
一株可裂解动物源性多重耐药志贺氏菌的噬菌体分离鉴定及其生物学特性研究
2023-12-06 02:29徐美余邓征宇罗成莹邓先余林连兵
微生物学杂志 2023年5期
徐美余, 张 瑶, 邓征宇, 何 秀, 罗成莹, 邓先余, 林连兵*, 王 峰*
(1.昆明理工大学 生命科学与技术学院, 云南 昆明 650500; 2.云南省高校饲用抗生素替代技术工程研究中心, 云南 昆明 650500)
志贺氏菌(Shigella)属肠杆菌科(Enterobacteriaceae)的志贺氏菌属(Shigella),革兰染色阴性,为人类传播细菌性痢疾中最普遍的食源性病原菌,严重危害人类健康。受卫生条件限制,志贺氏菌病主要发生在发展中国家,Jain等[1]和Shang等[2]研究发现,急性肠胃炎和腹泻样本中志贺氏菌的检出率高达4.2%。研究表明主要由志贺氏菌引起的重症溃疡性结肠炎表现为严重的菌群失调。志贺氏菌食物中毒是指由志贺氏菌所致的细菌性食物中毒,志贺氏菌侵袭人类肠道黏膜组织并释放大量内毒素引起症状,可导致肠道感染引起人尤其是幼儿腹泻,潜伏期一般10~14 h,其毒力主要依赖于Ⅲ型分泌系统分泌的效应因子[3]。志贺氏菌引起的腹泻是一种主要的公共卫生威胁,可通过粪口传播于人与人之间,感染的高发率主要发生在5岁以下儿童,且志贺氏菌对抗生素已产生较强耐药性,尤其是对阿莫西林和四环素耐药性极高[4-5]。噬菌体(Bacteriophage或Phage)是种类极其丰富的一类细菌病毒,噬菌体通常在充满细菌群落的地方广泛分布,感染力强,常以指数数量级增殖[6]。在形态上可分为丝状噬菌体、有尾和无尾的二十面体噬菌体和有脂质包膜的噬菌体[7]。噬菌体具有明显的宿主特异性,相对容易分离,并可进行遗传修饰。噬菌体是细菌的天然捕食者,裂解性噬菌体感染后可杀灭宿主菌。研究表明,利用噬菌体制备抗菌剂,可安全高效治疗耐药性细菌感染,例如,Alvi等[8]发现噬菌体对感染多重耐药铜绿假单胞菌的小鼠有治疗效果。其还可用于治疗小牛腹泻以及严重的儿童痢疾,能取得良好效果且具有很好的安全性[9-10]。另外,研究显示,噬菌体可替代抗生素应用于水产养殖行业中[11]。上述研究表明,噬菌体在农业、养殖业和畜牧业中具有良好的应用前景。自上世纪四十年代在临床实践中引入抗生素以来,抗生素在防治人类和动物细菌感染疾病中发挥着重要作用[12]。但近年来由于抗生素的使用不规范,各种耐药菌株,如泛耐药性“超级细菌”在全球不断出现,对全球公共卫生和国民健康造成严重威胁。据世界卫生组织预测,如果没有新的抗菌剂被发现,到2050年人类将会回到青霉素之前的时代。因此,探索新的抗生素替代品,研发抗生素替代技术迫在眉睫[13]。作为可侵袭、裂解细菌的天然武器,对于抗生素耐药细菌导致的感染,基于噬菌体的治疗策略是一个很好的选择。本研究从广西富凤鸡场病鸡肠道黏膜中分离出26株志贺氏菌。以其中的多重耐药性志贺氏菌BS26作为宿主菌,分离纯化得到一株新型的裂解性噬菌体PSF26,并研究其生物学特性,以期为耐药性志贺氏菌的噬菌体治疗及饲用抗生素替代实践提供参考。
1 材料与方法
1.1 材料
1.1.1 菌株来源 实验肉鸡源自广西富凤鸡场,黏膜取自病鸡肠道、粪便取自健康肉鸡。宋内志贺氏菌(Shigellasonnei)CMCC(B)51592、痢疾志贺氏菌(Shigelladysenteriae)CMCC(B)51105、大肠埃希菌(Escherichiacoli)CMCC(B)44102、福氏志贺氏菌(Shigellaflexneri)CMCC(B)51572、金黄色葡萄球菌(Staphylococcusaureus)ATCC6538、铜绿假单胞菌(Pseudomonasaeruginosa)ATCC27853和BNCC139675、大肠埃希菌(Escherichiacoli)O157、肠炎沙门氏菌(Salmonellaenteritidis)CMCC(B)50335、金黄色葡萄球菌(Staphylococcusaureus)1606BL1486、枯草芽胞杆菌(Bacillussubtilis)CMCC(B)63501、无乳链球菌(Streptococcusmastitidis)SAM12和沙门氏菌(Salmonella)CMCC(B)50094均为昆明理工大学生命科学与技术学院噬菌体与肠道微生物课题组保藏菌种。
1.1.2 培养基 LB液体培养基:NaCl 10 g,蛋白胨10 g,酵母粉5 g,去离子水1 000 mL;LB半固体培养基:在LB液体培养基基础上加7.5 g/L琼脂;LB固体培养基:在LB液体培养基基础上加15 g/L琼脂;PBS缓冲液:A液为NaH2PO431.2 g,1 000 mL容量瓶定容,B液为Na2HPO471.63 g,1 000 mL容量瓶定容;取A液28 mL、B液72 mL混合,加入100 mL去离子水稀释,用1 mol/L HCl调节pH至7.4;SM缓冲液:NaCl 5.8 g,MgSO42.0 g,1 mol/L Tris-HCl (pH 7.5) 50 mL,2%明胶5 mL,加去离子水至1 000 mL。
1.1.3 主要试剂与仪器设备 细菌基因组提取试剂盒(DP201101X),天根生化科技(北京)有限公司;志贺氏菌选择琼脂培养基,广东环凯微生物科技有限公司;细菌通用引物27F和1492R;2×TaqMaster Mix缓冲液;磷钨酸;0.22 μm滤膜(天津市津腾实验设备有限公司);DNase I、RNase A(上海翊圣生物科技有限公司);空白药敏纸片(直径6.0 mm,厚度1.0 mm);PEG8000(Bio Froxx公司)。高低温恒温振荡培养箱(HZQ-F160A,上海一恒科学仪器有限公司);恒温培养摇床(THZ-100, 上海一恒科学仪器有限公司);高速冷冻离心机(TGL-23,四川蜀科仪器有限公司);透射电子显微镜(JEM-1400,上海铸金分析仪器有限公司)。
1.2 方法
1.2.1 志贺氏菌的分离鉴定 在超净工作台刮取病鸡肠道黏膜溶于无菌水,梯度稀释至10-5,取100 μL涂布于LB固体培养基上,37 ℃倒置培养12 h。分别挑取不同形态单菌落于LB液体培养基中,37 ℃、150 r/min振荡培养24 h,采用四分体划线法在LB固体培养基上划线,37 ℃倒置培养12 h,得到纯化的单菌落。将纯化的单菌落接种至LB液体培养基中,培养至对数期(OD600约为0.8),按照细菌基因组DNA提取试剂盒操作说明提取细菌DNA。使用细菌通用引物27F (5′-AGAGTTTGATCCTGGCTCAG-3′)和1492R(5′- GGTTACCTTGTTACGACTT -3′)对分离株进行16S rRNA基因扩增。PCR反应体系(20 μL): 2×TaqMaster Mix缓冲液10 μL, DNA模板1 μL,上、下游引物(10 μmol/L)各1 μL, ddH2O 7 μL。PCR反应条件:94 ℃ 1 min; 94 ℃ 50 s, 56 ℃ 45 s, 72 ℃ 1 min, 30个循环;72 ℃ 10 min, 4 ℃保存。取PCR产物5 μL,用2%琼脂糖凝胶电泳检测。将检测后PCR产物送至生工生物工程(上海)股份有限公司测定16S rRNA基因序列,测序结果在NCBI中进行BLAST比对分析。通过Mega X构建志贺氏菌菌株BS26的遗传进化树。
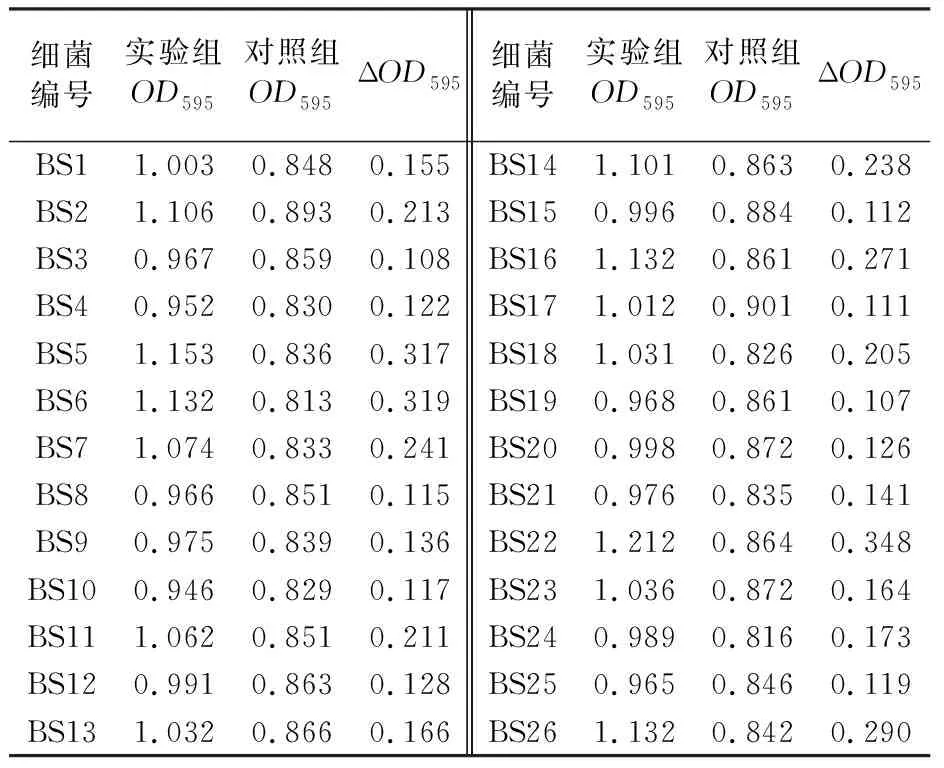
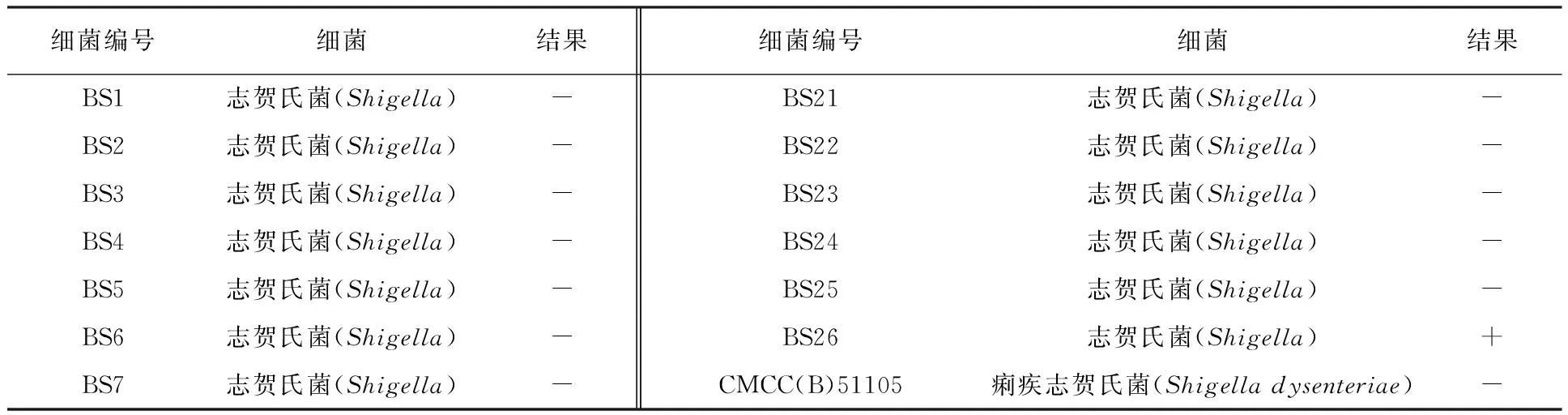
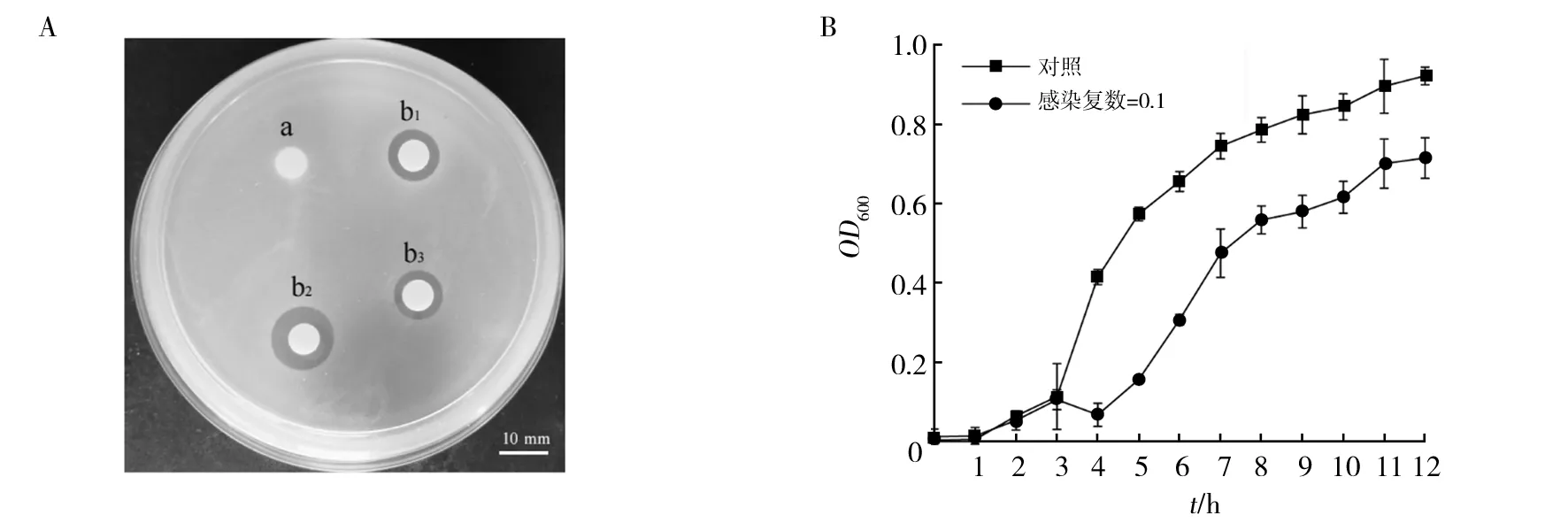
1.2.2 志贺氏菌耐药性测定 将菌株BS26培养至对数期,取100 μL BS26菌液于4 mL的EP管中,倒入灭菌后不烫手的LB半固体培养基,迅速混匀后倒至LB固体培养基平板上;用镊子夹取药敏纸片贴于LB半固体培养基表面,37 ℃倒置培养12 h;观察药敏纸片周围是否出现抑菌圈,并测量抑菌圈的直径(d)大小。抑菌圈直径d>20 mm为高度敏感;10 mm 1.2.3 噬菌体的分离纯化 从健康肉鸡粪便中分离噬菌体,将50 g健康的肉鸡粪便和10 mL 26株志贺氏菌分离株加入300 mL LB液体培养基中,37 ℃、150 r/min振荡培养48 h。培养液经13 000 r/min离心10 min,用0.22 μm滤膜过滤得到上清即为混合噬菌体液。噬菌体裂解菌体后溶液中的蛋白含量会增高,可以采用考马斯亮蓝染色检测蛋白含量的方法分离噬菌体。在26株志贺氏菌分离株菌液中分别加入100 μL噬菌体液,并以等量ddH2O为对照,37 ℃、150 r/min培养12 h,然后10 000 r/min离心1 min后取上清液,按照V样品∶V考马斯亮蓝染色液∶VddH2O=1∶1∶3测定OD595。相比于对照组,将OD595上升10%的噬菌体用于纯化鉴定。以菌株BS26为宿主,利用双层平板法对噬菌体进行纯化。取100 μL噬菌体液、900 μL ddH2O混合均匀进行梯度稀释,将100 μL稀释后的噬菌体液与100 μL宿主菌加入4 mL EP管中,然后倒入4 mL冷却至约50 ℃的LB半固体培养基,混匀后迅速倒入LB固体培养基中。37 ℃倒置培养12 h;选择分布均匀且完整的噬菌斑,接种到对数期的宿主菌菌液中,在37 ℃、150 r/min条件下培养5~8 h使噬菌体增殖;将培养后的混合液13 000 r/min离心10 min后取上清液,上清液用0.22 μm滤膜过滤,分装于灭菌后的EP管中。重复上述步骤,直到得到单一种类的噬菌体,将纯化后的噬菌体液放入4 ℃冰箱备用。 1.2.4 噬菌体形态观察 ① 噬菌体浓缩:将1 L噬菌体液17 000 r/min离心15 min取上清液,然后加入DNase I、RNase A (终质量浓度均为1 μg/mL),37 ℃恒温水浴锅中消化3 h,再加入NaCl(终浓度为1 mol/L),NaCl溶解后冰上放置2 h;然后17 000 r/min离心15 min取上清液,加入PEG8000 (终质量分数为10%),溶解后于4 ℃冰箱放置12 h;所得溶液于16 000 r/min离心20 min后收集沉淀,用12 mL SM缓冲液溶解,放置室温1 h;使用12 mL氯仿抽提1次,以除去PEG8000,之后10 000 r/min离心15 min后收集水相,采用双层平板法测定浓缩噬菌体液的效价并将样品保存于4 ℃冰箱[14]。② 透射电镜观察:吸取20 μL用PEG沉淀法制备的浓缩噬菌体液滴在复膜铜网上,室温自然沉淀15 min;在样品未完全干燥时,用2%磷钨酸(pH=7.0)滴染2 min,吸取多余的染液。常温放置2 min后,使用JEM-1400透射电子显微镜进行观察,将BS26对应的噬菌体,命名为PSF26。 1.2.5 噬菌体的生物学特性研究 ① 宿主谱鉴定:为了测定噬菌体PSF26的宿主范围以及宿主特异性,用26株志贺氏菌分离株BS1~BS26,以及13株标准菌株作为宿主,采用双层平板法对纯化后的噬菌体PSF26进行敏感性试验。取100 μL待测菌液于4 mL EP管中,然后倒入4 mL冷却至约50 ℃的LB半固体培养基,混匀后迅速将混合物倒入LB固体培养基中;吸取10 μL噬菌体液,滴在双层琼脂平板的上层,放置于超净工作台中风干后,倒置37 ℃恒温培养12 h。观察是否有透明噬菌斑出现。每组各三个平行重复。② 噬菌体PSF26最佳感染复数(Multiplicity of Infection, MOI)测定:将宿主菌株BS26接种于LB液体培养基中,37 ℃、150 r/min条件下培养至OD600值为0.6~0.8,梯度稀释后利用平板涂布法进行计数;对噬菌体液进行梯度稀释,将宿主菌液和噬菌体液按照感染复数为0.001、0.01、0.1、1、10、100的比例混合;使混合液的总体积为5 mL,少于5 mL则用LB液体培养基补足;之后于37 ℃、150 r/min恒温培养4 h,将培养后的混合液13 000 r/min离心15 min后取上清液,用0.22 μm滤膜过滤;取不同感染复数的上清液100 μL进行梯度稀释,并通过双层平板法验证噬菌体的效价。验证后效价最高的一组即为噬菌体PSF26的最佳感染复数。每组各三个平行重复。③ 一步生长曲线:噬菌体的一步生长曲线是定量描述噬菌体生长规律的实验曲线。具体操作如下:将宿主菌BS26接种于LB液体培养基中,37 ℃、150 r/min条件下培养至OD600值为0.6~0.8;以噬菌体PSF26的最佳感染复数加入噬菌体PSF26,并放置于37 ℃吸附15 min;之后11 000 r/min离心10 min后留沉淀,用LB液体培养基轻轻吹打混匀,离心留沉淀。沉淀中加入LB液体培养基30 mL,37 ℃、150 r/min恒温培养;每隔15 min取样 一次,直到150 min停止取样。通过双层平板法验证不同时间点噬菌体的效价。每组各三个平行重复。④ 噬菌体抗菌效果评估:采用空白药敏纸片法评估噬菌体PSF26对宿主菌BS26的抗菌效果。在平板中倒入适量LB固体培养基,用于制备双层琼脂平板的下层培养基,风干凝固后备用。取100 μL BS26对数期菌液于4 mL EP管中,在EP管中倒入4 mL左右LB半固体培养基,上下颠倒混匀后迅速将混合物倒入准备好的LB固体培养基中,放置空白药敏纸片,以LB液体培养基为空白对照,在空白药敏纸片上滴加30 μL噬菌体原液(1011pfu/mL),37 ℃恒温培养12 h,并观察透明圈。每组各三个平行。将宿主菌株BS26接种于LB液体培养基中,37 ℃、150 r/min培养至OD600值为0.6~0.8;以噬菌体PSF26的最佳感染复数加入噬菌体PSF26,并建立无噬菌体的阴性对照,37 ℃吸附15 min后,37 ℃、150 r/min条件下培养,每隔1 h,以LB液体培养基为空白对照,测600 nm处的OD值。每组各三个平行。⑤ pH和热稳定性分析:为了评估噬菌体PSF26在不同酸碱度下的稳定性,使用NaOH与HCl溶液调节SM缓冲液pH值。pH值梯度设置为2、3、4、5、6、7、8、9、10、11、12、13。取100 μL噬菌体液与900 μL不同pH的SM缓冲液混合均匀,37 ℃恒温孵育2 h,通过双层平板法验证不同pH值时噬菌体的效价。为了评估噬菌体PSF26的稳定性,温度梯度设置为4、20、30、40、50、60、70、80 ℃。取PBS缓冲液900 μL于1.5 mL EP管中,将EP管于不同温度下预处理30 min。在各EP管中加入100 μL噬菌体液,继续于不同温度下培养1 h。通过双层平板法验证不同温度下噬菌体的效价。每组各三个平行重复。 本研究共分离到26株菌株,并分别命名为BS1~BS26。在NCBI中利用BLAST在线工具对细菌的16S rRNA基因序列进行相似性比对,BS1~BS26均为志贺氏菌分离株[15]。遗传进化树结果表明BS26与ATCC 29903亲缘关系最近,鉴定为福氏志贺氏菌(Shigellaflexneri)。菌株BS26的分子量大小为1 500 bp (图1)。 图1 菌株BS26的鉴定结果Fig.1 Identification results of Shigella BS26A:菌株BS26的16S rRNA基因扩增图;1:D2000 plus DNA Ladder (100~5 000 bp);2:BS26;B:菌株BS26的系统发育树A:Amplification of 16S rRNA gene of Shigella BS26;1:D2000 plus DNA Ladder (100-5 000 bp);2:Shigella BS26;B: Phylogenetic tree of Shigella BS26 耐药性实验结果显示菌株BS26对氨苄西林、四环素、氯霉素、磺胺甲噁唑、阿莫西林完全耐药,无抑菌圈出现(表1)。其对庆大霉素、氨曲南、头孢噻肟、头孢曲松等抗生素较为敏感,抑菌圈直径均大于20 mm。但对阿米卡星、链霉素、头孢他啶等只有中低度敏感,抑菌圈直径在10~20 mm内。此结果表明志贺氏菌株BS26已经出现多重耐药性,如果任由抗生素继续滥用,后续耐药性可能会更加严重。这也表明探索、发展新的可以替代抗生素的对治疗细菌感染方法的紧迫性和重要性。 表1 志贺氏菌BS26耐药性试验Table 1 Drug resistance test of Shigella BS26 实验结果发现实验组在加入噬菌体液12 h后,实验组的菌液与对照组相比明显变澄清,使用考马斯亮蓝法染色后发现在较为澄清的菌液中,蛋白质含量OD595相较于对照组出现明显升高(表2)。挑取OD595明显升高且菌液变澄清的菌种进行双层验证试验,经过多次纯化后共得到26株噬菌体。将菌株BS26对应的噬菌体命名为噬菌体PSF26。 表2 实验组与对照组蛋白含量OD595对比Table 2 Comparison table of protein content OD595 between experimental group and control group 用双层平板法对噬菌体PSF26进行纯化后,噬菌斑呈透明圆形且大小均一,边缘清晰,无晕环。说明噬菌体PSF26纯化成功(图2)。 图2 噬菌体PSF26的纯化结果Fig.2 Purification results of phage PSF26 通过透射电镜观察(图3),噬菌体PSF26的头部是二十面体结构,直径约为61 nm;具有非收缩性可弯曲的尾部,长约140 nm;尾部有尾鞘,具有收缩性,根据形态将其归为有尾噬菌体目(Caudovirales),长尾噬菌体科(Siphoviridae)[16]。 图3 噬菌体PSF26的电镜照片Fig.3 Electron micrograph of phage PSF26 2.5.1 噬菌体PSF26宿主谱分析 采用双层平板法对纯化后的噬菌体PSF26进行敏感性试验。如表3所示,在待测的26株志贺氏菌分离株BS1~BS26和13株标准菌株中,噬菌体PSF26在志贺氏菌分离株BS8、BS9、BS13、BS26的双层平板中出现清晰且透明的噬菌斑,直径大小约为20 mm,而13株标准菌株及其余志贺氏菌分离株均无噬菌斑形成。该结果表明,噬菌体PSF26对志贺氏菌存在一定的宿主特异性,但其对其中的福氏志贺氏菌和宋内志贺氏菌分离株均有裂解作用。 表3 噬菌体PSF26的宿主谱分析Table 3 Host spectrum analysis of phage PSF26 2.5.2 噬菌体PSF26最佳感染复数 如图4所示,在感染复数为0.1时,噬菌体PSF26的效价最高,达2.20×109pfu/mL。因此,可确定噬菌体PSF26的最佳感染复数为0.1。 图4 噬菌体PSF26最佳感染复数Fig.4 The optimal multiplicity of infection (MOI) of phage PSF26 2.5.3 一步生长曲线的测定 在感染宿主菌BS26后的75 min内,噬菌体PSF26的效价变化不明显,表明此时间范围为其潜伏期;在感染后75~90 min内噬菌体的效价急剧升高,表明其爆发期约为15 min;之后趋于平稳,表明噬菌体PSF26的裂解期时长为90 min,裂解量为(58±17) pfu/cell; 在90 min后进入稳定期(图5)。 图5 噬菌体PSF26的一步生长曲线Fig.5 One-step growth curve of phage PSF26 2.5.4 抑菌圈分析 噬菌体PSF26在LB固体培养基上的抑菌效果如图6所示,可见边缘清晰的圆形抑菌圈,抑菌圈直径为(11.33±1.53) mm,和对照相比(6 mm),显著性P值小于0.05(图6A)。进一步分析噬菌体PSF26的抑菌时间曲线,和对照相比,噬菌体PSF26从3 h起开始发挥作用,肉眼可以观察到试管变澄清,添加噬菌体PSF26后,菌株BS26生长速度明显低于对照组(图6B)。 图6 噬菌体PSF26对菌株BS26的抑菌效果Fig.6 Bacteriostatic effect of phage PSF26 on Shigella BS26A:噬菌体PSF26对菌株BS26的抑菌平板图;a:LB液体培养基对照;b1、b2、b3:噬菌体PSF26对菌株BS26的抑菌圈直径的三个平行;B:噬菌体PSF26对菌株BS26的抑菌时间曲线A: Bacteriostatic plate diagram of phage PSF26 against Shigella BS26; a is the LB liquid medium control;b1, b2 and b3 represent three parallel diameters of bacteriostatic circle of phage PSF26 against Shigella BS26; B: Bacteriostasis time curve of phage PSF26 against Shigella BS26 2.5.5 pH和热稳定性分析 噬菌体PSF26的效价在pH 2~3 (极酸)和13 (极碱)时为0,表明噬菌体在这些pH条件下处于失活状态;在pH 5~8时,噬菌体处于高活性状态;在pH为8~10时,噬菌体效价稍有降低,但幅度较小,依然可以维持在较高数量级。这些结果表明,噬菌体PSF26在较宽的pH范围内依然可以保持抗菌活性,说明其对酸碱有一定的耐受性(图7A)。如图7B所示,温度为4~40 ℃时,噬菌体PSF26活性较高,其效价维持在较高的数量级;温度为40~80 ℃时,噬菌体效价逐渐降低,至80 ℃时效价为0,完全丧失活性。因此,噬菌体PSF26的适宜作用温度为4~40 ℃,这也是常规环境温度和大部分细菌的适宜生长温度。 图7 噬菌体PSF26的pH (A) 和热稳定性 (B) 分析Fig.7 pH (A) and thermal stability (B) analysis of phage PSF26 志贺氏菌为典型的食源性可致肠道感染病原体,对养殖业和人体健康具有很大的危害性[17]。本研究从养鸡场病鸡肠道中分离得到26株志贺氏菌,通过志贺氏菌分离株致病性评估,发现菌株BS26可导致肉鸡出现腹泻症状[15],通过筛选、纯化得到一株裂解性噬菌体PSF26,鉴定证明噬菌体PSF26属于有尾噬菌体目,长尾噬菌体科。纯化浓缩后的噬菌体液效价为1.65×1011pfu/mL,高于近期报道的噬菌体BLCC16-001(2.0×1010pfu/mL)[18]和噬菌体vB-Sau S-SAP3 (109pfu/mL)[19]效价,表明噬菌体PSF26对病原菌裂解效果较好。同时,相比于肠出血性大肠埃希菌O157噬菌体(爆发时间为30 min)、鼠伤寒沙门氏菌噬菌体(爆发时间为180 min)、福氏志贺氏菌噬菌体(爆发时间45 min)[20-21],噬菌体PSF26的爆发时间为15 min,具有爆发时间短的优势。 噬菌体PSF26对志贺氏菌分离株BS8、BS9、BS13、BS26均有裂解作用(其中BS9为宋内志贺氏菌,BS8、BS13及BS26为福氏志贺氏菌),但不能裂解其他种类细菌,说明该噬菌体对志贺氏菌具有较强的宿主专一性,这与噬菌体vB-EcoP-E21具有一定的相似性[22]。该结果也表明,噬菌体PSF26在实践应用中,可具有良好的靶向性,不会对周围环境中其他益生性细菌造成杀灭,从而具有很好的安全性[23]。 通过对噬菌体PSF26的抑菌特性进行探究,发现噬菌体PSF26对多重耐药性志贺氏菌BS26的抑菌圈直径为(11.33±1.53) mm,其抑菌效果优于噬菌体EW3-f4 (抑菌圈直径9.75 mm)[24]。在液体培养基中,以未加入噬菌体PSF26的菌株BS26培养物为对照,通过测定OD600值评估噬菌体PSF26对菌株BS26的生长抑制作用,发现添加低浓度噬菌体PSF26后,菌体生长速度降低,明显低于对照组。基于此,针对食源性志贺氏菌污染,可进一步探索噬菌体PSF26作为新型生物抗菌剂的应用价值。 环境耐受性是噬菌体生产应用过程中的重要考量因素之一。本研究中,噬菌体PSF26在pH 5.0~8.0范围内效价高于108pfu/mL,相比于其他噬菌体,如烈性青枯菌噬菌体(pH 7.0效价最高为106pfu/mL)[25],噬菌体PSF26具有很好的酸碱耐受性。在0~40 ℃范围内噬菌体PSF26效价可达109pfu/mL,80 ℃时,噬菌体PSF26才完全失活,相比于鸡粪中分离的噬菌体PNJ1809-36 (60 ℃即完全失活),噬菌体PSF26也表现了良好的温度耐受性[26]。 据统计,我国珠江流域排放阿莫西林浓度已高达3 384 ng/L,生态环境尤其是养殖业抗生素污染触目惊心,亟需严控[27-28]。噬菌体作为一类能特异性识别并感染细菌的病毒,专一性地以细菌为宿主进行复制和繁殖,一般对人和动物无害,成为对抗耐药性细菌感染的“天然武器”[29-30]。本研究系统分析了噬菌体PSF26的生物学特性,发现其具有爆发期短、裂解量高和耐受性强等特点,证实了其在防控耐药性志贺氏菌中具有良好的应用潜能,为今后鸡尾酒治疗策略制定、饲用抗生素替代和动物源性耐药志贺氏菌防控提供参考。2 结果与分析
2.1 志贺氏菌的分离鉴定

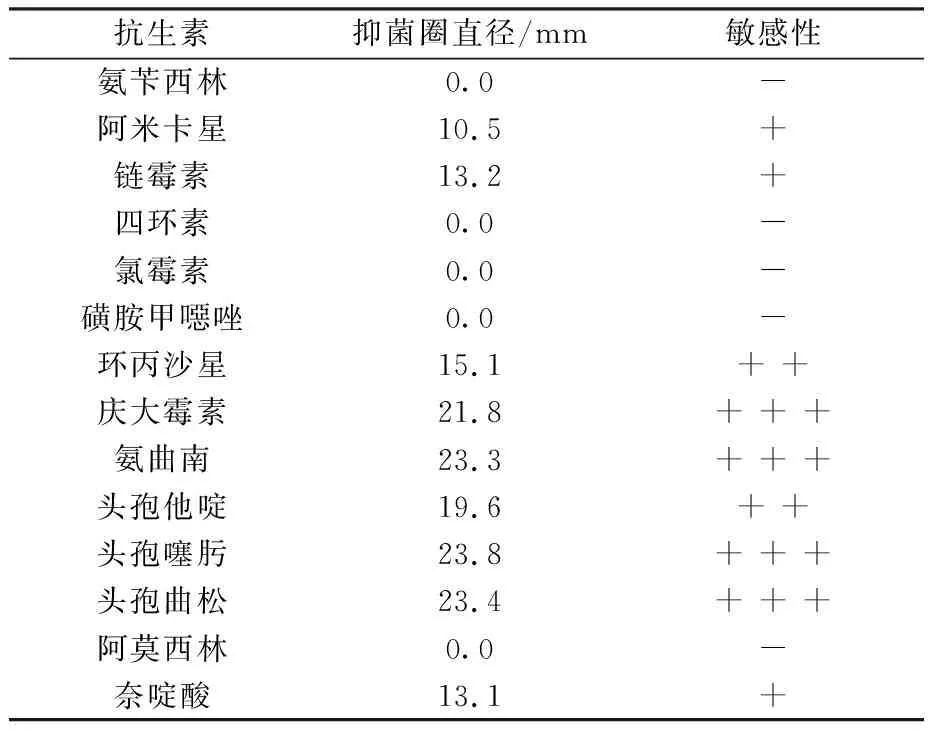
2.2 志贺氏菌耐药性测定
2.3 噬菌体的分离纯化
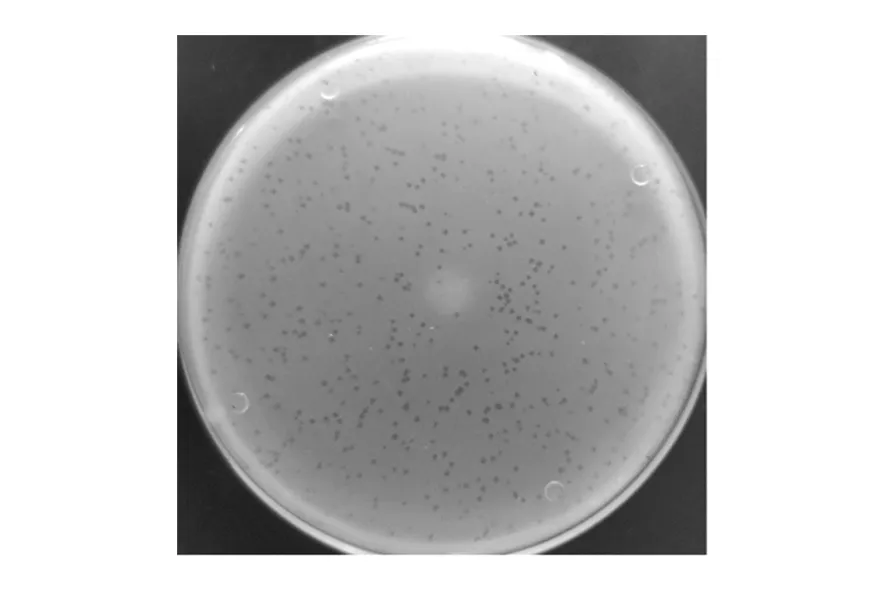
2.4 噬菌体的形态特征

2.5 噬菌体的生物学特性分析



3 讨 论
猜你喜欢
心理学探新(2022年1期)2022-06-07
昆明医科大学学报(2022年2期)2022-03-29
中国动物传染病学报(2021年3期)2021-07-21
猪业科学(2018年5期)2018-07-17
寻根(2016年3期)2016-06-28
广东海洋大学学报(2015年3期)2015-12-22
中国当代医药(2015年8期)2015-03-01
沈阳医学院学报(2014年4期)2014-12-27
中国信息化周报(2014年25期)2014-08-07
中成药(2014年9期)2014-02-28
