
不同年期发酵床垫料细菌群落多样性及功能
2023-12-06 03:04郑超超李买平柳晓东姜影影贺晓龙邓振山
微生物学杂志 2023年5期
郑超超, 李买平, 郝 雷, 柳晓东, 姜影影, 贺晓龙, 邓振山*
(1.延安大学 生命科学学院,陕西 延安 716000;2.延安市宝塔区畜牧兽医中心,陕西 延安 716000)
我国畜禽废物年总量高达20亿t,是工业废弃物的3.2倍。畜禽废物除了污染土壤、水体外,还会产生大量的臭味气体如氨气、硫化氢、硫醇和甲硫醇等,产生了养殖粪污堆积等问题,给环境及人畜的正常生活带来不利的影响。微生物发酵床技术是一种绿色养殖方式,通过分解消纳粪污符合环保要求,且利于家禽生长。发酵床养殖模式是一种环境友好的动物养殖技术,以木屑、稻壳、秸秆等农业废弃物为发酵床垫材料,添加微生物复合菌降解粪污,减少对环境的污染,改善畜禽的生存环境[1]。发酵床养猪模式的发展,不仅提升了养殖的经济效益,促进养殖产业生产能力及市场竞争力,同时解决了粪污堆积、环境污染及生猪健康养殖等问题[2]。微生物发酵床技术已列入国家重点环境保护应用技术中,对于解决畜禽粪便的污染问题具有很好的推广价值。目前,我国大部分发酵菌剂引进于国外,这与我国的养殖环境不相协调,菌剂微生物很难竞争过原有的土著微生物,发挥不了改良调节的作用等,有些菌剂如EM已经被应用于发酵床但是针对性不强,成本也相对较高。因此,发酵菌剂的研究势在必行。但引进发酵技术的同时,却忽略了对发酵床菌剂的改进研究。由于不同环境下,微生物的生长情况和代谢特性存在很大的差异,引进的发酵菌种往往在生产过程中出现各种问题,导致发酵床养殖成本升高、利润降低,甚至死床,已经成为该技术发展的瓶颈问题。目前国内专门应用于发酵床的菌剂很少,菌剂品质也良莠不齐。因此,发酵床垫料中存在的微生物是发酵床运行的核心,其微生物的种类及数量对发酵床功能的正常发挥及养殖动物的健康至关重要[3]。发酵床中细菌种类丰富,研究发酵床中蕴含的微生物群落,可累积丰富的微生物资源。由于发酵床的正常运行和功能发挥与其发酵床垫料中的相关微生物种类息息相关,因此,研究发酵床垫料微生物组成与结构具有重要意义。影响发酵床中微生物多样性因素众多,柳晓波等[4]研究发现与传统的水泥地养殖相比,发酵床养殖技术可提供稳定而丰富的菌群和稳定的温湿度,有利于猪仔生存。不同深度[5]和不同发酵程度发酵床垫料[6-7]的微生物组成水平也存在差异。常见的发酵床垫料的优势菌属有节杆菌属 (Arthrobacter)、芽胞杆菌属 (Bacillus)、梭菌属 (Clostridium) 及肠杆菌属(Enterobacte)[8]。目前,已有研究表明发酵床中的微生物随着发酵床垫料使用时间而发生变化[9]。发酵床技术的应用给养殖业带来了前所未有的发展,但由于不同地区环境存在差异,目前关于时间间隔较长且环境温差较大的陕北地区发酵床养殖及相关发酵床专用菌剂研制鲜有报道,且不同时段的发酵床垫料中细菌多样性变化对陕北地区生猪养殖具有启发意义。综上所述,开发陕北地区本土化复合发酵菌剂,建立微生物发酵床实现对畜禽废物的原位处理,减少排放,能在一定程度上降低生产成本,取得较好的环境和经济效益。鉴于此,本研究针对生猪养殖过程中造成严重污染和副产物资源化问题,围绕养殖污染微生物治理、微生物资源筛选,通过从陕北地区养殖场以及周边分离筛选出适合养殖环境、性能优良的菌株,使之在畜禽粪便中充分生长繁殖,形成优势菌群,形成发酵床复合菌剂。本研究选取延安市宝塔区通过发酵床技术养猪场铺设1、2、3和4年的发酵床垫料为研究对象,新铺设的发酵床垫料作为对照(CK),采用MiSeq高通量测序技术分析不同年期发酵床垫料细菌群落组成变化及其相互关系,以开发其中蕴含的有益微生物资源,为适合陕北地区的发酵床专用菌剂的研制、发酵床垫料的维护及养殖粪污降解等方面提供参考。
1 材料与方法
1.1 材料
1.1.1 研究区概况 样品采自延安市宝塔区姚店镇小张沟村(36°10′33″~37°2′5″N,109°14′10"~110°50′43"E)一家大型现代化养猪场,该养猪场于2016年从福建省农业科学院引进发酵床养猪技术,可为本研究提供充足的不同年限的发酵床垫料来源。采样点地处暖温带温和半湿润气候区,年平均温度9.7 ℃,一月平均气温-6.7 ℃,七月平均气温22.9 ℃,极端最高气温39.7 ℃,极端最低气温-28.4 ℃。最热月份平均相对湿度72%;年日照2 472 h;年降水量550 mm,57%集中在7~9月。
1.1.2 样品采集与处理 样品采集于2020年1月4日,新铺设的发酵床垫料作为对照(CK),试验组发酵年限分别为一年(1年期,FBM1)、两年(2年期,FBM2)、三年(3年期,FBM3)和四年(4年期,FBM4)的发酵床垫料。通过五点采样法采集样品,其中每一样点采集3个不同深度(0~10 cm、10~20 cm、20~30 cm)发酵床垫料样品混合,每个样点采集重量为10 g,共30 g。最后将五个样点采集的发酵床垫料混合为一个样品,共计150 g。一个年份采集3个样品,共采集15个发酵床垫料样品。分装于灭菌的自封袋中,低温保存运回实验室于-80 ℃保藏,用于土壤微生物DNA的提取及后续测试。
1.1.3 主要试剂与仪器设备 OMEGA-soil DNA Kit型DNA抽提试剂盒(Omega Bio-Tek,USA);biowest agArose 2%琼脂糖凝胶(biowest,Espana);FastPfu聚合酶(TransGen,China);Axygen Biosciences胶回收试剂盒(Axygen,USA)。DYY-6C电泳仪(北京市六一仪器厂,China);ABI GeneAmp®9700型PCR仪(ABI,USA);Illumina Miseq MISEQ测序仪(Illumina,USA);微型离心机(ABSON MiFly-6,合肥艾本森科学仪器有限公司);小型离心机(5430 R,Eppendorf,Germany);粉碎研磨仪(TL-48R,上海万柏生物科技有限公司)。
1.2 方法
1.2.1 基因组 DNA 的提取及扩增测序 2 mL离心管中加入0.5 g发酵床垫料,根据磁珠法土壤和粪便基因组DNA提取试剂盒(TIANGEN,中国)说明书进行总DNA提取。应用NanoDrop2000及1%琼脂糖凝胶电泳(电压5 V/cm,20 min)进行DNA纯度、浓度检测及DNA完整性检测。采用338F (5′-ACTCCTACGGGAGGCAGCAG-3′) 和806R (5′-GGACTACHVGGGTWTCTAAT-3′) 引物扩增细菌16S rRNA基因V3~V4区。每个样本3个PCR重复,将3个重复的PCR产物混合,使用2%琼脂糖凝胶电泳检测产物。利用Axygen Biosciences胶回收试剂盒对PCR产物纯化,QuantusTMFluorometer对其进行检测定量。根据Illumina、 San Diego、 DNA标准操作规程将纯化后的扩增片段构建PE2*300的文库。利用Illumina公司的Miseq PE300平台进行测序。
1.2.2 数据处理、发酵床垫料细菌群落多样性分析及功能预测 用Qiime2软件中插件 (DADA2) 对所有样品的全部原始序列 (input) 进行质量控制,修剪、去噪、拼接、去嵌合体,形成特征表格[10]。使用UPARSE软件(version 7.1 http://drive5.com/uparse/),根据97%的相似度对序列进行OTU聚类,应用UCHIME 软件剔除嵌合体,得到OTU代表序列;选出与OTU代表序列相似性在97%以上的序列,生成OTU表格。利用RDP classifier (http://rdp.cme.msu.edu/) 对每条序列进行物种分类注释,比对Silva 数据库(SSU128),设置比对阈值为70%。根据每个OTU中序列的条数获其OTU的丰度值[12-13]。选取物种绝对丰度前20进行优势物种分类。进行样品组间和组内共有及特有OTU数目分析、物种组成及物种差异性分析[14]。应用QIIME2 core-diversity插件计算多样性矩阵。Alpha多样性分析应用Observed OTUs、 Chao1、Shannon、Simpson和Faith′s phylogenetics diversity指数评估样本本身的多样性程度;Beta多样性指数应用Bray Curtis距离进行主坐标分析(PCoA)评估样本之间的微生物群落结构差异性。应用PICRUSt软件预测微生物群体可能的功能组成[15]。使用SPSS20.0软件进行单因素方差分析,并利用Duncan法进行差异显著性检验。
2 结果与分析
2.1 不同年期发酵床垫料细菌OTU及其多样性分析
经质量控制、去噪、拼接与去嵌合体(表1),共获得924 008条序列、686种OTU。1年期、2年期、3年期、4年期及对照组有效序列均值分别为41 030、44 136、38 654、43 179及37 394,其中最小值为CK-2(35 375),最大值为FBM2-1(48 150)。样品有效序列表明,不同年期发酵床垫料微生物组成有差异。特征序列数量最大为2年期,分别为1年期、3年期和4年期的1.07倍、1.14倍和1.02倍。四个年期发酵床垫料含量均高于对照组,表明发酵床垫料使用年期可影响其细菌多样性。样品OTUs范围在31 (CK) ~227 (FBM2) 之间,2年期OTU数目最多。稀释曲线 (faith-pd 图1) 显示,所有样品随着序列数量的增加,所观察到的OTU数量逐渐趋于平缓,达到饱和。说明随着测序深度增加,获得的新物种不会显著增加,即测序获得的数据量合理,能反映绝大多数的细菌信息。
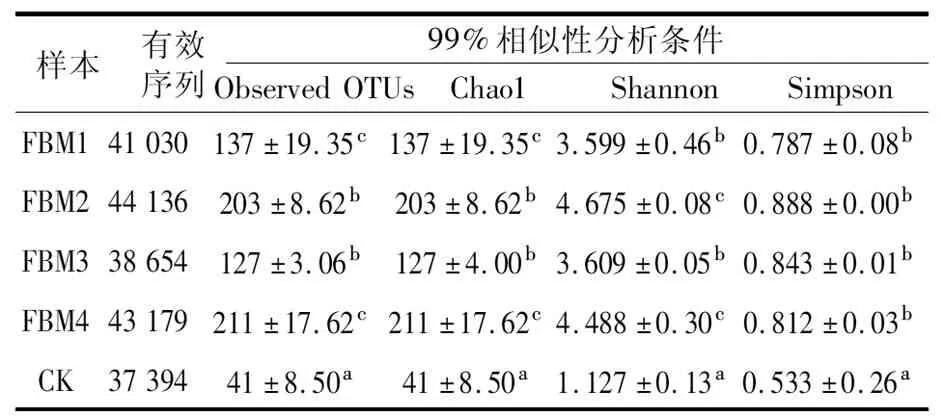
表1 发酵床垫料细菌多样性测序结果分析Table 1 The sequencing results of bacterial diversity of fermented mattress materials
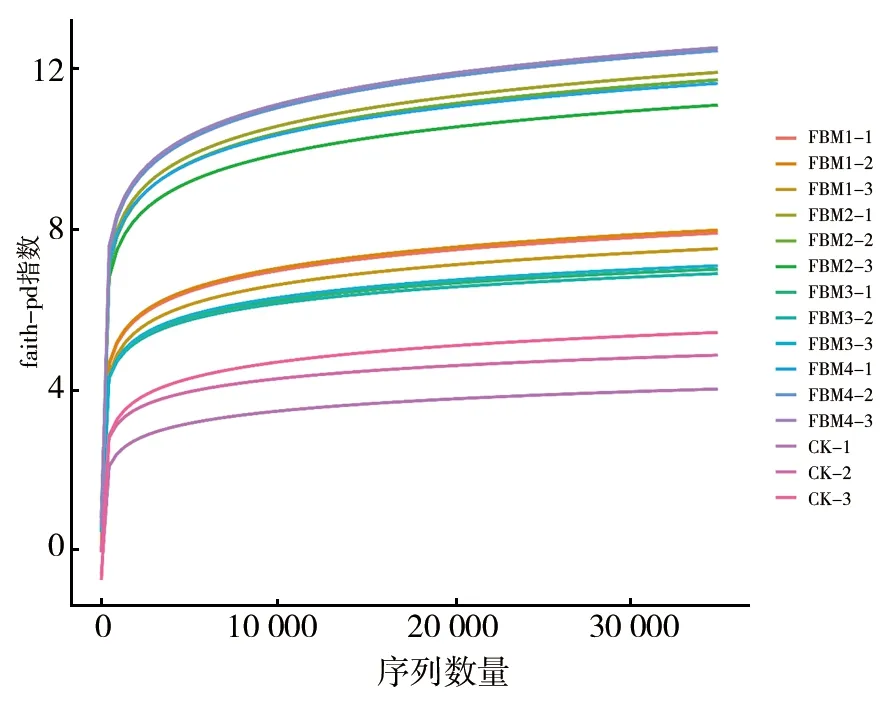
图1 基于faith-pd指数分析的各样品稀释曲线图 Fig.1 The seguencing results of bacterial diversity of fermened mattress materials
通过多样性指数分析,表1结果表明各样品均衡度较好,其中Chao1指数范围在124(FBM3)~227(FBM4),Observed OTU指数范围在127(FBM3)~211(FBM4)之间,4年期Chao1和Observed OTU指数较高,发酵床垫料细菌丰度最高,其次为2年期。 Shannon指数范围在3.599(FBM1)~4.675(FBM2),Simpson指数范围在0.787(FBM1)~0.888(FBM2),根据Shannon指数和Simpson指数反应样品细菌群落丰度,其中2年期指数最高,1年期指数最低,均高于对照组,其Shannon和Simpson指数分别为4.675和0.888。结果表明发酵床垫料中2年期细菌丰度最高,3年期最低。此外,四个年期各指数明显高于对照组,使用后的发酵床垫料细菌群落丰度高于原始发酵床垫料细菌丰度。
2.2 不同年期发酵床垫料细菌种群归类分析
共检测到15门,30纲,55目,117科,224属和278种细菌。OTU共有物种8种,特有物种中1年期OTU为90种,2年期141种,3年期96种,4年期164种,对照组22种。共有物种分别隶属于变形菌门 (Proteobacteria)、厚壁菌门 (Firmicutes)和拟杆菌门 (Bacteroidetes)。变形菌门主要含4个属:短波单胞菌属(Brevundimonas)、无色杆菌属(Achromobacter)、苍白杆菌属(Ochrobactrum)和一未知属;厚壁菌门3个属:芽胞杆菌属 (Bacillus)、赖氨酸芽胞杆菌属 (Lysinibacillus) 及一未知属;拟杆菌门2个属鞘氨醇杆菌属 (Sphingobacterium) 及一未知种。随着发酵床垫料使用时间延长,细菌特征种类也发生了变化,从1年期的90种到使用两年后增加到141种,三年后显著减少到96种,第四年闲置后上升到161种(图2)。

图2 样品共有及特有物种组成分析 Fig.2 Composition analysis of common and endemic species
2.3 不同年期发酵床垫料细菌门及科水平的优势种群分析
门分类水平微生物群落结构组成分析如图3所示,对照组优势菌门为厚壁菌门 (52.03%),其次为变形菌门 (40.28%),存在少量拟杆菌门。实验组中主要为厚壁菌门、变形菌门、拟杆菌门和放线菌门。其中1年期发酵床垫料厚壁菌门 (44.71%) 和变形菌门 (40.88%) 占主导地位,含少量拟杆菌门 (12.38%) 和放线菌门 (Actinobacteria,2.00%)。2年期细菌群落与1年期相比,数量增加且结构更为复杂,其中变形菌门68.57%、拟杆菌门16.01%、厚壁菌门9.94%及放线菌门2.01%,并出现绿弯菌门 (Chloroflexi,0.37%) 和疣微菌门 (Verrucomicrobia,0.30%)。3年期中细菌群落显著减少,其中变形菌门 (64.03%) 及厚壁菌门 (31.11%)为优势门。4年期发酵床垫料出现新分类群:疣微菌门 (1.08%)、蓝细菌门(Cyanobacteria) 与芽单胞菌门 (Gemmatimonadetes),变形菌门 (70.45%) 与拟杆菌门 (17.54%)为优势类群,含部分厚壁菌门 (6.53%) 与放线菌门 (3.00%)。总体变化为:2年期和4年期细菌群落丰度大于1年期和3年期,其中厚壁菌门在初始发酵床垫料中的含量随使用时间的增长显著降低,4年期时降低到6.53%(相对含量降低45.5%)。变形菌门群落结构与厚壁菌门呈相反趋势,初始发酵床垫料使用至4年期时变形菌门增加30.17%,且4年期细菌群落结构趋于复杂。拟杆菌门在1年期、2年期和4年期中较稳定存在,在对照组中仅有少部分存在。结果表明在不同时期发酵床垫料细菌群落有差异(图4)。在门群落水平上,有三个未明确分类的细菌差异显著,说明发酵床垫料中含有未被发掘的微生物资源。

图3 不同年期发酵床垫料物种组成分析(门水平)Fig.3 Analysis of species composition of fermented mattress materials in different ages (phylum level)
在科水平细菌群落组成中,对照组主要包括链球菌科 (Streptococcaceae) 和乳酸杆菌科 (Lactobacillaceae)。四个不同年期发酵床垫料主要含有芽胞杆菌科 (Bacillaceae)、柄杆菌科 (Caulobacteraceae)、布鲁氏菌科 (Brucellaceae)、鞘脂杆菌科 (Sphingobacteriaceae)、产碱杆菌科 (Alcaligenaceae)、丛毛单胞菌科 (Comamonadaceae) 和红杆菌科 (Rhodobacteraceae)。初始发酵床垫料使用一年后(1年期)细菌群落结构发生变化,由初始发酵床垫料中的链球菌科(79.26%)和猪皮杆菌科(20.03%)演替为芽胞杆菌科 (43.62%)、柄杆菌科(6.43%)、布鲁氏菌科(23.20%)、鞘脂杆菌科 (8.42%)和产碱杆菌科 (4.94%)。2年期主要为柄杆菌科(34.44%)、布鲁氏菌科(19.62%)、鞘脂杆菌科(12.27%)、芽胞杆菌科(8.73%)、丛毛单胞菌科(Comamonadaceae,5.73%) 和红杆菌科 (Rhodobacteraceae,4.18%)。3年期发酵床垫料主要有芽胞杆菌科(29.9%)、柄杆菌科(26.26%)、布鲁氏菌科(24.42%)、鞘脂杆菌科(1.82%)和产碱菌科 (8.25%)。4年期发酵床垫料优势科群落与3年期一致,其中柄杆菌科 (49.88%)和鞘脂杆菌科 (5.79%)含量增加,芽胞杆菌科(4.22%)、布鲁氏菌科(7.70%) 和产碱杆菌科(2.66%)减少。2年期中芽胞杆菌科 (8.73%)含量显著降低,于3年期中再次增加,在4年期时含量下降为4.22%。结果表明,不同使用期限的发酵床垫料细菌组成呈现曲线变化,主要细菌丰度有差异(图4)。
2.4 不同年期发酵床垫料优势细菌属的组成
选取丰度最高的前50个属OTUs代表序列进行系统进化分析,并绘制进化关系聚类树图(图5)。拟杆菌门鞘氨醇杆菌属(Sphingobacterium) 在1年期和2年期发酵床垫料中丰度较高,且与Parapedobacter和地杆菌属(Pedobacter)亲缘关系较近。黄杆菌属(Flavobacterium)与金黄杆菌属(Chryseobacterium)亲缘关系相近。黄杆菌属在3年期发酵床垫料中丰度较高;Parapedobacter与金黄杆菌属在4年期发酵床垫料含量较高;食冷菌属(Algoriphagus)在2年期发酵床垫料中丰度较高。厚壁菌门中梭菌属(Clostridium)与嗜碱菌属(Alkaliphilus)主要存在于1年期发酵床垫料中,2年期中少量存在;短芽胞杆菌属(Brevibacillus)与赖氨酸芽胞杆菌属(Lysinibacillus)存在于4年期发酵床垫料中;芽胞杆菌属(Bacillus)主要分布在1年期和3年期发酵床垫料中;乳酸杆菌属(Lactobacillus)与链球菌属(Streptococcus)在对照组中显著存在且两者系统发育亲缘关系较近。放线菌门中红球菌属(Rhodococcus)大量存在于4年期发酵床垫料中;纤维菌属(Cellulosimicrobium)在1年期及4年期中少量存在;白色杆菌属(Leucobacter)在1年期发酵床垫料中大量存在;微杆菌属(Microbacterium)于2年期发酵床垫料中少量存在,在4年期中丰度较高。不同年期发酵床垫料变形菌门细菌种类丰富,28个属OTUs代表形成两个分支。其中一分支主要分布于2~4年期发酵床垫料中。对氯苯胺属(Diaphorobacter)在1年期发酵床垫料显著存在。此外Hylemonella、寡养单胞菌属(Stenotrophomonas)、埃希氏菌属(Escherichia)、肠杆菌属(Enterobacte)、丛毛单胞菌属(Comamonas)和噬氢菌属(Hydrogenophaga)于1年期中少量存在,主要在2年期发酵床垫料中大量聚集;无色杆菌属(Achromobacter)、产碱杆菌属(Alcaligenes)及腾黄单胞菌属(Luteimonas)在3年期发酵床垫料中丰度显著高于其他年期发酵床垫料;假单胞菌属(Pseudomonas)和不动杆菌属(Acinetobacter)在4年期发酵床垫料中丰度较高,其次为2年期。第二分支在3年期发酵床垫料中丰度显著低于其他年期发酵床垫料。Pseudochrobacterum、Shinella、Defluvidbacter和Kaistia在1年期发酵床垫料中丰度较高,在2年期发酵床垫料中丰度较低;副球菌属(Paracoccus)、鞘氨醇盒菌属(Sphingopyxis)和枝动杆菌属(Mycoplana)于2年期发酵床垫料中显著存在;苍白杆菌属(Ochrobactrum)主要存在于3年期发酵床垫料中;土壤杆菌属(Agrobacterium)、柄杆菌属(Caulobacter)、德沃斯氏菌属(Devosia)和Balneimonas主要存在于4年期发酵床垫料中。属水平热图分析表明,不同年期样本中细菌组成丰度及结构不同。
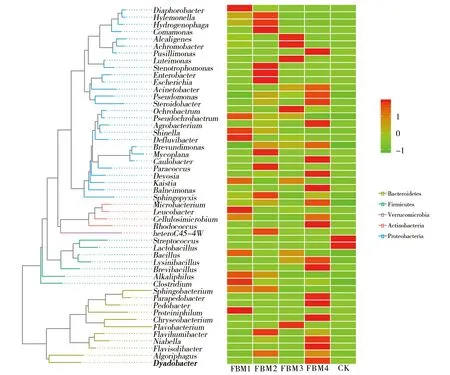
图5 系统进化及组间物种丰度分析(属水平)Fig.5 Phylogeny and species abundance analysis between groups (genus level)
2.5 不同年期发酵床垫料细菌功能基因预测
为探究不同年期发酵床垫料细菌组成结构差异与功能间的联系,本研究通过KEGG数据库进行相关功能预测。L1水平中新陈代谢 (metabolism, Mtbl)、环境信息处理 (Eenvironmental Information Processing, EnIP)、遗传信息处理 (Gebetic information Processiong, GnIP) 和未知分类功能 (unclassified, Uncl) 为不同年期发酵床垫料的主要功能,该水平不同年期发酵床垫料功能基因无显著差异。
基于PCA分析各等级功能基因在不同年期发酵床垫料中的主导能力(图6)。L3水平中初始发酵床垫料基因丰度主要为核糖体肽酶(Ribosome Peptidases, Rbsm)和转录因子(Transcription factors, Trnf);1年期发酵床垫料主要为氧化磷酸化作用(Oxidative phosphorylation, Othr),其次为ABC转运蛋白(ABC transporters, ABCt);2年期和4年期发酵床垫料主要为原核生物的碳固定途径(Carbon fixation pathways in prokaryotes, Cfpip)和乙醛酸和二羧酸的代谢(Glyoxylate and dicarboxylate metabolism , Gadm);3年期发酵床垫料主要为ABC转运蛋白功能。

图6 不同年期发酵床垫料细菌主导能力PCA分析图Fig.6 PCA analysis of bacterial dominance of bedding materials in different years
应用ANOVA+Duncan进行不同年期发酵床垫料L3水平各项功能基因丰度差异分析(图7)。ABC转运蛋白 (ABCt) 在3年期发酵床垫料中显著高于其他三个年期,其次为1年期,初始发酵床垫料显著最低。磷酸转移酶系统(Phosphotransferase system, PTS)在初始发酵床垫料显著高于四个年期发酵床垫料,其次1年期和2年期显著高于3年期和4年期。转运蛋白基因丰度对照组中显著高于1年期和3年期发酵床垫料,其次为2年期与4年期,两者间差异显著。细菌分泌系统 (Secretion system)、细菌毒素类 (Bacterial toxins ) 和调控系统 (regulatory system) 四个年期发酵床垫料均显著高于对照组,且细菌分泌系统在2年期发酵床垫料中显著最高,结果表明不同时期发酵床垫料中相应的细菌群落功能基因丰度有差异。

图7 不同年期发酵床垫料组间功能差异分析Fig.7 Functional differences between groups of fermented mattress materials of different ages
3 讨 论
本研究通过扩增不同年期发酵床垫料细菌V3~V4区域,采用Illumina Miseq二代测序技术进行四个不同年期发酵床垫料细菌组成和多样性分析,并对不同年期发酵床垫料进行功能预测,研究细菌群落与功能之间的联系。共获得924 008条序列,其中1年期至4年期有效序列均值分别为41 030、44 136、38 654和43 179,对照组最小为37 394;最小值为初始发酵床垫料对照组35 375,最大值为2年期发酵床垫料48 150。检测到共有物种OTU 8种,特有物种中丰度最高为4年期发酵床垫料。研究发现发酵年限不同会影响发酵床垫料细菌组成与结构,多样性分析表明4年期发酵床垫料的细菌丰度最高,其次为2年期。2年期细菌种类最多,3年期最少。此外,四个年期各指数分别显著高于对照组,并且样品组间差异较大。
物种多样性分析表明不同年期发酵床垫料中厚壁菌门、放线菌门、拟杆菌门及变形菌门为优势门群落,此结果与相关研究[16-17]结论一致。在微生物发酵床垫料与猪仔粪污形成一个发酵系统时,细菌含量会逐渐变化。随着发酵垫料的使用,发酵床垫料中细菌群落趋于复杂,2年期发酵床垫料与1年期发酵床垫料细菌组成相比,2年期细菌丰度增加且结构更加复杂,出现绿弯菌门和疣微菌门。随着时间延长,3年期发酵床垫料细菌群落减少,其中拟杆菌门与放线菌门含量仅为3.55%和1.29%。4年期发酵床垫料细菌群落结构出现新的分类群:疣微菌门、蓝细菌门和芽单胞菌门。疣微菌门在2年期发酵床垫料中出现,使用一年后其含量降低至消失,发酵床垫料闲置后该分类群再次出现。
不同发酵年期发酵床垫料中起主要降解作用的细菌不同,1年期发酵床垫料中主要细菌为鞘氨醇杆菌属 (Sphingobacterium)、梭菌属(Clostridium)、嗜碱菌属 (Alkaliphilus)、白色杆菌属 (Leucobacter)、芽胞杆菌属 (Bacillus)、短芽胞杆菌属 (Brevibacillus)、赖氨酸芽胞杆菌属 (Lysinibacillus)、Pseudochrobacterum、Shinella、Defluvidbacter和Kaistia,该时期病原菌丰度较低。鞘氨醇杆菌属隶属拟杆菌门鞘氨醇杆菌科,分布广泛,在堆肥[18]、污泥[19]、植物[20]及动物[21]中均存在,多具有降解特性。梭菌属、嗜碱菌属及芽胞杆菌属是堆肥中的常见菌,其中芽胞杆菌能够降解发酵床垫料有机质,参与猪粪降解与发酵床病原生防过程,在发酵中起重要作用[22-23]。而大量存在的Pseudochrobacterum、Defluvidbacter及Kaistia,可能具有特定功能,进行相关功能研究可获得更广泛的生物资源。2年期发酵垫料中食冷菌属 (Algoriphagus)、微杆菌属 (Microbacterium)、Hylemonella、寡养单胞菌属 (Stenotrophomonas)、埃希氏菌属 (Escherichia)、肠杆菌属 (Enterobacte)和假单胞菌属 (Pseudomonas) 含量较高。值得注意的是随着发酵床垫料的使用,该时期病原菌埃希氏菌属及肠杆菌属等致病菌大量聚集。研究表明食冷菌属这一新物种对多种抗生素具有抗性[24];生物防治潜能及机理研究发现微杆菌属菌株对青霉菌具有较强的抵抗作用,并与甲硫氨酸以及异戊二烯等诱导抗性相关物质合成单萜类基因[25];寡养单胞菌属部分菌株抵抗多重耐药病原体的作用在农业、工业、医药领域,表现出良好的应用研发前景[26]。假单胞菌属隶属于假单胞菌科,假单胞菌科分布广泛,其中许多细菌具有很强的降解油脂、烷烃、酚类和有机氮[27-28]的能力。该时期还有大量丛毛单胞菌属 (Comamonas)、噬氢菌属 (Hydrogenophaga)、副球菌属 (Paracoccus)、鞘氨醇盒菌属 (Sphingopyxis) 及枝动杆菌属 (Mycoplana)等具有降解特性[29-30]的有益细菌,供发酵床正常运行。随着发酵床垫料的使用,降解类细菌丰度发生变化。3年期发酵床垫料中,芽胞杆菌属、黄杆菌属 (Flavobacterium)、无色杆菌属 (Achromobacter)、产碱杆菌属、腾黄单胞菌属及苍白杆菌属为主要降解菌。同2年期相比,3年期发酵床垫料中的细菌群落结构较为简单,原因可能为随着使用年份的增加,部分菌株不再适应发酵床系统环境而被淘汰。2年期发酵床垫料中大量存在的致病菌丰度降低,可见发酵床的使用可以减少病原菌的滋生,该结果与陈倩倩等[31]对福建一养殖场发酵床细菌多样性研究结果一致,表明发酵床可以抑制致病菌生存,保证家禽健康。有研究表明产碱杆菌属的粪产碱菌 (Alcaligenesfaecalis) 可抑制大肠埃希菌和金黄色葡萄球菌 (Staphylococcusaureus) 等病原菌[32],这对揭示发酵床技术对病原菌控制具有防治作用和启发意义。该时期芽胞杆菌属丰度在3年期中依然极高,表明该属菌属适用于发酵床系统。4年期发酵床垫料中红球菌属、金黄杆菌属、短芽胞杆菌属 (Brevibacillus)、赖氨酸芽胞杆菌属 (Lysinibacillus)、假单胞菌属、不动杆菌属(Acinetobacter)、土壤杆菌属 (Agrobacterium)和柄杆菌属 (Caulobacter) 显著存在。由于该年期发酵床垫料已露天闲置,其微生物群落趋于复杂可能与环境有关。因3年期发酵床垫料处于一个致病菌降解的关键时段,到第四年发酵床致病菌丰度降低,细菌群落处于一个较好的环境。因此,可以通过挖掘高年期发酵床垫料中土著微生物应用于发酵床养殖技术中,以提高地区发酵床养殖效益。
为探究不同年期发酵床垫料中细菌功能差异,本研究初步进行功能预测。随着使用时间的增长,发酵床垫料内部物质结构发生变化[33],可能导致不同年期发酵床垫料行使功能有差异。研究结果表明,KEGG通路L1水平中各项功能基因丰度无显著差异。通过L3水平中基因丰度较高的环境信息加工功能差异分析表明,对照组初始发酵床垫料中转运蛋白及磷酸转移酶系统功能基因丰度显著高于已发酵的四个年期发酵床垫料。初始发酵床垫料中细菌种类较少,且与其他四个年期发酵床垫料相比较无特殊发酵环节,不与猪仔构成系统环境,此时对于系统调节及细菌分泌系统等功能的行使不显著,则相关基因丰度较低。对照组中细菌主要进行有机物质转换用于维系自身生命进程,根据物种变化,推测乳酸杆菌属与链球菌属在初始发酵床垫料中显著存在可能主导转运蛋白及磷酸转移酶系统相关功能的进行;1年期和2年期发酵床垫料中猪仔肠道粪污的排放及病原菌产生导致磷酸转移酶系统介导应激反应,调节粪污中碳氮物质代谢来调控发酵床的运行。ABC转运蛋白、细菌分泌系统、细菌毒素类及调控系统在四个年期发酵床垫料中均显著高于对照组。ABC转运蛋白可转运的底物包括有机离子、单糖、聚糖、胆固醇、磷脂、氨基酸、肽、蛋白质、毒素、药物、抗生素和异源物质等[34]。使用发酵床可降低病原菌的产生[3,35]的结果与该研究一致。发酵床垫料中埃希氏菌属及肠杆菌属丰度逐渐降低,推测发酵床可以降低病原菌生存,可能与发酵床中微生物拮抗作用有关,其中产碱杆菌属可能对病原菌抑制具有重要作用。并且随着粪污排泄、有机质降解以及细菌毒素的产生,导致发酵床微生物细菌分泌系统功能激活,控制向外分泌蛋白质,进行各级分泌系统调控,使发酵床正常发挥作用。
综上,本研究利用高通量测序技术,分析不同年期发酵床垫料细菌多样性。四个年期发酵床垫料中微生物数量及种类差异显著,起主要降解作用的细菌不同。1年期和3年期发酵床垫料细菌丰度及种类低于2年期和4年期发酵床垫料。其次发酵床垫料中微生物主要以变形菌门、厚壁菌门、放线菌门及拟杆菌门为主。1年期中起主要降解作用的是鞘氨醇杆菌属、梭菌属、嗜碱菌属、白色杆菌属、芽胞杆菌属、短芽胞杆菌属及赖氨酸芽胞杆菌属;2年期发酵床垫料中食冷菌属、微杆菌属、Hylemonella、寡养单胞菌属 (Stenotrophomonas)、丛毛单胞菌属、噬氢菌属 (Hydrogenophaga)、副球菌属、鞘氨醇盒菌属和假单胞菌属为主要降解菌;3年期发酵床垫料中芽胞杆菌属、黄杆菌属、无色杆菌属、产碱杆菌属,腾黄色单胞菌属及苍白杆菌属为主要降解菌;4年期发酵床垫料中红球菌属、金黄杆菌属、短芽胞杆菌属、赖氨酸芽胞杆菌属、假单胞菌属、不动杆菌属、土壤杆菌属和柄杆菌属显著存在。功能预测与物种分析表明初始发酵床垫料显著存在的乳酸杆菌属与链球菌属主要进行转运蛋白及磷酸转移酶系统功能。四个年期发酵床垫料中ABC转运蛋白、细菌分泌系统、细菌毒素类及调控系统为主要功能。研究表明,不同发酵年期有关降解类有益细菌丰度、组成及功能存在差异,而且随着发酵床垫料的使用,病原菌数量逐渐降低,因此发酵床养殖对猪仔生长具有积极作用。同时细菌群落组成与功能相适应,不同发酵时期发酵床垫料中细菌组成差异可能导致功能变化,这对发酵床正常运行和猪仔健康生长具有指导意义。
猜你喜欢
疯狂英语·新读写(2023年7期)2023-08-19
河南医学研究(2022年19期)2022-10-19
中国生物防治学报(2022年3期)2022-07-09
微生物学杂志(2021年2期)2021-07-01
微生物学杂志(2020年2期)2020-12-31
生态学报(2019年11期)2019-07-08
科教新报(2018年4期)2018-10-21
小朋友·快乐手工(2017年5期)2017-07-11
时代英语·高一(2016年5期)2016-10-12
食品工业科技(2014年23期)2014-03-11
