
大熊猫从幼年到亚成体粪便真菌多样性及功能分析
2023-11-25 08:57刘学锋郝菲儿崔多英郑常明杨明海张成林
野生动物学报 2023年4期
刘 燕,刘学锋,胡 昕,郝菲儿,崔多英,刘 赫,滑 荣,卢 岩,郑常明,杨明海,张成林
(北京动物园管理处,圈养野生动物技术北京市重点实验室,北京,100044)
大熊猫(Ailuropoda melanoleuca)是我国特有濒危珍稀物种。根据全国第4 次大熊猫调查结果,野生大熊猫分布在秦岭、岷山、邓峡山、大相岭、小相岭和凉山6 大山系,共有1 864 只,圈养大熊猫种群数量达到375只[1]。有研究表明,肠道菌群对大熊猫机体健康有重要影响[2-3],包括参与营养物代谢、能量获取、免疫系统发育及炎症性肠病等,同时肠道菌群多样性也受饮食结构和生活地域的影响[4]。大熊猫本身不能分泌纤维素酶,消化纤维素主要由肠道微生物来完成,肠道微生物的改变可引起消化功能紊乱甚至疾病。与细菌相比,肠道中的真菌所占比例不高,但可影响大熊猫机体疾病的发生及对纤维素的消化利用。由于大量真菌不可培养,通过分离培养方法获得的真菌在种类、数量和功能上无法反映自然状态下微生物群落的真实情况,而宏基因组测序技术具有操作简单、所需样本量少、检测快、高通量以及可以检测出不可培养的微生物的优点,因此可应用到大熊猫肠道真菌的研究中。
针对大熊猫肠道真菌的研究主要集中在成年和老年[5-6],对幼年向亚成体过渡过程中肠道真菌的研究还鲜见报道。大熊猫从幼年向亚成体过渡阶段伴随着食物的转换,对成年后的健康影响较大。鉴于此,对北京动物园3 只年龄相仿的大熊猫开展研究,分别于不同月龄采集新鲜粪便,采用高通量测序技术对粪便内真菌多样性和群落功能进行分析,以期为幼年至亚成体大熊猫的肠道疾病诊断提供基础理论数据。
1 样本采集
2013 年8 月出生的3 只大熊猫呼名分别为“萌大”“萌二”和“福禄”,出生后一直饲养于成都,于2014 年10 月转到北京动物园饲养,其中,“萌大”和“萌二”是一对双胞胎。分别于14、16、21、26、31、36 月龄采集3 只大熊猫的粪便样本,每只每次采集3份,即3个重复,共计54份。将“萌大”“萌二”和“福禄”分别编号为D、E 和F。粪便采集时,佩戴一次性无菌手套,使用无菌棉签从新鲜粪便内部采集10 g左右,立即用液氮保存,回到实验室放入冰箱-80 ℃保存待检。
2 试验方法
2.1 基因组DNA抽提
用E.Z.N.A.®Stool DNA Kit 试剂盒(广州捷倍斯生物科技有限公司)提取大熊猫粪便中的基因组DNA,具体操作步骤按照说明书进行。用1%琼脂糖凝胶电泳检测抽提的基因组DNA,抽提合格后置于冰箱-20 ℃保存备用。
2.2 18S rRNA基因扩增及测序
以大熊猫基因组DNA 为模板,利用正向引物817F(5'-TTAGCATGGAATAATRRAATAGGA-3')和反向引物1196R(5'-TCTGGACCTGGTGAGTTTCC-3')扩增18S rRNA 基因。PCR 扩增反应体系:5×Fast Pfu Buffer 4.0 µL,2.5 mmol/L dNTPs 2.0 µL,上游引物(5 µmol/L)0.8 µL,下游引物(5 µmol/L)0.8 µL,Fast Pfu 聚合酶(5 U/µL)0.4 µL,模板DNA10.0 ng,然后补ddH2O至20.0 µL。PCR扩增程序:95 ℃预变性3 min;95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸45 s,共27 个循环;72 ℃延伸10 min,10 ℃保存。将每只每次采集的3 个重复样本的PCR 产物混合后,取3 µL 用2%琼脂糖凝胶电泳检测PCR 产物,并将合格的产物使用AxyPrep DNA 凝胶回收试剂盒(AXYGEN公司)切胶回收纯化PCR产物。
2.3 高通量测序
将扩增产物送上海美吉生物医药科技有限公司,利用Illumina MiSeq测序平台进行高通量商业化测序。
2.4 数据分析
经MiSeq测序得到的PE reads,根据overlap关系拼接,然后对序列质量过滤和质控。应用Mothur v1.30.1软件进行Coverage、Simpson和Ace 3种常用指数的测定。基于OTU 聚类分析结果,进行多样性指数分析和测序深度检测,并运用统计学t检验检测两组间的指数值是否具有显著性差异。应用Usearch v7.0 软件在相似性97%的条件下,对OTU 代表序列进行聚类分析和物种分类学分析。根据得到的真菌群落丰度数据,利用LEfSe 多级物种差异判别方法进行分析。使用非参数因子克鲁斯卡尔-沃利斯秩和检验检测显著丰度差异特征,得到与丰度有显著性差异的类群。通过真菌功能注释数据库(http://www.funguild.org),利用FUNGuild 通过微生态guild对真菌群落进行分类分析。
3 结果
3.1 样本数据统计
54 份样本共获得1 680 845 条有效序列,每个样本平均含有31 126 条序列,每条序列平均长度为289.4 bp。序列共分属于23 门、42 纲、76 目、101 科、108属、170种和2 407个OTU。
3.2 Alpha多样性分析
3 只大熊猫54 份样本的覆盖率(coverage)均达到99.5%以上。根据反应真菌群落多样性的Simpson 指数可知(图1A),14~26月龄真菌群落多样性差异不显著,而26~36 月龄有所下降。根据反应真菌群落丰富度的Ace 指数可知(图1B),14~16 月龄真菌群落丰富度有所下降,16~26 月龄逐渐升高,但26~36 月龄又逐渐下降。总体看,26 月龄大熊猫粪便真菌的多样性和丰富度均达到最高值。3 只大熊猫个体间比较,真菌群落多样性和群落丰富度均没有差异(图2)。

图1 不同月龄大熊猫间Simpson指数(A)和Ace指数(B)Fig.1 Simpson index(A)and Ace index(B)of giant pandas at different month of age
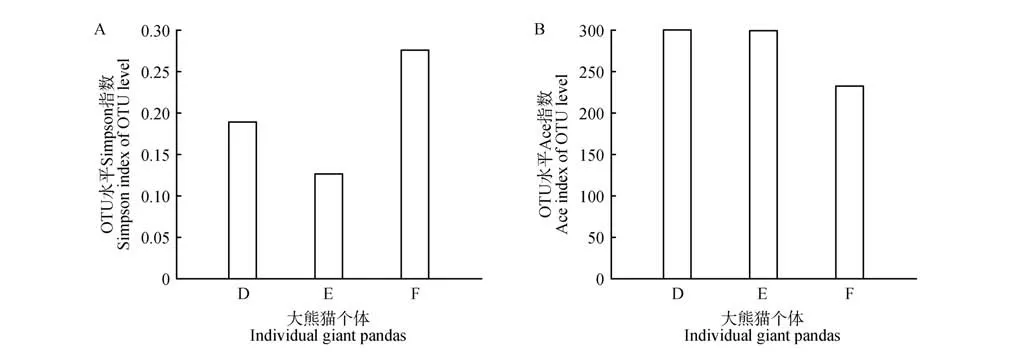
图2 大熊猫个体间的Simpson指数(A)和Ace指数(B)Fig.2 Simpson index(A)and Ace index(B)of giant pandas
3.3 Venn图分析
3 只大熊猫在属水平上共有和特有的真菌情况见图3A。3只大熊猫共同拥有66个菌属,D特有4个菌属,E 特有19 个菌属,F 特有8 个菌属。不同月龄间,共同拥有14 个菌属,14 月龄特有6 个菌属,16 月龄无特有菌属,21月龄特有5个菌属,26月龄特有4个菌属,31 月龄特有1 个菌属,36 月龄特有40 个菌属(图3B)。

图3 大熊猫个体(A)和不同月龄(B)间的真菌Venn图Fig.3 Venn graph of fungi of three giant pandas individuals(A)and at different month of age(B)
3.4 菌群组成分析
在门水平上,3只大熊猫不同月龄粪便样本中的真菌菌门主要为子囊菌门(Ascomycota)、unclassified_k__Fungi、unclassified_k__Metazoa__Animalia_、unclassified_d__Eukaryota、担子菌门(Basidiomycota)和Phragmoplastophyta 等(图4A)。在目水平上,3 只大熊猫不同月龄粪便样本中的真菌菌目主要为假球壳目(Pleosporales)、unclassified_p__Ascomycota、unclassified_c__Dothideomycetes 和 煤炱目(Capnodiales)等(图4B)。
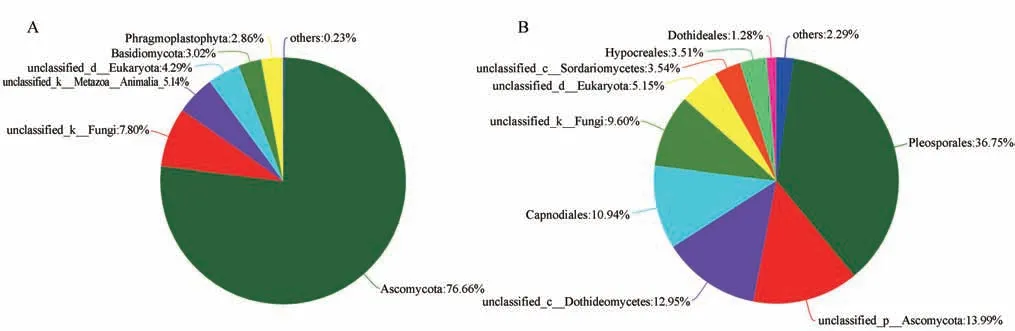
图4 大熊猫粪便样本中门(A)和目(B)水平的真菌组成Fig.4 Fecal fungi of giant pandas at the level of phylum(A)and order(B)
3.5 真菌群落差异分析
根据偏最小二乘法判别分析(PLS-DA),在属水平上对不同月龄大熊猫粪便样本真菌进行比较,发现31 月龄的真菌组成与其他月龄开始出现分离,至36月龄均表现出明显不同(图5)。在门水平上,36月龄与其他月龄比较,主要差别体现在担子菌门、Phragmoplastophyta、unclassified_d__Eukaryota、unclassified_k__Metazoa__Animalia_ 和 unclassified_k__Fungi 等(图6)。36 月龄大熊猫的担子菌门比例高于其他月龄,而其余菌门的比例低于其他月龄。

图6 36月龄与其他月龄间门到属水平的真菌差异Fig.6 Fungi difference analysis at the level of phylum to genus between 36 months of age and the other months of age
3.6 群落功能分析
FUNGuild 功能预测结果表明,从不同月龄大熊猫粪便中得到的真菌从功能上主要属于未知、植物病原菌(plant pathogen)、未定义腐生菌(undefined saprotroph)和地衣-木质腐生菌(lichenized-wood saprotroph)等10 类。除未知功能的部分,有6 类是通过降解死亡的宿主细胞来获取营养的腐生营养型,2类是通过损害宿主细胞而获取营养的病理营养型,没有通过与宿主细胞交换资源来获取营养的共生营养型,说明已知真菌主要是通过腐生方式与大熊猫互惠互利。不同月龄间比较,发现36 月龄与其他月龄的差异较大,未知菌占比较少,而未定义腐生菌占24.3%、内生-植物病原菌占20.4%,均高于其他月龄(图7)。
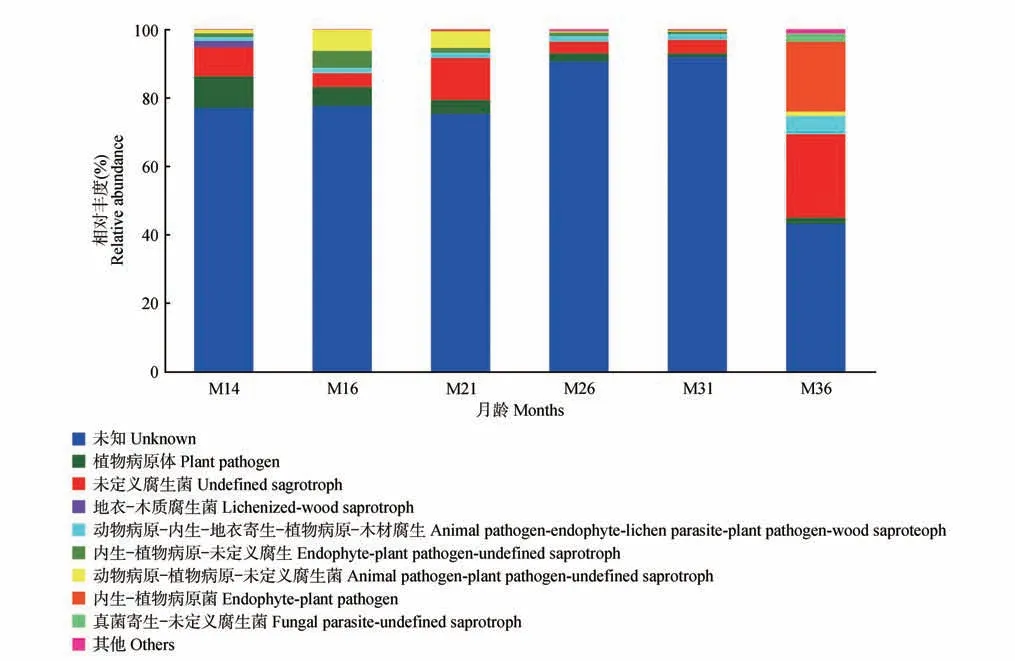
图7 不同月龄大熊猫真菌群落功能组成Fig.7 Composition of functional groups of giant pandas at different months of age
4 讨论
已有研究表明,子囊菌门是成年和老年大熊猫肠道的优势菌门,真菌多样性从高至低依次为成年、老年和亚成体;放归个体的真菌多样性和比例有明显变化,且断奶前后肠道真菌丰富度会显著改变,子囊菌门大幅下降,担子菌门大幅上升[7-11]。在大熊猫成长过程中,最初是以营养丰富的母乳为主食,成年后则以低营养高纤维的竹子为主食,营养代谢和食物代谢的变化具有独特性,开展从母乳到竹子饮食结构转变过程的肠道菌群变化研究对其保护具有重要意义。本研究采用高通量测序技术分析了14~36月龄大熊猫从幼年向亚成体过渡的真菌菌群组成和变化规律,反映了粪便中真菌的种类和丰度,研究结果可为亚成体大熊猫肠道微生态健康状况评估提供基础数据参考。
本研究中3 只大熊猫因生活史和年龄非常相似,个体间粪便真菌多样性基本没有差异,尤其是1 对双胞胎更为相似。子囊菌门是3 只大熊猫粪便中丰度最高的真菌菌门,这与以往的研究结果[8,12]基本相似。2~5 岁是大熊猫亚成体时期,2 岁时饲料有较大调整。从月龄上看,14~36 月龄是大熊猫从幼年到亚成体的过渡阶段,在饲料调整的26 月龄显现出比较明显的多样性变化,真菌多样性和丰富度在26月龄时都处于峰值。26月龄时饲喂大熊猫的奶粉量略有下降,竹笋、窝头等饲料量显著提升,这可能是26 月龄时真菌多样性和丰富度较高的原因。从真菌组成和群落功能改变来分析,36 月龄与其他月龄间相比变化较大,36 月龄时真菌组成和群落功能发生明显改变,担子菌门比例增加,未定义腐生菌和内生-植物病原菌增加,可能是36 月龄时基本不再提供奶粉所致。本研究表明饲料改变和年龄是影响大熊猫肠道真菌组成的重要因素。通过对亚成体大熊猫真菌组成变化的研究,为大熊猫肠道微生态研究提供了重要的基础数据,为疾病预防、日粮搭配等提供参考依据,以期待可以提高圈养大熊猫的福利。
猜你喜欢
小哥白尼(野生动物)(2021年7期)2021-11-20
中国比较医学杂志(2020年4期)2020-05-26
湖南畜牧兽医(2020年6期)2020-03-22
娃娃乐园·综合智能(2019年10期)2019-12-07
水生生物学报(2019年4期)2019-07-20
生物安全学报(2019年3期)2019-02-15
川北医学院学报(2019年6期)2019-02-10
学苑创造·A版(2018年7期)2018-08-07
天天爱科学(2017年12期)2017-08-15
中学生物学(2008年12期)2008-12-27
