
黄、东海秋季群落净生产力的分布特征及其影响因素❋
2023-11-25 12:53韩宗奕郑文静张桂玲简慧敏
中国海洋大学学报(自然科学版) 2023年12期
韩宗奕, 秦 川, 郑文静, 张桂玲, 简慧敏❋❋
(1. 中国海洋大学深海圈层与地球系统前沿科学中心和海洋化学理论与工程技术教育部重点实验室, 山东 青岛 266100; 2. 青岛海洋科学与技术试点国家实验室 海洋生态与环境科学功能实验室, 山东 青岛 266237)
群落净生产力(Net Community Production,NCP)是总初级生产力(Gross Primary Production,GPP)和群落呼吸(Community Respiration,CR)的差值,反映了表层海水可以向深层海洋输送的最大有机质量,是衡量生物活动对上层海洋碳循环影响的重要指标[1-2]。由于群落净产生力在不同时间尺度上都表现出高度动态变化,连续和准确测定群落净生产力是非常必要的,对认识全球碳循环和海洋碳汇有重要作用[3]。在透光层中,NCP通过光合作用与呼吸作用的耦合与生物活动紧密相连,O2作为海洋初级生产的主要产物,在光合作用中与碳存在化学计量关系,因此可以利用O2的变化定量估算NCP[4-5]。但O2在海洋中会同时受到物理和生物过程的影响,为了准确定量NCP,必须消除物理过程的干扰。由于惰性气体Ar与O2有相似的溶解特性,通过测定O2/Ar比值将二者归一化可去除物理过程对于O2质量平衡的影响,从而实现高分辨率的NCP估算[6]。目前O2/Ar比值法已经成为估算海洋混合层群落净生产力的重要方法,广泛地应用到南大洋、太平洋、亚极地海域等[7-9],获得了这些代表性大洋中NCP高分辨率时空分布特征及其影响因素,但是目前对陆架边缘海的观测仍较匮乏。近岸或陆架边缘海具有富营养和高生物生产的特征,其生产力约占全球海洋总初级生产力的10%[10]、群落净生产力的29%[11],每年吸收大约0.2~0.4 Pg C[12],是重要的海洋碳汇;但是其碳通量具有高度时空变异性,碳源汇格局及生物泵效率要比大洋更为复杂。因此研究代表性陆架边缘海群落净生产力的时空变化及其影响因素对准确评估全球海洋碳汇强度有重要意义。
东、黄海是西北太平洋的陆架边缘海[13],具有复杂多变的海洋环境[14]。受到陆源输入、高温高盐的黑潮水入侵以及不同水团相互作用等高度动态的物理过程影响,东、黄海初级生产力具有较大的时空差异[15-17]。与广泛开展的高时空分辨的碳吸收研究相比,目前关于东、黄海碳输出的研究还相对不足,鲜有群落净生产力的数据报道,仅有韩国科学家报道了南黄海及东海部分海域的群落净生产力分布[15]。因此,本文基于连续走航观测获得的高分辨率O2/Ar数据,估算了黄、东海混合层NCP并结合其他辅助水文和生化参数,分析探讨了NCP的分布特征及影响因素。
1 材料与方法
1.1 调查区域及航行轨迹
本文于2015年10月19日至11月2日搭乘“东方红2”考察船GEOTRACES GP06-CN航次对黄、东海陆架(26°N—36°N,121°E—128°E)进行了为期14天的调查,调查站位和航迹见图1。本文根据研究区域的主要水文特征,将其分为A—D四个区域(见图1):南黄海中部、长江口(S<30)、浙闽沿岸(深度<50 m)和东海陆架(S>30,50 m<深度<200 m)。利用膜进样质谱仪(MIMS)走航连续测定了表层海水中(-5 m)的O2、Ar等溶解气体,同时走航测定了表层海水的温度、盐度、溶解氧以及叶绿素等基本参数,以认识东海、黄海表层海水的氧氩比值和群落净生产力的分布规律。
1.2 高分辨率走航观测系统
参考Guéguen和Tortell[18]的方法,我们搭建了用于测定溶解气体的连续走航观测系统[9]。利用考察船走航水系统抽取表层海水(约水下5 m)并分为三路:第一路用于海水中溶解气体的测定,海水首先流入缓冲池以消除气泡,随后利用蠕动泵(Masterflex,USA)以恒定流速(约220 mL·min-1)抽取水样。为了减小温度效应和水蒸气压力变化造成的O2/Ar波动,海水先流经置于恒温水浴中的不锈钢管(6 m长)进行恒温(温度设置为低于表层海水温度约2 ℃),然后进入膜进样质谱仪(HPR-40,英国Hiden公司)的进样池。在质谱仪的高真空作用下,海水中的溶解气体分子会扩散通过半透膜进入到仪器内部,经离子化后,根据气体分子不同的质核比(m/z)完成不同气体(O2、Ar和CO2)的浓度信号值测定。第二路海水流入一个安装有多参数水质分析仪(RBR Maestro,加拿大)的水箱进行温度、盐度、溶解氧(DO)、叶绿素a(Chla)等参数的测定。第三路则用于排掉过量海水并粗略调节其他两路的流量。走航过程中的经纬度、船速和风速等参数由船载的自动气象站提供。不同仪器获取的走航数据根据北京时间一一对应,并最终处理为5 min间隔的数据集。
1.3 气体信号值的校正方法
MIMS获取的O2和Ar的信号比值(O2/Ar)利用与大气平衡后的海水标准样进行校正[18]。取1 L过滤(0.22 μm)后的海水装入聚碳酸酯瓶中,置于恒温水槽中 (温度设置为与表层海水温度相同),用空气泵连续鼓泡24 h,使其与大气达到平衡,然后利用MIMS测定其O2/Ar。测定前先用200 mL海水清洗管路,取进样2~3 min后信号达到平衡时的数据平均值,记为([O2]/[Ar])eq。O2/Ar的校正每隔6~8 h进行一次,且同时进行多组标准样的制备以满足校正需求。
每隔6 h从走航观测系统中采集离散水样进行DO和Chla校正。其中DO样品采用Winkler滴定法测定,而Chla水样(2 L)则用0.22 μm滤膜过滤后,将滤膜冷冻保存于-20 ℃,带回实验室后,依照《海洋调查规范》采用荧光法测定。
1.4 基于Δ(O2/Ar)的群落净生产力计算
由于O2和Ar具有相似的物理特性,Δ(O2/Ar)作为生物过饱和氧的指标,可消除物理过程(如海气交换、温度变化、压力变化、气泡注入等)对O2的影响[6],其计算公式如下:
式中:[O2]/[Ar]是MIMS走航测定的海水中溶解O2和Ar的信号比值;([O2]/[Ar])eq则是空气鼓泡得到的与大气平衡的标准海水样中O2与Ar的信号比值;Δ(O2/Ar)是测量的O2和Ar的信号比值偏离平衡的百分比。在忽略垂直混合的稳态条件下,NCP可视为Δ(O2/Ar)和海气交换速率的函数[4]:
NCP(mmol·m-2·d-1)≈kO2·[O2]eq·
Δ(O2/Ar)·rC∶O2·ρ。
式中:kO2是加权后的O2海气交换速率常数(m·d-1);[O2]eq是溶解O2在混合层中的饱和浓度[19](μmol·kg-1);ρ是海水密度(kg·L-1);是光合作用中C和O2的摩尔光合商,通常为1∶1.4[20]。我们使用了欧洲中期天气预报中心(ECMWF)风速再分析数据产品,分辨率为0.25°×0.25°(https://www.ecmwf.int/en/forecasts),利用Wanninkhof[21]报道的方法将风速进行参数化得到气体交换速率k,然后结合Teeter等[22]报道的方法对调查前30天的k值进行加权计算,最终得到kO2。在近岸复杂多变的环境条件下,基于O2/Ar估算的NCP并不反映该区域实际的群落净生产的日产量,而是代表O2在混合层中停留时间内积累的生物氧通量[8,23]。
2 结果与讨论
2.1 水文和生化参数的分布
2015年10月黄、东海表层的温度和盐度分布见图2。表层海水的温度范围为19.37 ℃~26.73 ℃,平均值为(23.38±1.89) ℃,从北向南温度逐渐升高,黄海表层海水的温度(19.37 ~23.21) ℃明显低于东海(21.33~26.73) ℃。图1(b)显示了黄海沿岸流将温度和盐度较低的沿岸水向东南方向输送[24-25],除了受长江冲淡水影响的长江口区域,东海表层海水的盐度(33.29±0.79)明显高于黄海(31.38±0.56),受长江冲淡水(S<30)向东南方向扩散与东海沿岸水的共同影响[26-27],浙闽沿岸的盐度明显低于东海外陆架。此外,受台湾暖流和黑潮表层水的影响,东海的西南和东南海域分别出现了高温和高温高盐现象[28]。叶绿素(Chla)的整体变化范围在(0.15~8.76) μg·L-1之间,平均值为(0.92±1.07) μg·L-1,在长江口出现极大值(最高可达8.76 μg·L-1),浙闽沿岸叶绿素浓度高于其它调查区域。黄海DO浓度((6.47±0.28) mg·L-1)整体略高于东海((6.31±0.36) mg·L-1),但低于长江口和浙闽沿岸。上述海区出现了DO浓度高值,其中最大值(8.15 mg·L-1)出现在长江口附近,本文结果与石晓勇等[29]报道的2006年11月长江口邻近海域DO结果(5.32~8.47 mg·L-1)基本一致。

图2 2015年10月黄、东海表层温度(℃)(a)、盐度(b)、叶绿素a(μg·L-1)(c)和溶解氧(mg·L-1)(d)的水平分布

图3 2015年10月黄东海表层氧氩比(%)(a)和NCP (mmol· m-2·d-1)(b)的分布
2.2 Δ(O2/Ar)与NCP的分布
2015年10月黄、东海的Δ(O2/Ar)范围为(-30.21%~44.38%),平均值为(0.32±8.29)%。其中,低值主要出现在区域A和区域B交界的长江口北侧,其溶解氧浓度也较低,Δ(O2/Ar)处于不饱和状态。而在长江口(11.75±13.75)%和浙闽沿岸(5.42±5.21)%的Δ(O2/Ar)明显高于南黄海和东海陆架,这是由于长江冲淡水携带了大量营养盐输入,导致生物量较高,因此叶绿素和Δ(O2/Ar)均较高。NCP与Δ(O2/Ar)的分布趋势大体一致,黄海群落净生产力在(-120.41~41.65) mmol· m-2·d-1之间变化,其平均值为(-9.24±23.15) mmol· m-2·d-1,与该航次其他三个研究区域相比处于最低值(见表1),但是去除DO<6.5 mg·L-1的区域后黄海的平均NCP约为(4.37±9.24) mmol· m-2·d-1。在长江口(B区域)和浙闽沿岸(C区域)群落净生产均为正值,其中长江口区的平均NCP((24.49±35.74) mmol· m-2·d-1)是浙闽沿岸((13.56±13.42) mmol· m-2·d-1)的2倍。而东海陆架区(区域D)受陆源输入和沿岸流的影响较小,NCP平均值(-4.04±18.68) mmol· m-2·d-1)相较于近岸显著降低,为净异养状态,这与Chen[26]和叶文琪等[30]分别于1998年和2011年秋季在东海进行浮游生物群落呼吸研究发现其呈异养状态的结果相一致。
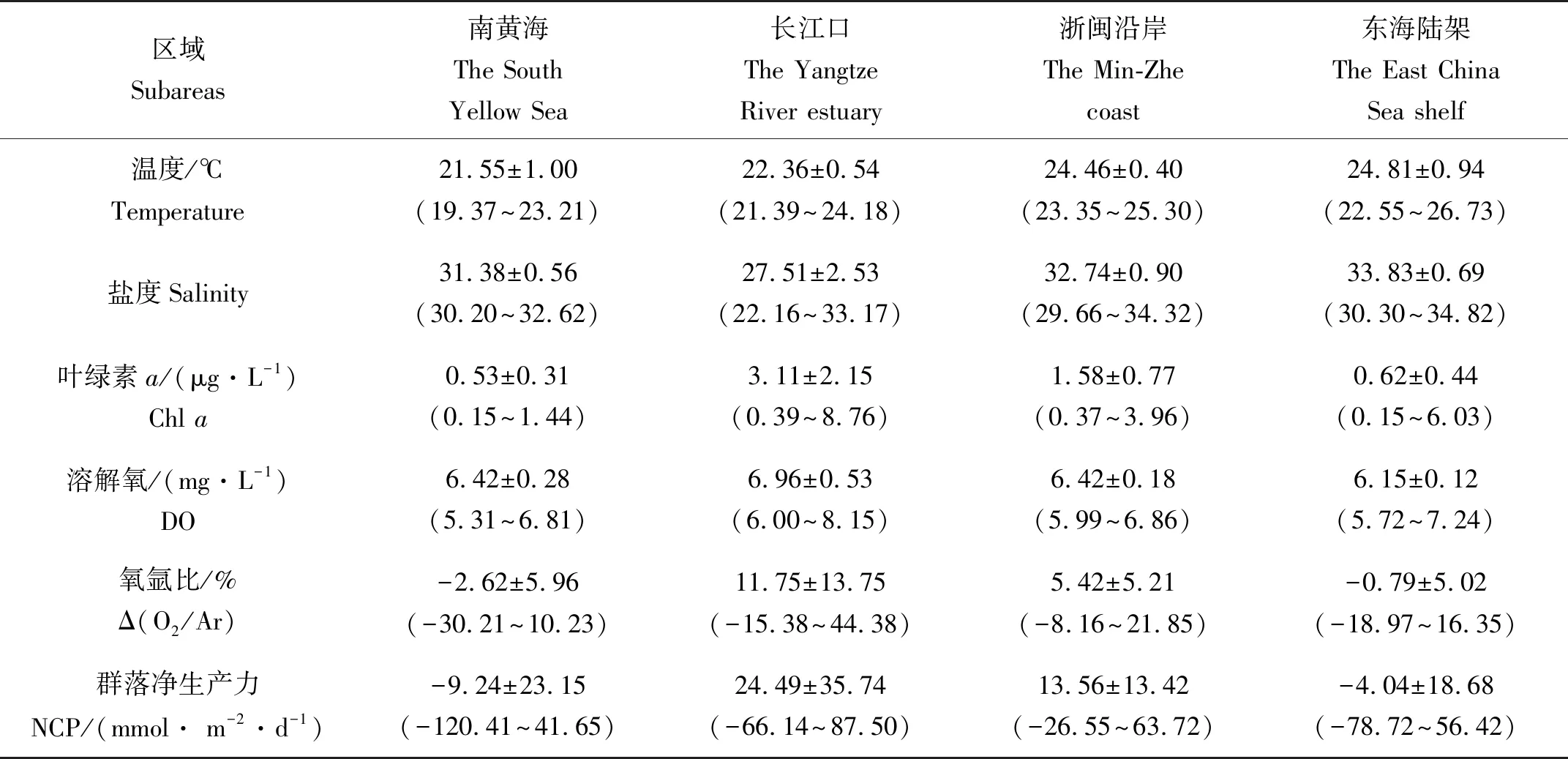
表1 2015年10月黄东海航次不同区域的Δ(O2/Ar)、NCP和相关参数结果汇总
受到季风、陆源输入、沉降和蒸发、黑潮及多种水团的物理过程影响[31],黄、东海生态系统生产力有着复杂的时空变异性,然而目前对黄东海群落净生产力的研究较少。Jang等[32]利用13C同位素示踪技术测定碳吸收速率,估算了夏季黄海中部的总初级生产力平均值为(291±165) mmol· m-2·d-1。Lee等[15]基于O2/Ar法估算得到,春季受黄海冷水团控制的济州岛西北部海区NCP为(35±13.3) mmol· m-2·d-1,然而该航次在黄海南部与长江口交汇的区域也观测到了净异养的结果。目前,陆源输入有机质被发现是沿岸陆架区域系统出现异养状态的主要原因[33],秋季高有机质含量的沿岸水和陆源输入对黄海影响较大[34-35]。在黄海南部区域出现DO低值区(DO<6.5 mg·L-1)[36],与谢琳萍[35]在南黄海发现的TOC高值区高度重合,高有机质含量会导致呼吸作用的加强。同时在叶文琪的调查中,南黄海秋季的浮游生物群落呼吸速率(Plankton Community Respiration,PCR)平均值为(237.17±221.42) mg·m-2·d-1,远高于东海陆架区[30]。所以研究区域呼吸作用较强,初级生产力通过光合作用产生的O2低于呼吸作用消耗的O2可能是本航次南黄海陆架区出现净异养的主要原因。张玉荣等[37]利用14C方法测得东海秋季初级生产力为490.9 mmol· m-2·d-1,这里可以看作是总初级生产力(GPP),该航次利用氧氩比值方法计算得到的东海NCP约占GPP的6.9%,与之前研究报道的NCP/GPP变化区间(5%~40%)一致[38]。
2.3 影响黄、东海NCP分布的因素
黄、东海NCP分布呈现明显的区域分布特征,受水团性质及水团混合的影响。图4为该航次Chla和NCP的T-S点聚图,不同区域表现出不同的生物量和群落净生产状态,其基本参数汇总于表1。南黄海(区域A)主要受到黄海沿岸水和黄海冷水团的影响,秋季在东北季风的驱使下该区域平均温度明显低于其他三个区域,22 ℃以下的低温水普遍出现在黄海。长江口北侧的DO低值区对应谢琳萍[35]观测到的TOC含量高值区,表层和底层TOC分别可高达6.93和13.8 mg·L-1,由于长江冲淡水向南移动,该区域水体混合加强,同时该区域还受到南下的苏北沿岸流的影响,导致水体TOC含量较高,呼吸作用加强,出现净异养状态。而在黄海中部,黄海冷水团一定程度上阻隔了底层的营养盐向上补充[34],限制了上层水体营养盐的来源[39],浮游植物生物活动减弱导致该区域的群落净生产力较低。长江口(区域B)的平均盐度与其他区域相比明显偏低(27.51±2.53),这是长江冲淡水(26<盐度<30)影响的结果[28]。秋季长江冲淡水会携带高浓度的DIN和硅酸盐等营养盐入海[40],并且沿浙闽沿岸的狭带向南扩展[41]。该航次从北向南经过长江口时对应图4(a)中温度>22 ℃的表层海水,其Chla浓度处于较高水平。高浓度的营养盐会促进浮游植物的生长从而带来较高的生产力,距离长江口最近的P2站观测到了本航次DIN(27.13 μmol·L-1)和硅酸盐(18.48 μmol·L-1)的最高浓度(见图6(a)),同时对应着NCP的高值。然而在同一区域的P3站NCP却表现为负值,尽管P3站无机氮含量仍较高,硅酸盐含量却大幅降低。叶文琪等[30]发现秋季黄东海PCR与硅酸盐呈极显著的负相关,由于群落净生产力是总初级生产力与群落呼吸的差值,因此可推测P3站浮游生物呼吸速率较强导致了净异养状态的出现。但是总的来说,因为受到陆源输入的影响,长江口区域Chla平均浓度达到(3.11±2.15) μg·L-1,是浙闽沿岸的2倍、南黄海和东海陆架区的5倍,NCP也是该航次4个区域中的最高水平((24.49±35.74) mmol· m-2·d-1)。区域C位于浙闽沿岸,主要受到东海沿岸流和向东南方向延伸的长江冲淡水羽流的影响[26,28]。浙闽沿岸纬度较低,温盐较高,其Chla浓度仅次于长江口,对应的群落净生产相较于长江口下降了50%,整体呈现净自养状态。东海陆架在温度>24 ℃的海区主要受到高温高盐的黑潮表层水和台湾暖流的影响[28],具有最高的温度和盐度(见表2),同时继承了黑潮水寡营养盐的特点,表现出低浮游植物生物量的特点,Chla平均浓度((0.62±0.44) μg·L-1)基本与南黄海持平。而在东海陆架北部温度和盐度有所减弱(t<24 ℃),与南黄海相当,这是部分长江冲淡水向东北扩展与苏北沿岸流朝东南方向扩散混合的结果[40]。同时长江口东北方向存在一个高营养区[42],在图6(a)中可以观察到东海北部陆架的QT14,QT15和QT16三个站位的DIN浓度明显高于其他邻近站位,这可能是受到河流输入影响造成的无机氮相对过剩,因此在Chla浓度有所增加的情况下NCP并未相应提高,这与王保栋在东海北部和南黄海的调查中发现浮游植物的生长不受N限制相符[40],可能存在其他因素抑制了浮游植物的初级生产。叶文琪等[30]研究发现秋季长江口表层的PCR相对较低,同时王保栋[40]和郝锵[42]的研究发现长江冲淡水在扩散过程中伴随着悬浮泥沙的沉降,导致水体浑浊,光可利用率降低,从而限制了浮游植物的生长[42],因此光照可能是长江口附近群落初级生产力的限制因素。

(图中的四色框线代表A—D四个区域的温度和盐度范围。The four coloured box lines in the diagram represent the temperature and salinity ranges in the four subareas A—D.)
为了认识不同区域内NCP的分布特征和主要贡献因素,利用origin分析了四个区域NCP与环境因子(温度、盐度、DO、Chla和营养盐)之间的相关性关系(见图5)。在南黄海,NCP与温度呈显著的负相关(r=-0.43,p<0.01),NCP与DO呈明显的正相关,在长江口及浙闽沿岸都观察到了NCP与Chla呈显著正相关(r=0.79,p<0.01;r=0.85,p<0.01)。而在区域D观察到NCP与温度呈一定的正相关,表明温度降低可能减弱生物活动,对于NCP有较大的影响。长江口及浙闽沿岸呈现群落净生产力较高的特征,主要是由于高营养盐支持着浮游植物的高生物量。从图6中可以看出区域B和C的无机氮、磷酸盐和硅酸盐含量整体上高于区域D,但是高营养盐含量并不一定带来高的NCP。在图6(a)中我们可以观察到该航次NCP与DIN没有统计学意义上的关联,DIN浓度的高值出现在P2(27.13 μmol·L-1),QT4(13.37 μmol·L-1)和P3(9.06 μmol·L-1)站,然而除P2站对应着高的群落净生产力,QT4和P3站的NCP都很低甚至在P3出现了较大的负值。这可能是由于长江冲淡水携带大量无机氮进入东海陆架,导致浮游植物生长所需的营养盐远超适宜其生长的浓度,DIN不再是浮游植物生长的限制因素。王保栋[40]发现秋季长江口和东海近岸浮游植物存在磷限制,在图6(b)中观察到长江口附近(QT14、P3站)和浙闽沿岸(QT1,QT2和QT3站)部分站位的磷酸盐浓度极低(<0.15 μmol·L-1),这些站位都受到了北上的寡营养盐的台湾暖流和黑潮表层水的影响,NCP与磷酸盐总体呈现显著的负相关(r=-0.615,p<0.05)。虽然王保栋[40]发现硅酸盐不是该区域浮游植物的限制性因素,而本研究发现NCP与硅酸盐有极显著的正相关(r=0.708,p<0.05)(见图6(c)),这与叶文琪等[30]在黄东海秋季观察到PCR与硅酸盐呈负相关相一致,同时郝锵[42]指出长江口附近较高的硅酸盐浓度(>0.5 μmol·L-1)有助于硅藻占优的浮游植物群落保持较高的生物量水平。综上所述,营养盐会影响东黄海NCP的分布,但是并不会成为控制NCP分布的限制因素。

图5 2015年10月A—D(a—d)四个区域的温度,盐度,叶绿素a,溶解氧与群落净生产力的相关性分析
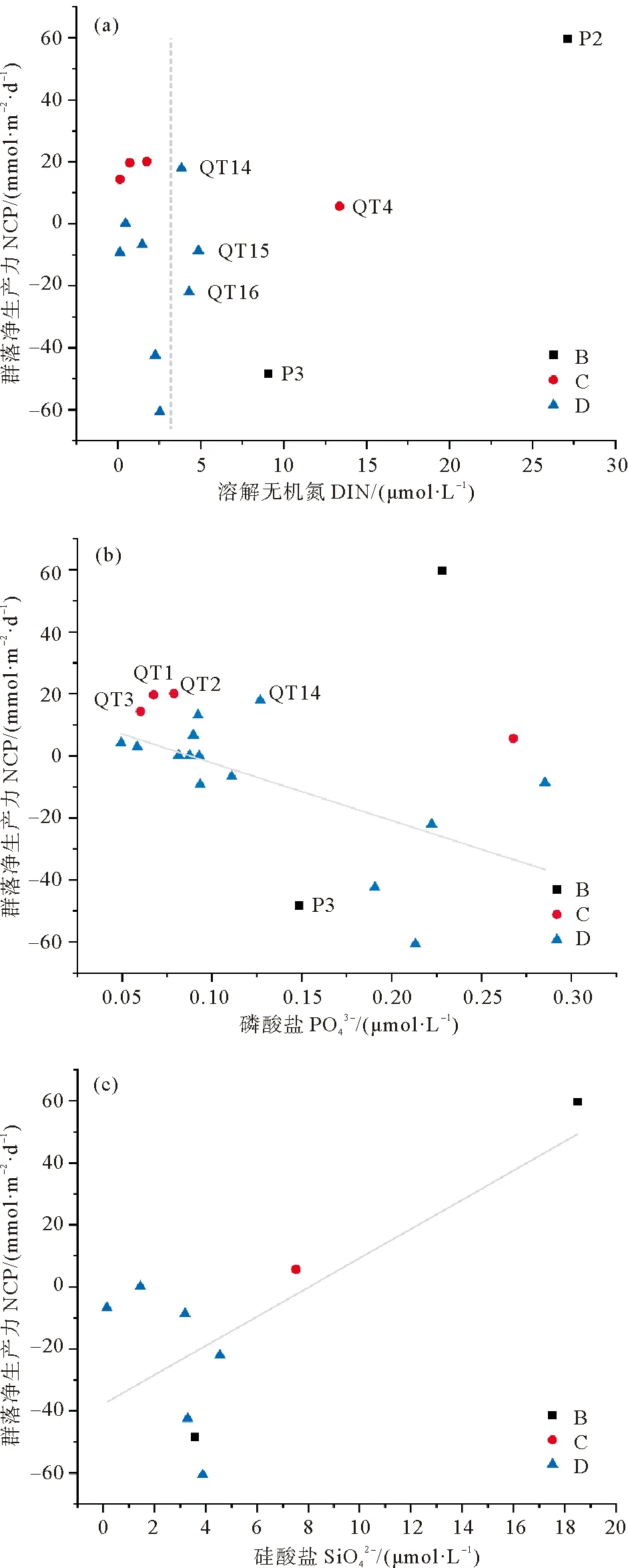
图6 2015年10月黄东海航次NCP与溶解无机氮(a)、磷酸盐(b)和硅酸盐(c)的相关性分析
除此之外,混合层深度(MLD,Mixed Layer Depth)可以作为混合层光可利用程度的重要指标,较浅的混合层深度意味着更充足的光照,为了归一化由MLD引起的NCP变化,定义NCP与MLD的比值为单位体积水体的群落净生产力(NCPvol)[43-44]。选取从北向南的区域B—D的部分站位进行分析,发现各站的MLD在5~45 m之间,且MLD与NCPvol表现为一定的负相关(见图7),在MLD较浅的站位对应着混合层中单位体积更高的NCP。同理,在QT16和QT15站位MLD到达40 m左右时,单位体积的群落净生产大幅下降。傅明珠等[39]研究也发现真光层与混合层的相对深度对群落净生产有影响,混合层深度大于真光层深度时浮游植物白天也会处于无光区,呼吸作用大于光合作用从而减弱NCP。另外,王保栋[40]和郝锵[42]研究发现,长江口及东海沿岸大量悬浮的泥沙会使透光率下降导致真光层变浅。综上所述,光照是秋季影响黄、东海NCP的重要限制因素,也是富营养的近岸海区NCP呈现低值的主要原因。

图7 2015年10月黄东海航次单位体积水体的群落净生产力与混合层深度的相关性分析
3 结语
该研究首次基于O2/Ar比值法获得了中国陆架边缘海—东、黄海秋季生物氧饱和度((O2/Ar)和群落净生产力(NCP)的高分辨率分布特征。在受到长江冲淡水强烈影响的长江口NCP较高((24.49±35.74) mmol· m-2·d-1),是浙闽沿岸((10.85±12.17) mmol· m-2·d-1)的2倍多。水温较低的南黄海和受到黑潮影响的东海陆架区NCP分别为(-9.24±23.15)和(-4.04±18.68) mmol· m-2·d-1,均呈现净异养状态。秋季黄海和东海的群落净生产分布呈现高度的空间异质性,主要受到陆源输入和黑潮水、台湾暖流水团混合的影响。同时,营养盐和光照也是影响该区域NCP分布的重要因素。
致谢:感谢中国海洋大学海洋化学理论与工程技术教育部重点实验室的同学们在采样中提供帮助!
猜你喜欢
海洋通报(2022年2期)2022-06-30
石油地球物理勘探(2022年3期)2022-06-11
水利技术监督(2017年6期)2017-12-19
海洋渔业(2016年6期)2016-04-16
海洋渔业(2016年6期)2016-04-16
广东海洋大学学报(2015年3期)2015-12-22
应用海洋学学报(2015年3期)2015-11-22
应用海洋学学报(2014年1期)2014-11-22
华东师范大学学报(自然科学版)(2014年3期)2014-03-11
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
