
古老月季叶绿体基因组密码子分析
2023-10-24 10:36:32程培蕾严陶韬高静瑶张彤钰
江苏农业科学 2023年17期
关键词:月季
程培蕾 严陶韬 高静瑶 张彤钰

摘要:我国古老月季月月粉具有连续开花、重瓣和标志性茶香等优良性状,是重要的育种材料。为明确月月粉叶绿体基因组密码子的特点,从NCBI网站获得了完整的叶绿体基因组信息,并对所有基因的密碼子使用偏性进行计算分析。结果表明,月月粉叶绿体包含53个基因,1~3位密码子GC含量平均值分别为47.08%、39.54%、29.14%,绝大多数密码子前2位的GC含量较高。编码区有效密码子数平均值为46.86,表明其偏性较弱。相关性分析显示密码子第3位与前2 位存在明显差异,可能对密码子使用偏性具有关键作用。29个RSCU>1的密码子中以A、U 结尾的有28个。中性绘图分析表明,GC12与GC3间相关系数为0.088,相关性不显著,ENC-plot分析、PR2-plot分析结果表明自然选择在月月粉叶绿体基因组密码子使用模式中起决定性作用,并筛选到16个最优密码子。结果可为月季遗传转化过程中提高外源基因的表达提供有用信息。
关键词:月季;叶绿体基因组;密码子偏好性;最优密码子
中图分类号:S685.120.1 文献标志码:A 文章编号:1002-1302(2023)17-0034-07
三联体密码子通过简并性方式编码氨基酸[1],由多个同义密码子编码相同氨基酸的现象极其普遍,仅由单个密码子编码的氨基酸数量极少,已知的只有蛋氨酸(Met)和色氨酸(Trp)[2]。不同植物基因组中任意氨基酸同义密码子的选择是非随机的,这被称为同义密码子使用偏好性[3]。密码子使用偏好性不仅因物种、家族或生物界内的群体而异,也因生物体内基因的不同而异。自然选择和突变在生物体密码子使用偏好性方面起着决定性作用[4-5]。密码子使用偏好性在多种生命过程中发挥作用,例如蛋白质翻译和折叠等过程,也反映了物种或基因的起源、突变模式和进化[6-7]。对密码子使用偏好性的研究可以揭示生物之间的系统发育关系,水平基因转移,基因的分子进化,确定驱动其进化的影响因素,并且对外源基因密码子的优化可以在一定程度上提高转基因育种中的外源基因表达水平[8]。
多数植物叶绿体基因组高度保守,广泛应用于植物间的进化规律、遗传关系、叶绿体基因表达和叶绿体转化研究[9]。近年来,高通量测序技术丰富了叶绿体基因组数据库,与植物基因组结构和系统发育分析相关的研究也更加广泛和深入。越来越多的园艺作物开展了叶绿体基因组密码子使用偏好性分析,例如山茶、睡莲、含笑、连翘、云南油杉、木薯等[10-15]。
月季(Rosa chinensis)是栽培品种极其丰富的木本花卉,不仅具有很高的观赏价值,更可作为食品添加剂、熏蒸剂、化妆品和传统医疗产品使用[16-17]。我国古老月季月月粉是重要的育种材料,具有多种优良性状,包括连续开花、重瓣和标志性茶香[18]。随着其全基因组序列的公布[19-20],月月粉成为研究木本植物花发育、花颜色、气味释放和春化反应分子机制的理想模式物种。
本研究从月季叶绿体编码序列的角度,系统分析了古老月季月月粉叶绿体基因组的密码子组成、密码子偏好性情况及影响因素,并筛选出最优密码子。本研究揭示了月月粉密码子使用偏好性,以期为进一步研究月季的遗传进化和转基因提供参考方向。
1 材料与方法
1.1 材料
2022年8月于苏州农业职业技术学院进行相关数据分析。月季月月粉完整的叶绿体基因组从NCBI(https://www.ncbi.nlm.nih.gov)网站获得,GenBank登录编号为MH332770。叶绿体基因组大小为156 591 bp,根据各基因的注释结果获得不重复的85条CDS(coding DNA sequence)。对获得的原始编码序列进行筛选分析,保留了序列长度 300 bp 以上,起始为ATG、结尾为TAG、TGA、TAA的53条非重复CDS用于下一步分析。
1.2 方法
1.2.1 密码子组成分析 参照梁湘兰等的方法统计了53条月月粉叶绿体基因密码子数量和不同位点的GC含量[21]。有效密码子数 (effective number of codon,ENC)测定参考李魁印等的方法,ENC数值用以表征同义密码子使用偏度,数值区间为20~61,ENC值=20表示偏度最高,随数值增加偏度变弱[22]。RSCU(relative synonymous codon usage)指相对同义密码子使用度,当数值高于1时表示实际使用频率高于其他密码子,确定为高频密码子,其计算方法参照文献[23]。
1.2.2 密码子中性绘图分析 月月粉叶绿体基因组密码子中性绘图的横、纵坐标分别为GC3值和GC1、GC2平均值,散点表示相应的基因。横纵坐标间的相关关系代表了密码子偏好性的主要影响因素,当两者呈现显著相关时,不同位点的碱基使用差别不明显,突变成为决定性因素;而当两者呈现不显著相关时,碱基组成差异性与回归系数呈负相关关系,自然选择成为决定性影响因素,绘图方法参照文献[24]。
1.2.3 ENC-plot分析 ENC-plot分析参考杨祥燕等的方法[25]。ENC-plot绘图可以表征突变和自然选择在密码子偏好性中的作用,以散点到标准曲线的距离为基本划分依据,当二者距离近时代表偏好性主要以突变影响为主,而当两者距离相对较远时,偏好性则主要取决于自然选择对其的影响。以ENC比值频数的分布情况来量化突变和自然选择的影响,其比值具体计算公式为ENC差值(ENC期望值-ENC实际值)与ENC 期望值之比,其中,ENC期望值基本计算公式:2+GC3+29/[GC23+(1-GC3)2][25]。
1.2.4 PR2-plot分析 统计月月粉53个叶绿体基因各密码子第 3 位点的A、T、C、G分布情况,PR2偏倚分析(PR2-bias plot analysis)图中,A3/(A3+T3)与G3/(G3+C3)的取值分别被设置为纵、横坐标,纵横坐标的中心交界点数值为0.5,该点A、T相同,C、G也相同,即密码子无偏性使用,其他散点距离中心点的矢量距离代表偏好程度[25-26]。
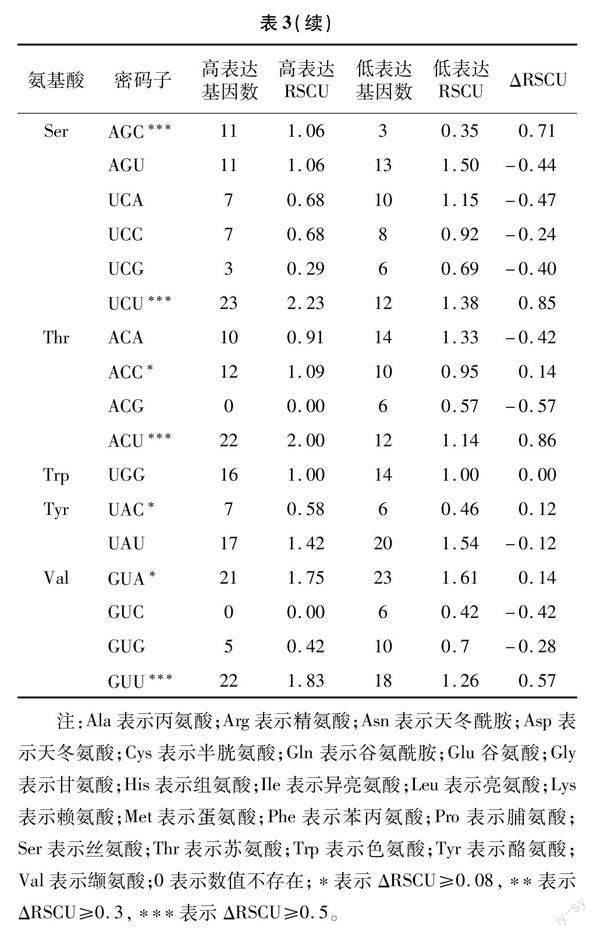
1.2.5 最优密码子筛选 将ENC值由低到高排序,最低和最高的10%分别作为月月粉叶绿体基因高、低偏性库,RSCU>1表示高频密码子,高、低偏性库间差值ΔRSCU(高表达RSCU-低表达RSCU)在0.08及以上时表示高表达优越密码子,最优密码子的筛选标准为高频率密码子和高表达优越密码子的交集[27]。
2 结果与分析
2.1 月月粉密码子偏好性相关分析
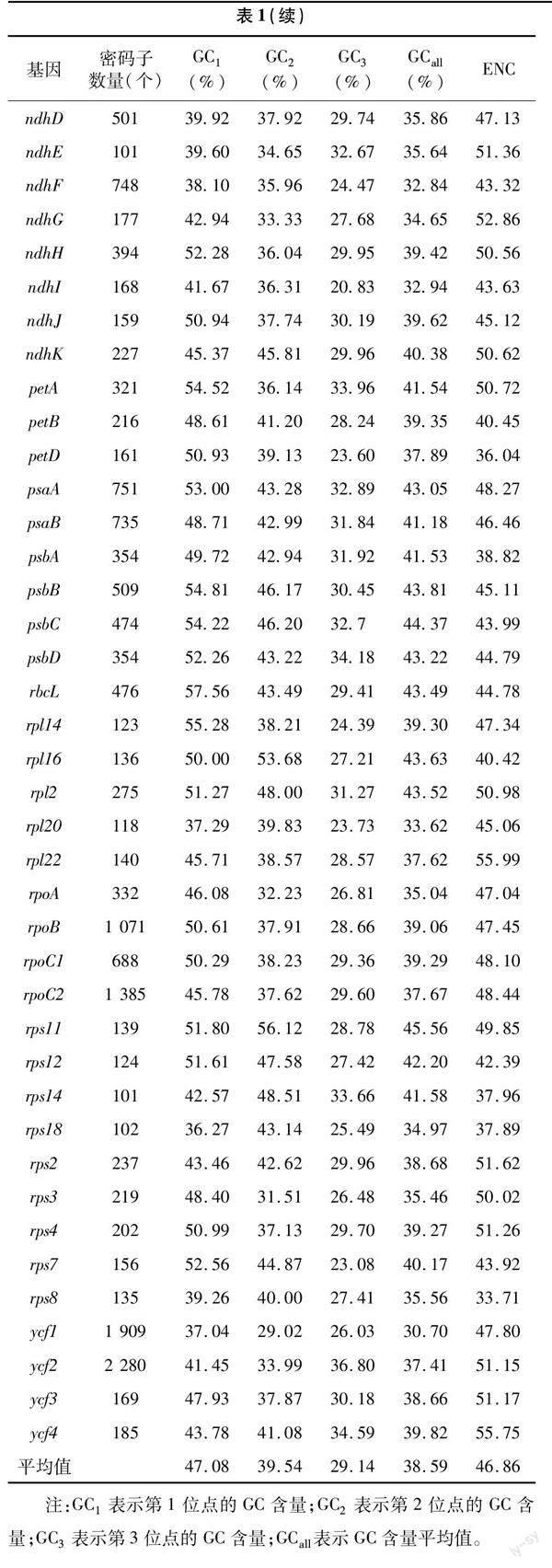
月季叶绿体基因组序列下载自NCBI 数据库(GenBank登录号:MH332770),53个叶绿体基因的密码子偏好性相关参数分析结果见表1。其密码子的GC含量平均值为38.59%,其中,密码子由第1位点至第3位点的GC含量平均值分别为47.08%、39.54%、29.14%,1位GC含量最高的基因clpP为57.87%,2位GC含量最高的基因rps11为56.12%,第3位点GC含量最高的基因ycf2为36.80%,可见,密码子GC含量在不同位点差别较大,且绝大多数基因密码子前2位的GC含量较高,第3位的GC含量较低,表明叶绿体基因更加偏好使用A 或U结尾。此外,月季53个叶绿体基因ENC平均值为46.86,最大值和最小值分别为55.99和33.71,月月粉53个编码序列密码子的主要参数分析结果表明密码子偏性较弱。
对月季叶绿体基因密码子参数的相关性分析结果(图1)表明,GC1与GC2极显著正相关,GC1与GC3以及GC2与GC3均不显著相关,GCall与3个位点的GC值(GC1、GC2、GC3)均为极显著正相关,表明第1位与第2位碱基呈现极高的相似度,但这两者与第3 位均存在较大差异;ENC与GC1无显著相关关系,与GC2和GC3相关性较强,分别为显著和极显著相关,猜测密码子使用的偏好性主要受到GC3含量的影响;密码子数与其他参数均不显著相关,说明月季叶绿体中密码子碱基组成不影响密码子数。
月月粉53个叶绿体基因RSCU分析结果见图2,这些基因由 21 304个非终止密码子组成。Ile(AUA)的RSCU值为0.925 2,Leu(CUA)的RSCU值为0.748 8,其他以A或U结尾的密码子RSCU值均大于1.0。共筛选到29个RSCU值大于1.0的密码子,其中以U结尾的有16个、以A 结尾的有12个、以G结尾的有1个。表明月月粉叶绿体基因密码子更加偏向于在末位选择使用A或U。
2.2 密码子碱基组成的差异性
中性绘图分析结果(图3)表明,月季叶绿体基因组GC分布较窄,GC3 的变化范围为20.83%~36.80%,GC12 的取值范围为32.83%~53.96%。GC12与GC3的相关系数为r=0.08 8(r2=0.007 8),相关性不显著,且基因均位于对角线的上方,猜测自然选择作为影响密码子偏好性的主要原因发挥作用。以基因的功能为依据对其进行分类,53条序列可以进一步分为15类,其中保守的假定叶绿体ORF(conserved hypothetical chloroplast,ORF)和被膜蛋白(envelope membrane protein)靠近对角线, 其他基因均距离对角线较远。
2.3 ENC-plot绘图分析
对叶绿体基因的密码子进行ENC-plot分析(图4),标准曲线下方聚集了大多数叶绿体基因,且距离较远,标准曲线两侧仅均匀分布了部分基因,表示月季叶绿体基因组密码子偏好性受突变的影响较小,受自然选择的影响较大。对月季叶绿体基因的ENC频数分布进行了统计(表2),有17个基因分布在-0.05~0.05区间,基因频率占比为32%,有25个基因分布在0.05~0.15区间,基因频率占比为47%,表明实际的ENC数值与预期值之间差距较大,因此主要是自然选择影响了月月粉叶绿体基因组密码子的偏好性。
2.4 月季叶绿体基因组PR2-plot绘图分析
对月季53个叶绿体基因进行PR2-plot 绘图分析(图5),结果表明密码子使用情况为A3/A3+T3(平均值为0.460)低于G3/G3+C3(平均值为0.543)。这些基因在平面4个区域内呈不均勻分布,其中右下角区域基因数量最多,而左上角区域基因数量最少。此外,4种碱基在使用频率方面也存在差异,G的使用频率高于C,T的使用频率高于A;且基因小核糖体亚基蛋白和光系统Ⅱ亚单位偏离中心点(A=T且C=G)的矢量距离最远。嘧啶与嘌呤间的使用频率差异也进一步证明, 自然选择决定了月月粉叶绿体基因组密码子使用模式,而突变在这一过程中发挥的作用较弱。
2.5 最优密码子确定
月季53个叶绿体基因的高、低偏性库,以及2个库的RSCU值统计结果见表3。高表达优越密码子的筛选标准为高、低偏性库之间差值在0.08以上, 共筛选到21个。进一步分析了这21个高表达优越密码子的末位碱基情况,绝大多数(14个)以A/U结尾,以C结尾的次之(6个),以G结尾的最少(仅1个)。将RSCU>1的密码子作为高频密码子,共筛选到29个,其中28个以A/U 结尾,仅1个以G 结尾。高频密码子与高表达优越密码子的交集作为月月粉最优密码子,本研究中共获得了16个最优密码子,分别为GCA、GCU、CGU、UGU、CAA、GAA、GGU、AUU、UUA、UUG、AGC、UCU、ACC、ACU、GUA、GUU,这些最优密码子第3 位碱基有5个为A,8个为U,2个为C,1个为G。
3 讨论与结论
古老月季月月粉叶绿体基因组的密码子使用偏好性对于探讨其进化过程至关重要,有助于该物种的分子育种应用。尽管先前已经有关于大花香水月季叶绿体基因组密码子使用特点的相关研究,两者均为二倍体多年生植物,但其基本性状差异较大,最典型的差异为月月粉可以周年连续开花,而大花香水月季为早春一次开花型[28-29]。此外随着全基因组信息的公布,月月粉成为研究木本植物花发育、花颜色、气味释放和春化反应分子机制的理想模式物种[19-20]。本研究旨在系统分析古老月季月月粉叶绿体基因组的有效密码子数、相对同义密码子使用、密码子偏好性及其影响因素,这将为优化月月粉分子育种提供一定参考。
同义密码子使用偏好性是生物类群中常见的现象,一般指不同物种之间或同一物种基因组内密码子出现频率的差异[30]。密码子第3位碱基的同义突变不能改变氨基酸的类型,但仍被认为是决定氨基酸类型的一个重要特征,且其GC含量决定了密码子偏好性[31-32]。本研究中,月月粉叶绿体基因组GC含量平均值为38.59%,密码子第3位的GC含量为29.14%,密码子末位碱基使用A、U的频率高于G、C,在多个物种中存在相似的现象,包括菠萝[25]、苦马豆[21]、文心兰[33]等。对密码子使用模式的两大主要影响因素(碱基突变和自然选择)进行确认,当突变为绝对原因时,则应按比例使用4个碱基(A、C、G 和 T)[34]。然而,月月粉叶绿体基因组中的大多数编码序列都是A、U结尾,这表明自然选择是对其密码子使用模式的更大影响因素。
中性理论认为碱基突变和自然选择以中性或接近中性的形式影响了密码子第3个碱基的使用[4]。如果自然选择作为主要的影响因素,则第3个位点的GC含量往往分布在一个小范围内,且与前2位没有显著相关性[35]。在本研究的中性分析结果显示,月月粉叶绿体基因组的密码子GC3 的变化范围较小(20.83%~36.80%),GC12 与GC3 的相关系数为r=0.088(r2=0.007 8),相关性不显著,表明其密码子使用偏好性主要受自然选择的影响。此外,月月粉叶绿体ENC频数比值结果显示,其主要的分布区间为0.05~0.15,实际数值与预期数值之间存在较大的差别[25];PR2-plot分析结果中的嘌呤A/G与嘧啶T/C间的差异都表明自然选择在月月粉密码子使用偏好性中的决定性作用,而突变发挥的作用较弱[24-28]。自然选择、突变等因素的作用,促进了密码子使用偏性的发生,而最优密码子形成的重要条件就是强烈的正向选择和突变压力[36]。本研究筛选到16個最优密码子,这些最优密码子将为月月粉遗传转化过程中提高基因的表达效率提供有用信息。
参考文献:
[1]Grantham R,Gautier C,Gouy M,et al. Codon catalog usage and the genome hypothesis[J]. Nucleic Acids Research,1980,8(1):49-62.
[2]Guan D L,Ma L B,Khan M S,et al. Analysis of codon usage patterns in Hirudinaria manillensis reveals a preference for GC-ending codons caused by dominant selection constraints[J]. BMC Genomics,2018,19(1):542.
[3]Wright F. The ‘effective number of codons used in a gene[J]. Gene,1990,87(1):23-29.
[4]Sharp P M,Stenico M,Peden J F,et al. Codon usage mutational bias,translational selection,or both?[J]. Biochemical Society Transactions,1993,21(4):835-841.
[5]Lesnik T,Solomovici J,Deana A,et al. Ribosome traffic in E.coli and regulation of gene expression[J]. Journal of Theoretical Biology,2000,202(2):175-185.
[6]Supek F. The code of silence:widespread associations between synonymous Codon biases and gene function[J]. Journal of Molecular Evolution,2016,82(1):65-73.
[7]Athey J,Alexaki A,Osipova E,et al. A new and updated resource for codon usage tables[J]. BMC Bioinformatics,2017,18(1):391.
[8]Parvathy S T,Udayasuriyan V,Bhadana V. Codon usage bias[J]. Molecular Biology Reports,2022,49(1):539-565.
[9]Wicke S,Schneeweiss G M,DePamphilis C W,et al. The evolution of the plastid chromosome in land plants:gene content,gene order,gene function[J]. Plant Molecular Biology,2011,76(3):273-297.
[10]Wang Z J,Cai Q W,Wang Y,et al. Comparative analysis of codon bias in the chloroplast genomes of Theaceae species[J]. Frontiers in Genetics,2022,13:824610.
[11]毛立彦,黄秋伟,龙凌云,等. 7种睡莲属植物叶绿体基因组密码子偏好性分析[J]. 西北林学院学报,2022,37(2):98-107.
[12]周 涛,杨 林,舒军霞,等. 3种含笑属植物叶绿体基因组密码子偏好性分析[J]. 西部林业科学,2022,51(3):91-100.
[13]宋 芸,贾孟君,曹亚萍,等. 连翘叶绿体基因组特征分析[J]. 园艺学报,2022,49(1):187-199.
[14]李江飞,王 瑜,颜廷雨,等. 云南油杉叶绿体基因组密码子偏好性分析[J]. 中南林业科技大学学报,2022,42(4):30-39.
[15]赵 英,付海天,金 刚,等. 木薯叶绿体基因组密码子偏好性分析[J]. 江西农业学报,2022,34(4):46-52.
[16]Fougère-Danezan M,Joly S,Bruneau A,et al. Phylogeny and biogeography of wild roses with specific attention to polyploids[J]. Annals of Botany,2015,115(2):275-291.
[17]Friedman H,Rot I,Agami O,et al. Edible flowers:new crops with potential health benefits[J]. Acta Horticulturae,2007(755):283-290.
[18]Bendahmane M,Dubois A,Raymond O,et al. Genetics and genomics of flower initiation and development in roses[J]. Journal of Experimental Botany,2013,64(4):847-857.
[19]Saint-Oyant L H,Ruttink T,Hamama L,et al. A high-quality genome sequence of Rosa chinensis to elucidate ornamental traits[J]. Nature Plants,2018,4(7):473-484.
[20]Raymond O,Gouzy J,Just J,et al. The Rosa genome provides new insights into the domestication of modern roses[J]. Nature Genetics,2018,50(6):772-777.
[21]梁湘蘭,郭 松. 苦马豆叶绿体基因组密码子偏好性分析[J]. 西北林学院学报,2022,37(2):121-126.
[22]李魁印,张海玲,张 鸿,等. 荨麻叶绿体基因组密码子偏好性分析[J]. 种子,2022,41(2):51-58.
[23]何亚玲,彭业军,李 锦,等. 天山雪莲叶绿体基因组密码子偏好性分析[J]. 石河子大学学报(自然科学版),2022,40(1):84-92.
[24]原晓龙,李云琴,张劲峰,等. 降香黄檀叶绿体基因组密码子偏好性分析[J]. 广西植物,2021,41(4):622-630.
[25]杨祥燕,蔡元保,谭秦亮,等. 菠萝叶绿体基因组密码子偏好性分析[J]. 热带作物学报,2022,43(3):439-446.
[26]原晓龙,刘 音,康洪梅,等. 蒜头果叶绿体基因组密码子偏好性分析[J]. 西南林业大学学报(自然科学),2021,41(3):15-22.
[27]罗 茜,方 正,余 婕,等. 圆锥南芥叶绿体基因组特征及密码子偏好性分析[J]. 分子植物育种,2022,20(7):2261-2270.
[28]牛 元,徐 琼,王嵛德,等. 大花香水月季叶绿体基因组密码子使用偏性分析[J]. 西北林学院学报,2018,33(3):123-130.
[29]Guo X L,Yu C,Luo L,et al. Comparative transcriptome analysis of the floral transition in Rosa chinensis ‘Old Blush and R.odorata var. gigantea[J]. Scientific Reports,2017,7(1):6068.[HJ2mm]
[30]Wei L,He J,Jia X,et al. Analysis of codon usage bias of mitochondrial genome in Bombyx mori and its relation to evolution[J]. BMC Evolutionary Biology,2014,14(1):262.
[31]Ingvarsson P K. Gene expression and protein length influence codon [JP+3]usage and rates of sequence evolution in Populus tremula[J].Molecular Biology and Evolution,2007,24(3):836-844.
[32]Ye Y,Ni Z,Bai T,et al. The analysis of chloroplast genome codon usage bais in Pinus massoniana[J]. Genomics and Applied Biology,2018,37(10):4464-4471.
[33]李冬梅,吕复兵,朱根发,等. 文心兰叶绿体基因组密码子使用的相关分析[J]. 广东农业科学,2012,39(10):61-65.
[34]Liu H B,Lu Y Z,Lan B L,et al. Codon usage by chloroplast gene is bias in Hemiptelea davidii[J]. Journal of Genetics,2020,99(1):1-11.
[35]Kawabe A,Miyashita N T. Patterns of codon usage bias in three dicot and four monocot plant species[J]. Genes & Genetic Systems,2003,78(5):343-352.
[36]Hershberg R,Petrov D A. Selection on codon bias[J]. Annual Review of Genetics,2008,42:287-299.
收稿日期:2022-12-29
基金项目:江苏省高等学校基础科学(自然科学)研究面上项目(编号:21KJB210006);江苏省种业振兴“揭榜挂帅”项目[编号:JBGS(2021)091];江苏省林业科技创新与推广项目[编号:LYKJ(2021)02];苏州农业职业技术学院博士提升计划科研启动基金项目(编号:BS2103)。
作者简介:程培蕾(1990—),女,山东蒙阴人,博士,讲师,从事观赏植物遗传育种与分子生物学研究。E-mail:plcheng@szai.edu.cn。
猜你喜欢
文物鉴定与鉴赏(2018年2期)2018-03-07 20:09:09
湖北农业科学(2018年2期)2018-03-06 00:32:21
现代园艺·综合版(2017年8期)2018-01-16 21:42:52
江苏农业科学(2017年10期)2017-07-21 16:35:32
农业研究与应用(2017年2期)2017-07-01 06:16:51
商业经济(2017年6期)2017-06-15 22:03:46
现代农业研究(2017年1期)2017-03-20 18:53:56
中国科技博览(2016年5期)2016-04-23 10:56:35
安徽农学通报(2015年20期)2015-11-07 00:52:19
江苏农业科学(2014年9期)2014-11-15 05:08:43
