
基于模型模拟的QTT 台址树木形成层物候与气候因子对径向生长的响应*
2023-12-19 08:40蒋靖张勇康剑
林业与环境科学 2023年5期
蒋 靖 张 勇 康 剑
(1.中国科学院新疆天文台, 新疆 乌鲁木齐 830011; 2.中国科学院华南植物园, 广东 广州 510650)
树木生长受到自身生理因素及环境气候因子的影响。 一般来说, 温度是寒冷地区树木生长的主要限制因子, 而在半干旱半湿润地区, 水分的作用更为重要[1-2]。 为了深入了解在气候变化背景下树木生长及其物候的动态变化, 往往需要开展长时间的微尺度监测[3]。 但这种监测在高海拔的山区是无法实现的, 因此, 可以运用VS 模型来对树木的生长动态进行长时间尺度的模拟[4-5], 从而揭示区域树木生长的限制因素以及形成层物候的动态变化。
QTT 奇台射电望远镜, 全称为110 m 口径全可动射电望远镜, 是中国科学院和新疆维吾尔自治区重点推进的国家重大科技基础设施, 选址于新疆奇台县境内的东天山北麓。 该区域具有海拔高、 气候干旱、 水汽含量低、 风速低、 无线电干扰少等特点。 基于其独特的地理位置及电磁环境,QTT 是世界一流的射电台址, 建成后将成为世界最大全可动射电望远镜, 具有极高的国际影响力。QTT 建成后, 其所在区域将是重要的国内国际天文观测、 科研交流、 科普教育的重要场所, 营造良好的工作学习环境显得至关重要。 本研究基于树轮模拟长时间尺度该区域树木生长动态, 探究气候变化背景下树木生长的限制因素, 以期为山区水土保持、 QTT 台址区树木植被建成及管理提供科学支撑。
1 材料与方法
1.1 研究区域
QTT 选址于新疆东天山北麓昌吉州奇台县境内, 属于典型的温带大陆性气候。 研究区年平均降水总量1 692 mm, 6—8 月降水较为集中, 年平均温5.95 ℃, 7 月平均温度最高(21.4 ℃) (图1)。 研究区内以森林、 灌丛、 草本植被为主, 雪岭云杉是该区域的优势种, 伴生有天山桦Betula tianschanica、 叉 子 圆 柏Sabinavulgaris、 黄 刺 玫Rosaxanthina等。 土壤类型以黑钙土为主[6]。

图1 研究区月平均气温、 降水变化趋势Fig.1 Variation trend of monthly mean temperature and precipitation in the study area
1.2 采样及年表
2022 年8 月于研究区东侧海拔1 100 m 处选取了1 个样地, 样地以雪岭云杉为优势树种, 伴生有天山桦, 样地中设置1 个30 m×30 m 的无人为干扰、 未经过火灾、 虫害等的林分作为样方, 样方树木平均胸径30.98 cm, 平均高度14.17 m, 样方内共选取目标树木20 株。 用生长锥在每株树木胸径处(树高1.3 m 处) 从平行于所在坡面的两个不同方向分别钻取一个样芯, 封装于塑料管中带回实验室。
采集的树轮样本在实验室经过固定、 干燥、打磨等处理后, 利用树轮宽度测量仪及配套TSAP 软 件 (LinTab 6.0, Frank Rinntech, Heidelberg, Germany) 测量年轮宽度 (精度为0.001 mm)。 采用目视初步定年、 骨架图定年等方式交叉定年, 确定每一轮的精确年份, 并使用COFFECHA 程序检测交叉定年的准确度以及缺失轮判断的可靠性[7]。 运用R 软件中的 “dplR”包[8], 使用响应截止频率为50%, 级数长度67%的样条函数对年表进行去趋势处理, 并建立标准年表。 同时, 计算平均敏感度(MS, Mean sensitivity)、 样本相关系数(rbar, Correlation coefficient for all series)、 信 噪 比 (SNR, Signal-tonoise ratio) 以及样本总体解释量 (EPS, Expressed population signal) 等年表统计参数[9], 评估年表的可靠程度。
1.3 气象数据
研究采用的气象数据来自中国气象数据网( http:/ /data.cma.cn/site/index.html ) ( 1991—2022 年) 以及荷兰皇家气象研究所(Koninklijk Nederlands Meteorologisch Instituut, KNMI) (CRU TS4.03 数据库, 数据精度为0.5°×0.5°, 1950—2022 年)。 包括平均气温(Tmean)、 降水(Prec)等指标。
1.4 数据分析
本研究基于VS 模型对树木季节性生长以及形成层物候进行模拟评估[10]。 模型通过分析气候和树木年轮生长过程之间的非线性关系, 将气候信号转换为依赖于温度、 土壤湿度和日照长度的部分树木年轮生长速率, 从而生成用于评估季节性形成层活动的累积生长速率[11-12]。 模型能够模 拟 出 受 温 度 (GrT)、 降 水 (GrW)、 光 照(GrI) 影响的部分生长速率, 并形成累计生长速率(Gr=GrI×min {GrT, GrW} )。 通过输入每日气象数据, 模型通过感知环境变化对树木生长的影响, 将树木年轮生长的速度转化为形成层细胞分裂生长的速度, 当春季累积生长速率大于0.1 时, 生长季开始, 相反, 当秋季累积生长速率小于0.1 时, 生长季结束[13-15]。 模型模拟的精度通过相关系数 (R)、 最小均方根误差(RMSE)、 Gleichlaufigkeit 统计量(Glk) 进行评估, 选 取1975—2022 作 为 校 准 区 间, 1950—1974 作为验证区间[16]。
运用R “treeclim” 包对年表与气候因子之间的Pearson 系数进行Bootstrap (1000 次迭代) 重采样分析(上一年9 月至当年9 月)[17]。 制图使用R 软件进行。
2 结果与分析
2.1 树轮年表基本特征
年表统计结果显示, 年表长度为1919—2022年。 平均敏感度0.26, 说明雪岭云杉树木生长对于区域气候敏感。 样本间相关系数较大, 信噪比值达到15.31, 表明该年表中用于分析的环境信息量较大[18]。 样本总体解释量远高于0.85 临界值,说明采集的样本量中包含的信号可以代表区域内的总体特征[9](表1)。 该年表对气候变化敏感,包含较多的气候信息, 可以开展树木年轮学的研究。
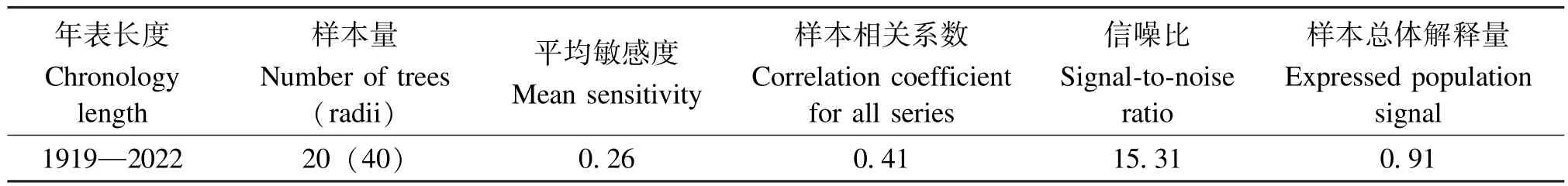
表1 树轮年表特征统计Table 1 Characteristics of tree-ring chronology
2.2 树轮模拟及生长速率的动态
模拟结果显示, 模拟年表与实测年表(1950—2022) 具有显著的相关性, 相关系数0.543 (p<0.05)。 年表间同步性(Glk) 72.603%, 远高于临界值50%。 RMSE 值为0.219, 低于临界值0.3, 表明模型模拟结果具有较高的精度。 通过分段校准验证, 结果显示, 校准段(1975—2022)R为0.499,Glk 为64.583%, RMSE 为0.184; 验证段(1950—1974)R为0.578, Glk 为84.000%, RMSE 为0.275 (图2)。 模型模拟效果良好, 表明生长率及形成层物候的模拟具有较高的可靠性。
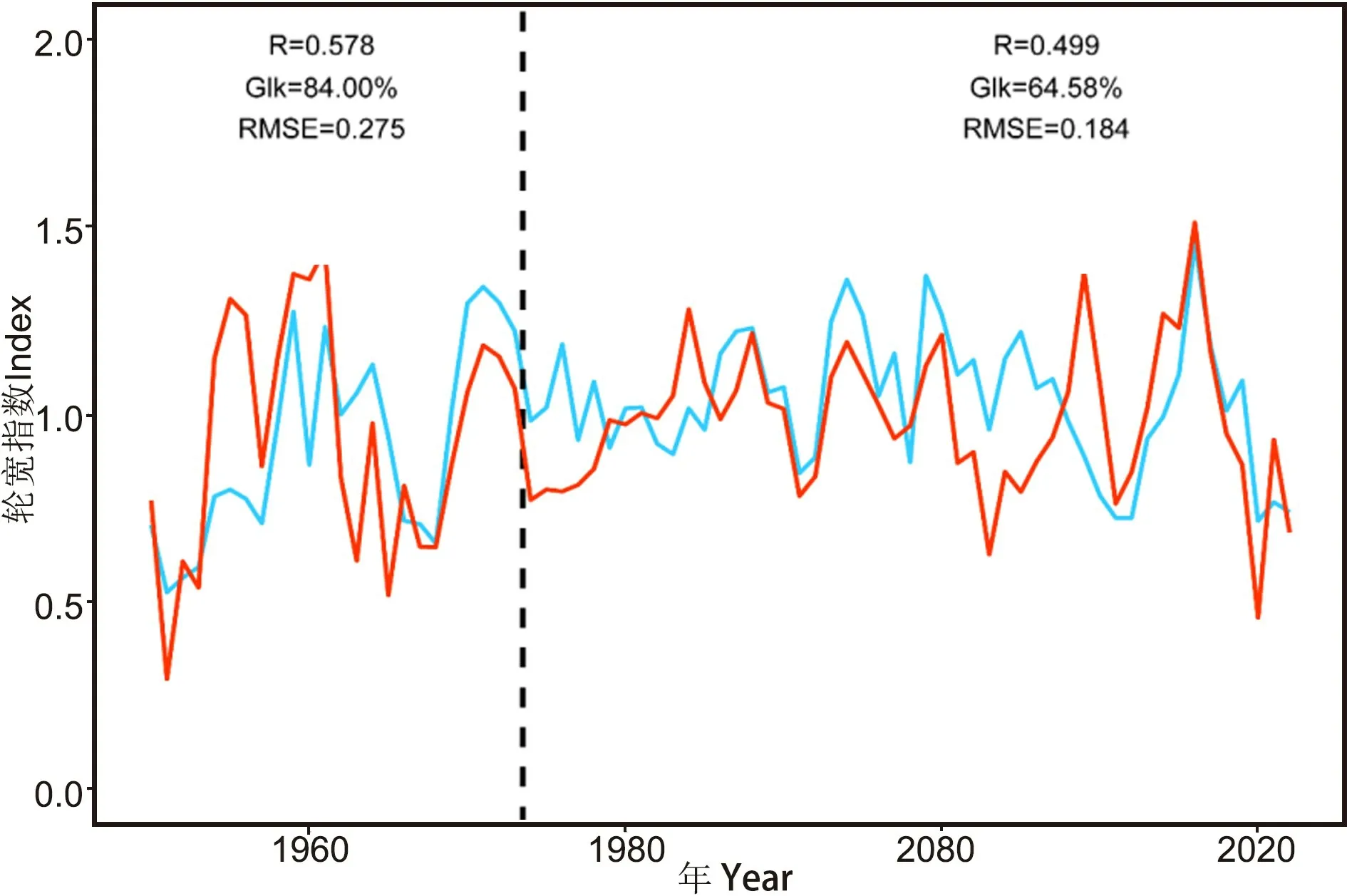
图2 模型模拟年表与实测年表对比Fig.2 Comparison of model simulation chronology and measured chronology
生长率的动态模拟结果显示, 整体生长速率Gr 呈现明显的右偏单峰分布, 在6 月初达到最大值0.53 (DOY 160)。 水分控制的生长率GrW显示出与Gr 基本一致的分布模式, 在5 月底达到峰值0.56 (DOY 143), 较Gr 提前了一段时间。 温度控制的生长率GrT在6—8 月达到峰值, 平均值为0.98 (DOY 170-231) (图3)。
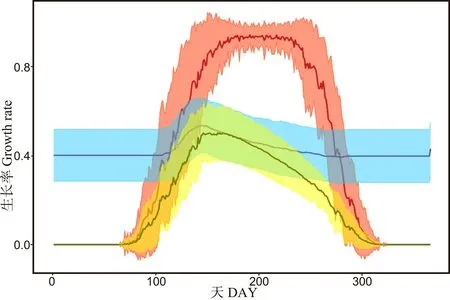
图3 年内平均的日尺度部分生长率及累计生长率Fig.3 Intra-annual variability in mean daily partial growth rates and mean integral growth rates
2.3 树木形成层物候的动态变化
在1950—2022 年间, 研究区树木生长的生长季起始时间(SOS) 不断提前, 平均每10 年提前1.5天(p<0.01)。 生长季结束时间(EOS) 尽管表现出一定的变化趋势, 但变化不显著。 生长季长度(DOS) 呈现明显的增长趋势, 平均每10 年增加2.3 天(p<0.01), 且增加趋势较为显著(图4)。
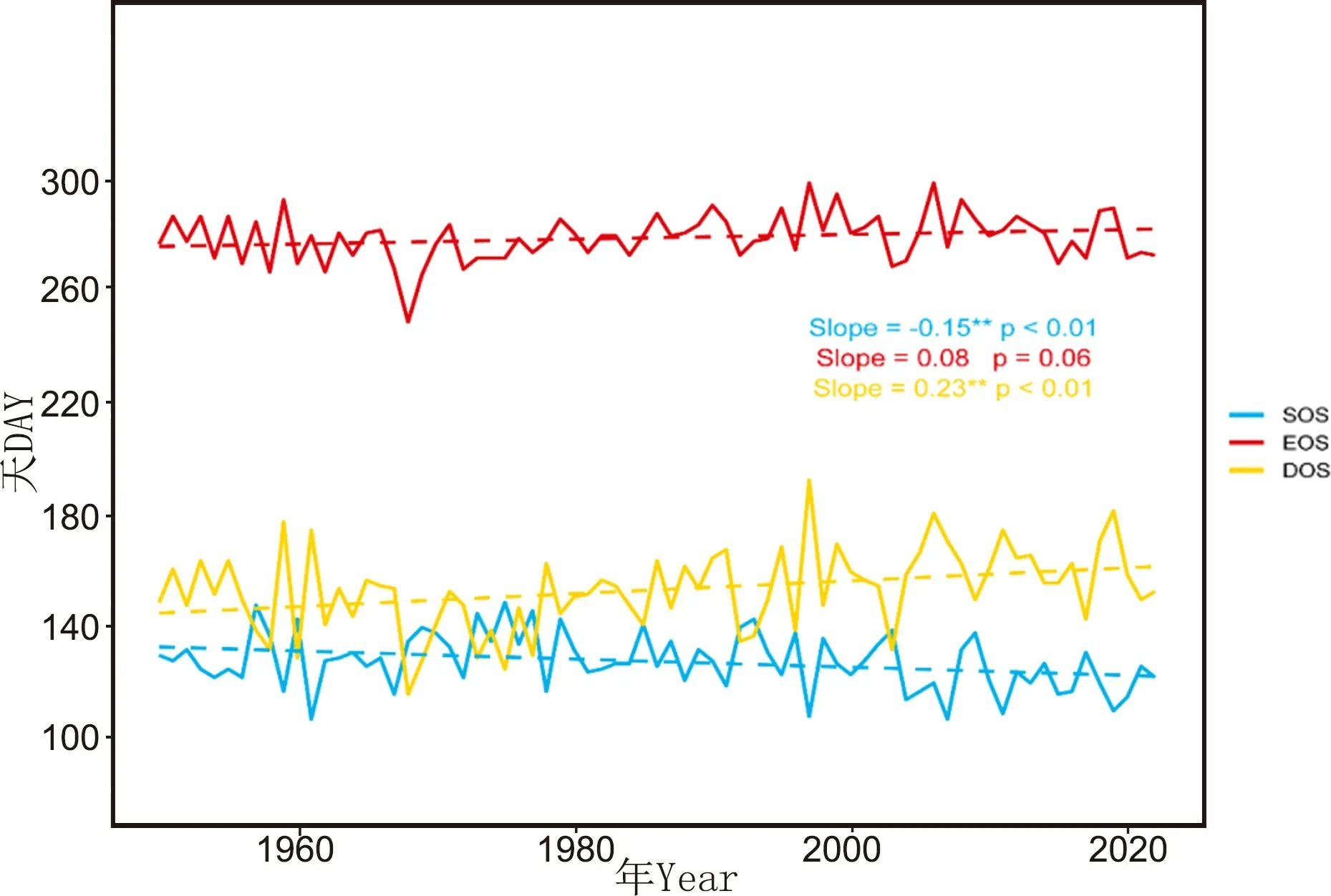
图4 年际间生长季开始、 结束以及长度的动态变化Fig.4 Dynamic of the start, end and length of the growing season
2.4 树木生长对气候的响应
通过对树轮年表与气候因子(1950—2022 年间) 进行相关性分析, 结果显示, 树木生长与当年5 月(r=-0.38,p<0.05)、 6 月(r=-0.34,p<0.05) 份平均气温呈现显著负相关关系。 研究区降水与树木生长间有着较为密切的联系, 树木生长与上一年9 月 (r= 0.21,p<0.05)、 11 月(r=0.20,p<0.05) 的平均降水显著正相关; 与当年4—6 月(4 月r=0.27, 5 月r=0.36, 6 月r=0.26,p<0.05)、 9 月(r=0.21,p<0.05) 降水呈现显著正相关关系(图5)。
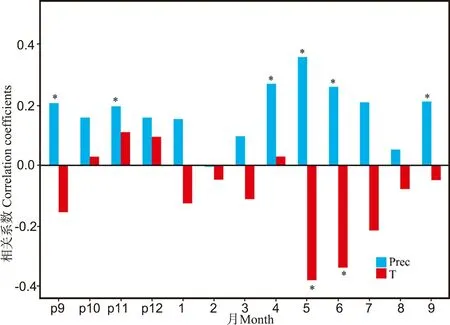
图5 树轮年表与气候因子的响应关系Fig.5 Correlation between ring width chronology and climate factors
3 讨论与结论
3.1 研究区树木生长呈现右偏单峰型, 在生长旺盛阶段, 温度控制生长率接近于1, 远高于水分控制的生长率, 说明水分是主要的生长限制因素。这一结果与阿尔泰山东南部的研究结果一致[19]。研究显示, 前期的水分限制对于生长季后期的生长速率产生了一定的负面影响, 导致了右偏单峰的结果[12]。 树木生长与降水的响应结果进一步证明了水分是研究区树木生长的重要限制因素。 一方面, 可能是由于研究区处于相对低纬度地区,温度相对较为适宜, 对于树木生长的限制作用较弱[20-21]。 另一方面, 研究区靠近准噶尔盆地, 干旱程度更高。 尽管有研究显示, 西北地区正在朝着暖湿化发展[22-23], 但这一趋势在局部区域地变化仍不明显。 在很长一段时间里, 土壤水分将是该区域树木生长的主要限制因子。
3.2 研究区树木生长季起始时间明显提前, 生长季长度显著延长, 平均每10 年延长2.3 天。 与中欧地区每10 年延长3.1 天相比略低[24], 与阿尔泰山东部结果较为一致。 随着全球气候变化, 温度升高, 早春更加温和的温度条件可以加快细胞的分化速度, 提前启动时间[14]。 研究表明, 春季温度是生长季节提前开始的主要驱动因素[25-26]。 与此同时, 温度升高也提前了融雪的时间, 积雪融化为形成层的早期发育提供了充足的土壤水分,能够促进形成层细胞的分裂和增大[27]。 在阈值范围内, 伴随着气候变暖, 树木生长的生长季起始时间会呈现持续提前的趋势, 生长季长度得以延长。 然而, 温度持续的升高可能会造成树木向更高海拔、 纬度地区迁移。
综上所述, VS 模型对于该区域的树木生长有着良好的模拟效果, QTT 台址区域树木生长呈现右偏单峰模式, 且树木生长主要受到水分的限制。伴随着气候变化, 水分限制将持续存在, 形成层物候起始时间提前, 生长季长度延长。 因此, 该区域开展树木植被营造、 管护过程中, 需要通过人工措施缓解生长季初期的干旱胁迫, 合理利用生长季延长的优势, 促进植被快速生长, 形成景观。 同时, 植被快速形成有助于台址区水土保持,实现防洪减灾的作用。
致谢:此次野外实验的数据获取得到了中国科学院新疆天文台奇台观测站的大力支持, 在此表示衷心感谢!
猜你喜欢
世界建筑(2022年10期)2022-10-25
广西农学报(2021年2期)2021-11-21
生态学报(2021年14期)2021-09-01
景德镇陶瓷(2021年1期)2021-03-24
林业科学(2020年10期)2020-11-30
西藏人文地理(2020年4期)2020-11-19
农业工程技术·综合版(2020年1期)2020-04-23
艺术品鉴(2019年12期)2020-01-18
环球人文地理·评论版(2016年6期)2016-08-23
山东农业科学(2014年4期)2014-07-18
