der Sandeet al., 2017)。较高的单位面积胸高断面积提高了大树死亡概率(Prado-Junioret al.,2016),由于光照限制、树木枯倒或树枝坠落,林下植被死亡量增加(McDowellet al.,2018)。单位面积胸高断面积对3 种生物量动态过程均产生显著影响,显示植被数量假说在调节森林生态系统功能中的重要作用。
4.2 林木分化程度对不同生物量变化量的影响
群落中个体大小分化能促进光捕获,提高光利用效率。另外,林木大小差异造成光环境异质性,这有利于对资源有不同需求的个体共存,在林木大小均一的群落中,个体对资源需求相似,减弱生态位分化,加剧竞争,不利于个体生长(吴兆飞等, 2019)。本研究发现,林木分化程度仅对进阶木生产力有显著正效应,而其对保留木生产力无显著效应,以往研究也得出类似结论(de Avilaet al.,2018)。这可能是由于林木分化程度增加时,上层木的竞争力下降,下层木的竞争力上升(Aliet al.,2017)。另外,进阶木大多生长于光照强度较弱的林下,光照条件改善有助于提高其生长速度,而保留木大多生长于上林层,可能无法从多样的垂直结构中获益(Ali,2019)。
4.3 生物多样性对不同生物量变化量的影响
生物多样性通过提高资源利用效率、降低宿主特异性病原体的侵害等途径促进林木生长并避免林木死亡(Prado-Junioret al.,2016;van der Sandeet al.,2017)。大量研究证实生物多样性能提高生态系统生产力、降低死亡量(郭志华等,2002; Yuanet al.,2019)。本研究只观察到生物多样性提高群落进阶木生产力,并未对保留木生产力和死亡量产生显著影响。在环境条件恶劣时,互补效应对树木生长的促进作用较强(Mori,2018),进阶木位于光照条件较弱的林下,经历强烈光竞争,因此,多样性提高产生的生态位互补对进阶木生长的正效应较强。多样性对保留木生产力无显著效应,可能是由于保留木生产力受优势木性状的影响更大(Grime,1998)。本研究未观察到生物多样性对死亡量有显著影响,可能由于群落死亡量主要是大树死亡贡献,其死亡易受强风、闪电等随机事件影响,生物多样性效应相对较弱。系统发育多样性对树木生长的影响大于物种多样性,与物种多样性相比,系统发育多样性能量化共存物种生态特征的差异,从而更准确反映生物多样性与生产力的关系(Purschkeet al.,2013;Haoet al., 2018)。
4.4 地形因子对不同生物量变化量的影响
在林分水平上,海拔、坡度、坡向等地形因子的差异造成微环境异质性(水、光、土壤养分有效性等资源的差异),进而影响生产力(Fortunelet al., 2018)。本研究发现,保留木生产力随坡度增加而降低,进阶木生产力随海拔、坡度增加而降低。这可能是由于坡度较大、海拔较高的植被群落,其土壤水分和养分条件较差,导致生产力下降(Haoet al., 2018;温纯等,2019)。另外,地形因子影响树种的多样性和分布,进而影响林分的物理属性(如树木大小分布)和生物属性(如叶片生理特征),从而对生产力产生影响(Fotiset al.,2018)。与保留木、死亡木相比,进阶木受地形因子的影响更强,体现出幼树对环境条件更敏感。本研究表明生境异质性对森林生态系统具有调节作用。
4.5 不同生物量变化量对生物量净变化量的影响
生物量净变化量主要受死亡量和保留木生产力影响,这可能是由于与进阶木生产力相比,死亡量和保留木生产力数值更大。死亡量的模型校正R2最低,可预测性最低,与van der Sande 等(2017)研究结果相似。树木死亡在推动森林功能组成的变化中起到关键作用(Ruiz-Benitoet al.,2017a;2017b),从而对生态系统产生影响。在演替早期和干扰后的森林恢复阶段中,进阶木生产力对生物量净变化量的影响可能更强(Prado-Junioret al.,2016)。在个体尺度、区域尺度上,死亡量可较准确地预测(Chaoet al.,2008;Quesadaet al.,2012),然而在局域尺度上,林分死亡量受大树死亡、单株树木衰老及强风等自然条件影响,具有较大随机性(van der Sandeet al.,2017)。
5 结论
高的单位面积胸高断面积可促进林木死亡和保留木生长,抑制进阶木生长。林木分化程度及系统发育多样性可促进进阶木生长。坡度对保留木及进阶木生长均有抑制作用,海拔仅对进阶木生长有抑制作用。生物量净变化量主要受死亡量和保留木生产力影响。在森林经营中,应维持合理的单位面积胸高断面积,以促进保留木和进阶木生长,减少林木死亡,并通过提高林分中个体大小不均匀性和系统发育多样性来促进林木更新。
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
林业科学研究(2021年6期)2022-01-05
内蒙古林业调查设计(2021年5期)2022-01-05
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
林业调查规划(2020年3期)2020-06-03
汽车观察(2019年2期)2019-03-15
汽车观察(2018年12期)2018-12-26
汽车观察(2018年12期)2018-12-26
能源(2018年8期)2018-09-21
