
芹菜素对SH-SY5Y细胞内质网应激诱导凋亡的影响*
2023-11-04 11:38杨全伟黄文涛陈一璇胡松
医药导报 2023年11期
杨全伟,黄文涛,陈一璇,胡松
(武汉市第一医院药学部,武汉 430022)
神经元丢失是多种神经退行性疾病的主要诱发原因。神经损伤的临床表现主要为记忆缺陷、震颤麻痹、舞蹈症状及肌肉萎缩等功能性障碍等,从而影响记忆、运动、思考、交流、吞咽和呼吸等功能。目前,减少或控制神经元损伤或丢失的方法或药物仍然有待进一步发现。
现已证实内质网应激与神经退行性疾病发展进程密切相关。内质网内环境的稳定是实现内质网功能的基本条件,由于内质网应激常导致内质网内未折叠蛋白质或错误折叠蛋白的蓄积,诱导内质网应激标志蛋白的表达,如内质网应激球蛋白结合蛋白(globulin-binding protein,BIP)、C/EBP同源蛋白(C/EBP homologous protein,CHOP)及半胱氨酸天冬氨酸蛋白水解酶12(csyteine aspartic acic specific protease,Caspase-12)。一方面,内质网应激通过将信号逆向传入细胞核,产生转录调控,保护细胞免受内质网应激导致的损伤[1];另一方面,长期高强度内质网应激,触发内质网应激诱导的细胞凋亡发生[2],引起抗凋亡B细胞淋巴瘤-2蛋白(B cell lymphoma/leukemia-2,Bcl-2)与凋亡蛋白Bcl-2相关X蛋白(Bcl-2-associated X protein,Bax)比值改变。
芹菜素(apigenin,Api)又称芹黄素、洋芹素,是一种黄酮类单体化合物,存在于多种果蔬(苹果及芹菜等),尤以芹菜中含量最高。近年来研究发现,芹菜素具有多种生物学活性,如抗炎、抗氧化、抗肿瘤、抗衰老等作用,也可用于治疗老年痴呆、帕金森病等神经退行性疾病[3]。虽然目前有文献报道芹菜素可以通过抗氧化作用阻止1-甲基-4-苯基吡啶(1-methyl-4-phenylpyridinium,MPP+)诱导人神经母细胞瘤细胞(human neuroblastoma,SH-SY5Y)凋亡发生[4],防治帕金森病,但大部分研究集中于抗肿瘤领域,对芹菜素是否能抑制神经元丢失、尤其是内质网应激诱导的细胞凋亡发生笔者未见报道。笔者在本实验以毒胡萝卜素(thapsigargin,TG)诱导SH-SY5Y细胞产生内质网应激模型,探讨芹菜素逆转TG所致SH-SY5Y细胞内质网应激诱导凋亡的分子机制。
1 材料与方法
1.1试剂 BIP、CHOP、Bcl-2、Caspase-12、β激动蛋白(β-actin)、Bax 均购于武汉爱博泰克(ABclonal)生物科技有限公司,批号分别为A0241、A0221、A0209、A0217、AC026及A19684;芹菜素(含量≥98%)、4-苯丁酸(4-phenylbutyric acid,PBA,含量≥98%)购于上海阿拉丁生化科技有限公司,批号分别为A106676、#F1828076;TG购于默克中国有限公司(批号:67526-95-8,含量≥98%);改良Eagle(Dulbecco's modification of Eagle's medium,DMEM)培养基购于美国HyClone公司(批号:D5671);胎牛血清购于Gibco公司(批号:10099);二辛可宁酸(bicinchoninic acid,BCA)测定蛋白定量试剂盒、细胞计数试剂-8(cell counting kit-8,CCK-8)检测试剂盒购于上海翊圣生物科技有限公司,批号分别为2021-A、40210ES10;强裂解液(radio immunoprecipitation assay,RIPA)购于杭州弗德生物科技有限公司(批号:FD008);SH-SY5Y细胞购于武汉谷歌生物有限公司,鉴定报告由武汉谷歌生物有限公司提供。
1.2仪器 水平摇床(海门市其林贝尔仪器制造有限公司,型号:TS-1);细胞培养箱(Thermo公司,型号:360);电泳槽、电转膜仪(Bio-Rad公司,型号:165-8004);超净工作台(苏州净化公司,型号:SW-CT-2D);多功能酶标仪(美国Bio-Tek公司,型号:Synergy-2);化学发光凝胶成像系统(Bio-Rad公司,型号:ChemiDoTMXRS+)。
1.3细胞的培养 用含10%胎牛血清、青霉素(100 U·mL-1)和链霉素(100 mg·mL-1)的DMEM高糖培养基培养SH-SY5Y细胞,培养条件:37 ℃,5%二氧化碳(CO2),恒温恒湿。隔天换培养基1次,细胞融合度达到80%传代,进行下一步实验。
1.4实验分组 以TG制备内质网应激模型,研究内质网应激所致内源性凋亡途径,如浓度和时间依赖性TG诱导内质网应激标记物BIP显著增加,则判定为内质网应激造模成功。实验分组[5]:对照组(CON组,2%胎牛血清DMEM高糖培养基)、TG组(以2%胎牛血清DMEM高糖培养基配制TG)、TG+芹菜素组(以2%胎牛血清DMEM高糖培养基配制TG与芹菜素)。
为考察TG造模最佳浓度与时间,设置TG浓度梯度为0.5、1.0、2.0、3.0 μmol·L-1;TG处理细胞时间分别为4、6、8、10 h。另外,当细胞贴壁密度达80%时,设置预防性给予芹菜素浓度分别为10、20、30 μmol·L-1,检测细胞活力,观察芹菜素在TG所致内质网应激发生的作用。用2%胎牛血清DMEM高糖培养基配制4-丁苯酸(PBA)浓度为1.5 mmol·L-1溶液,-4 ℃保存备用。PBA是内质保护剂,可抑制内质网应激。
1.5CCK-8法检测细胞活性[6]取对数生长期细胞,调整细胞浓度为1×105个·mL-1,每孔100 μL接种于96孔板,在37 ℃、5%CO2条件下培养约24 h,按照“1.4节”分组方法给药,给药完成后,每孔加入CCK-8溶液10 μL,在37 ℃、5%CO2培养箱中给药2 h,酶标仪在波长450 nm处测定各孔吸光度(A)值。细胞活力(%)=[A(加药)-A(空白)]/[A(0加药)-A(空白)]×100%。A(加药)、A(空白)及A(0加药):参照说明书指导方法,按是否含有细胞、CCK-8溶液和药物溶液,测定A值。
1.6蛋白免疫印迹法(western blotting,WB)检测内质网应激蛋白及凋亡蛋白表达[4]取对数生长期细胞,调整细胞浓度为2.5×105个·mL-1,接种于6孔板,37 ℃、5%CO2条件下,过夜培养12~14 h后,待次日细胞密度为70%~80%时,给予10 μmol·L-1芹菜素预处理1 h;随后,1.0 μmol·L-1TG给药10 h。收集细胞,PBS洗3次,加入RIPA,冰浴裂解15 min,收集细胞裂解液于1.5 mL离心管内;12 000 r·min-1离心10 min(r=6 cm),取上清液,采用BCA蛋白定量检测法,酶标仪波长562 nm条件下测定蛋白含量。定量后的蛋白样品采用10%十二烷基硫酸钠-聚丙烯酰胺凝胶(sodium dodecyl sulfate polyacrylamide gelelectrophoresis,SDS-PAGE)进行蛋白电泳,蛋白上样量10~20 μg。电泳条件起始电压60 V,到达分离胶后调整为120 V。转膜条件为聚偏二氟乙烯(polyvinylidene fluoride,PVDF)膜恒流(电流300 mA)转膜2 h。室温下5%脱脂奶粉封闭1 h,4 ℃过夜,给药一抗BIP(1:1000)、CHOP(1:1 000)、Bcl-2(1:1 000)、CASPASE-12(1:1 000)、Bax(1:1 000)以及β-actin(1:5 000)蛋白,TBST缓冲液洗3次,给药二抗,ECL化学发光显影,化学发光凝胶成像系统分别检测蛋白表达水平。
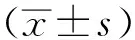
2 结果
2.1芹菜素对TG诱导的SH-SY5Y细胞活力的影响 结果显示,与CON组比较,随着TG浓度增加,SH-SY5Y细胞活力呈剂量依赖性下降(P<0.01),当TG浓度>1.0 μmol·L-1时,细胞活力未见明显改变,故选择TG浓度1.0 μmol·L-1作为内质网应激诱导细胞凋亡的造模浓度。TG时间依赖性实验结果显示,与CON组比较,随着TG处理时间延长,细胞活力均明显下降(P<0.01)。TG处理10 h后,细胞活性降至最低。因此,选择TG给药时间为10 h。分别给予10、20、30 μmol·L-1芹菜素预处理细胞1 h,给予1.0 μmol·L-1TG诱导内质网应激下,确定10 μmol·L-1芹菜素预处理可明显改善细胞活力(P<0.01)。因此本实验选择芹菜素预处理浓度为10 μmol·L-1,结果见图1。
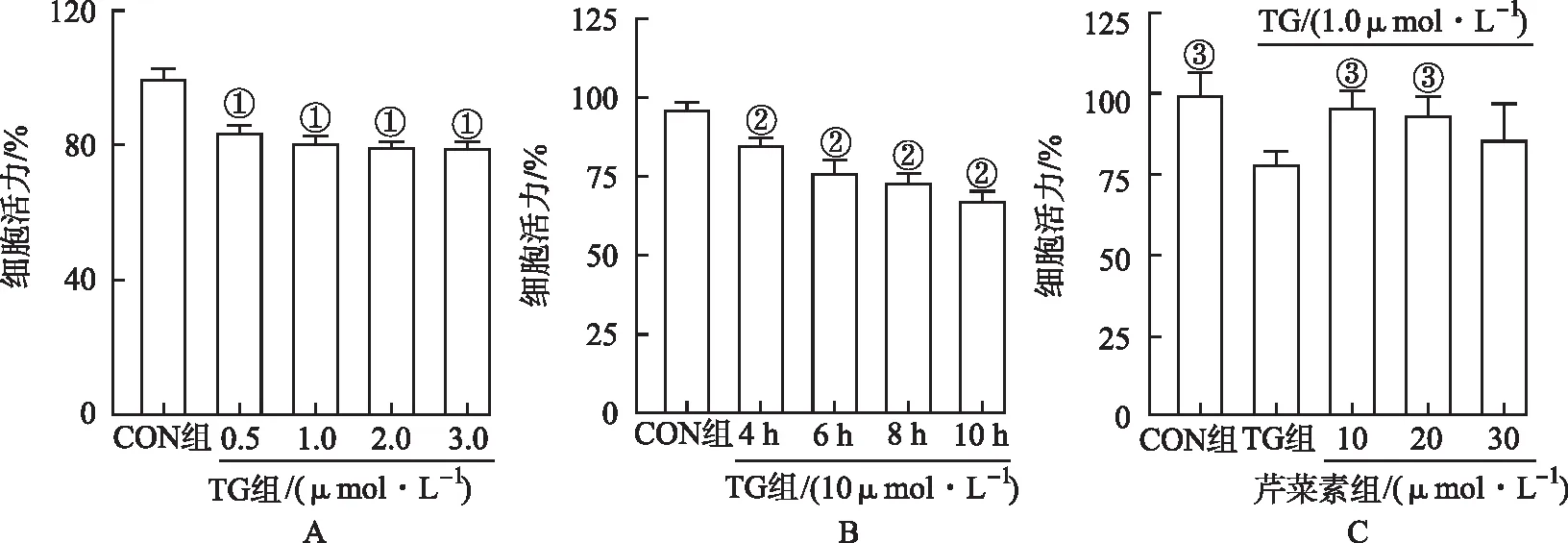
①与CON组比较,t=5.31~7.14,P<0.01;②与CON组比较,t=3.13~9.62,P<0.01 ;③与TG组比较,t=0.39~6.47,P<0.01。图1 芹菜素对TG诱导的SH-SY5Y细胞活力的影响 ① Compared with CON group,t=5.31-7.14,P<0.01;② Compared with CON group,t=3.13-9.62,P<0.01;③ Compared with TG group,t=0.39-6.47,P<0.01.Fig.1 Effect of apigenin on the viability of SH-SY5Y cells induced by TG
2.2芹菜素对内质网应激相关蛋白BIP表达的影响 与CON组比较,TG组BIP表达明显上调(P<0.01);与TG组比较,芹菜素预处理组BIP表达降低(P<0.05),TG+PBA组内质网应激被明显抑制(P<0.05),见图2。

①与CON组比较,t=5.25,P<0.01;②与TG组比较,t=3.81,2.397,P<0.05。图2 芹菜素对BIP蛋白表达的影响① Compared with CON group,t=5.25,P<0.01;② Compared with TG group,t=3.81,2.397,P<0.05.Fig.2 Effect of apigenin on BIP protein
2.3芹菜素对内质网应激诱导蛋白CHOP表达的影响 WB结果显示,与CON组比较,TG组CHOP蛋白表达显著增多(P<0.01);与TG组比较,TG+芹菜素组CHOP表达显著降低(P<0.05),见图3。
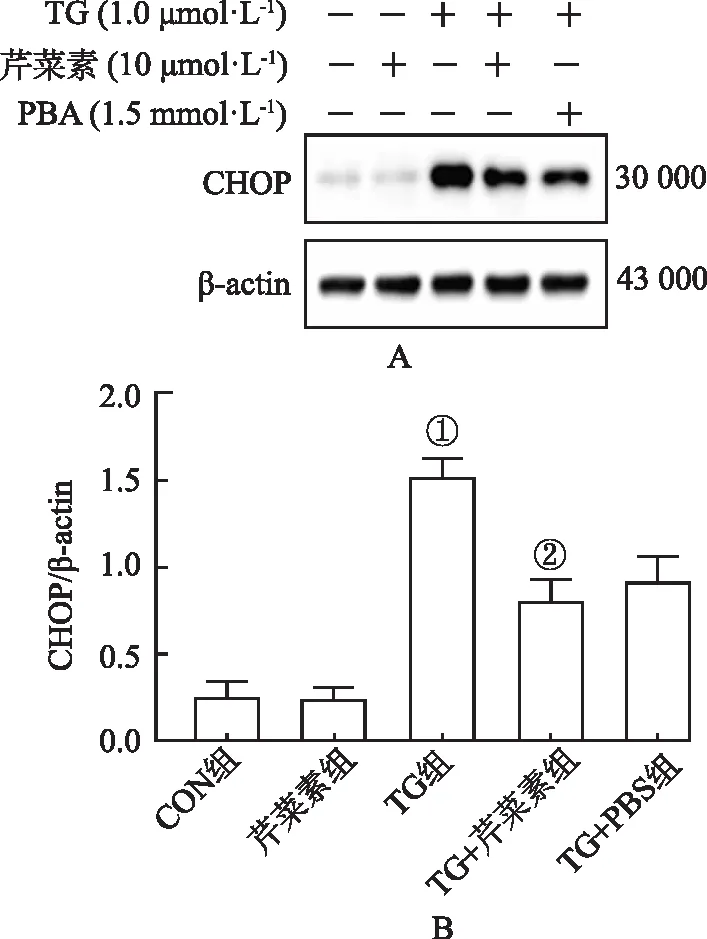
①与CON组比较,t=9.43,P<0.01;②与TG组比较,t=4.61,P<0.05。图3 芹菜素对CHOP蛋白表达的影响① Compared with CON group,t=9.43,P<0.01;②Compared with TG group,t=4.61,P<0.05.Fig.3 Effect of apigenin on CHOP protein
2.4芹菜素对内质网应激诱导的Caspase-12蛋白表达的影响 与CON组比较,TG组Caspase-12蛋白表达明显增多(P<0.05);与TG组比较,TG+芹菜素组表达趋势降低(P<0.05),见图4。
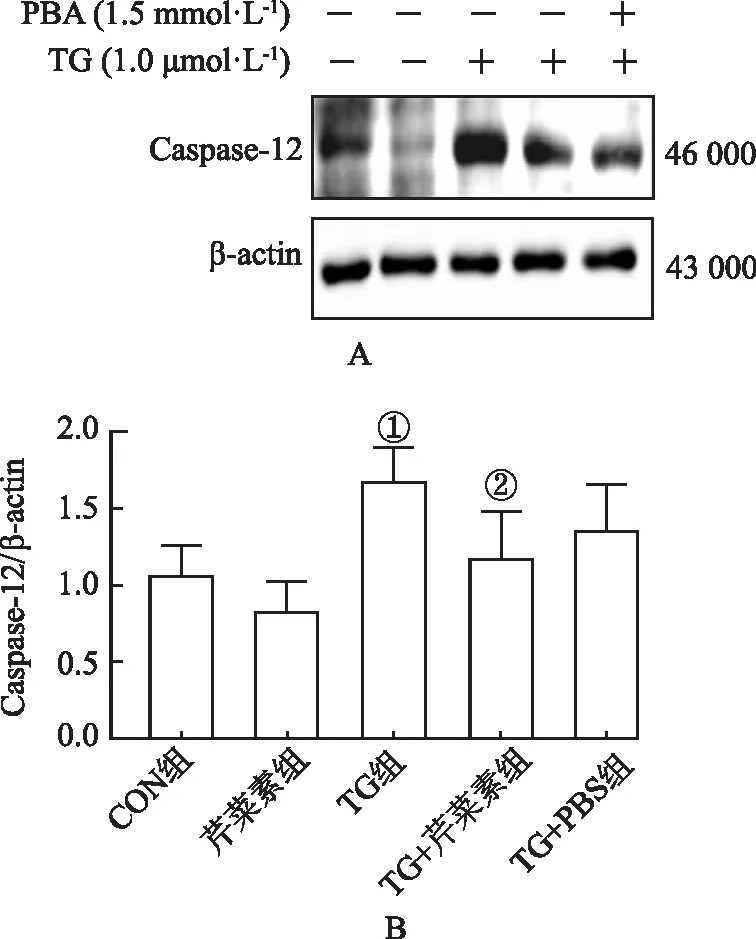
①与CON组比较,t=3.02,P<0.05;②与TG组比较,t=4.56,P<0.05。图4 芹菜素对Caspase-12蛋白的影响①Compared with CON group,t=3.02,P<0.05;②Compared with TG group,t=4.56,P<0.05.Fig.4 Effect of apigenin on Caspase-12 protein
2.5芹菜素对内质网应激诱导的Bcl-2/Bax蛋白表达的影响 与CON组比较,TG组抗凋亡蛋白Bcl-2表达明显减少 (P<0.05),促凋亡蛋白Bax表达增多,Bcl-2/Bax比值显著下降(P<0.05)。与TG组比较,TG+芹菜素组Bcl-2表达明显上调(P<0.05),Bax表达也有下调趋势,Bcl-2/Bax比值显著增加 (P<0.05),结果见图5。
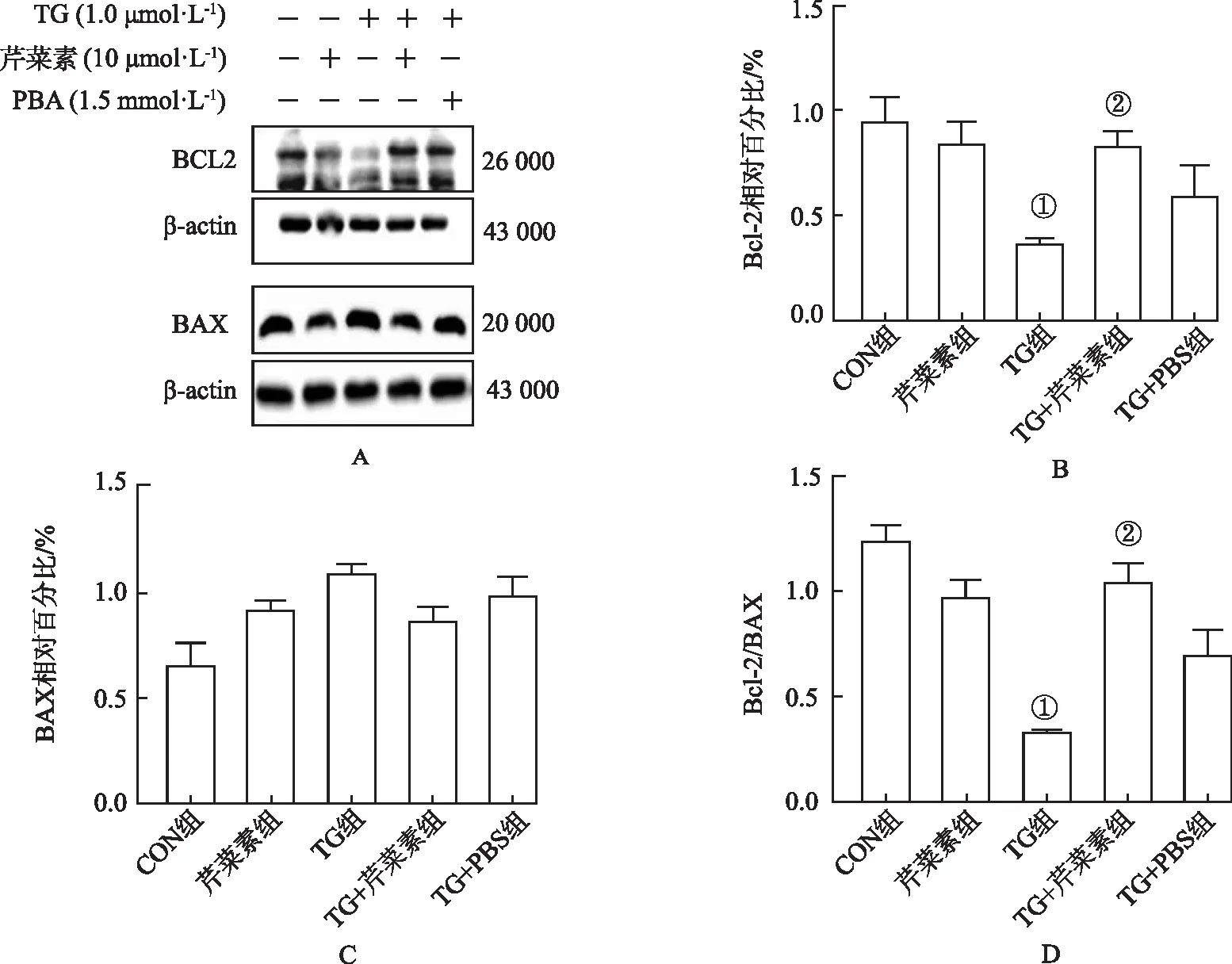
①与CON组比较,t=3.31~5.67,P <0.05;②与TG组比较,t=2.84~5.16,P<0.05。图5 芹菜素对BCL2/BAX蛋白表达的影响① Compared with CON group,t=3.31-5.67,P<0.05;② Compared with TG group,t=2.84-5.16,P<0.05.Fig.5 Effect of apigenin on BCL2/BAX protein
3 讨论
神经退行性疾病的发生机制包括活性氧生成、线粒体功能障碍、炎症、折叠蛋白积累和泛素蛋白酶体系统功能障碍等。其中内质网是蛋白质合成和折叠的场所,其错误折叠或合成障碍将诱发内质网应激。为了克服内质网应激,细胞内形成一套特定的信号处理通路,称为内质网应激反应通路[7]。BIP作为分子伴侣蛋白和内质网应激信号通路的主要调控因子,利用细胞降解机制使未折叠的蛋白质重新折叠或降解。研究发现,包括PERK/Caspase-12激酶通路和ATF6/CHOP通路等都参与BIP介导的内质网应激的发生[8]。随着高强度或长时间内质网应激可诱导内源性细胞凋亡发生,在此过程中,细胞调动一系列精确机制,来保护细胞内蛋白质的折叠和组装的动态平衡[9]。
文献[10]报道,SH-SY5Y细胞系作为神经退行性疾病研究的常用细胞系,主要用于研究帕金森病、阿尔茨海默病等神经退行性疾病的病理机制。本研究显示,在内质网应激诱导剂TG(1.0 μmol·L-1)处理时间为10 h时,SH-SY5Y细胞活力显著下降,而1.0 μmol·L-1芹菜素预处理1 h可恢复TG所致SH-SY5Y细胞活力下降,起到保护作用。此外,还发现TG可引起内质网应激相关蛋白BIP、CHOP、Caspase-12表达增多,Bcl-2表达明显降低,而Bax表达未见明显改变。研究证实,TG可引起内质网应激,并引起细胞凋亡。
前期研究认为,芹菜素可以抑制淀粉样蛋白诱导的SH-SY5Y细胞凋亡发生[11],淀粉样蛋白也可以作为一种内质网应激诱导剂,用于各种神经元细胞模型。笔者发现,在TG诱导的内质网应激中,芹菜素预处理组可以明显逆转BIP、CHOP、Caspase-12、Bcl-2蛋白表达以及Bcl-2/Bax比值,但对Bax的作用不明显。提示芹菜素可能通过抑制TG所致内质网应激发生,阻止细胞凋亡,从而抑制神经细胞损伤。然而,芹菜素抑制内质网应激诱导细胞凋亡通路的具体分子机制仍值得进一步研究。
综上所述,芹菜素能够逆转TG诱导内质网应激,恢复内质网应激所致细胞活力改变,并抑制内质网应激引起的细胞凋亡。本研究为芹菜素治疗神经退行性疾病提供了实验参考。
猜你喜欢
解放军医学杂志(2021年12期)2022-01-18
科学大众(2021年9期)2021-07-16
现代临床医学(2021年1期)2021-01-26
基层中医药(2018年7期)2018-12-06
娃娃乐园·综合智能(2018年11期)2018-07-10
老年医学与保健(2017年6期)2017-02-06
安徽医科大学学报(2016年12期)2017-01-15
中华肩肘外科电子杂志(2017年1期)2017-01-11
中外医疗(2015年5期)2016-01-04
中国当代医药(2015年33期)2015-03-01
