
云南省怒江州独龙牛内阿米巴原虫的感染情况调查
2023-10-24 13:33庄尔俊陈远腾尹雪宇李海龙
动物医学进展 2023年10期
庄尔俊,陈远腾,尹雪宇,李海龙,2*
(1.大理大学基础医学院,云南大理 671000;2.云南省滇西抗病原植物资源筛选研究重点实验室,云南大理 671000)
内阿米巴原虫(Entamoebaspp.)是一种能够感染人、家畜和野生动物的单细胞原虫,在全世界范围均有流行[1-2]。大多数内阿米巴原虫主要分为包囊和滋养体两个阶段,其在宿主之间的传播主要通过粪-口途径来实现,感染后主要表现为肠道疾病以及宿主肝脏、大脑和其他器官的损伤[3-4]。阿米巴病是常见的寄生虫病之一,主要是由溶组织内阿米巴引起。我国西南地区成为人感染阿米巴病的高发地区[5-7]。在我国西藏、青海、甘肃、云南、江苏、湖南、江西、福建等地均已有报道[4,8-9]。
反刍动物常可作为内阿米巴原虫的宿主,独龙牛是云南省怒江州的珍贵肉用牛种,内阿米巴原虫作为一种人畜共患寄生虫,可能对当地独龙牛养殖业造成损失,与独龙牛密切接触的牧民可能存在感染的风险。本研究以怒江州独龙牛为调查对象,调查并分析内阿米巴原虫的感染情况和主要虫种,旨在为当地内阿米巴原虫的防控提供理论参考,也为云南省内阿米巴原虫进一步的研究奠定基础。
1 材料与方法
1.1 材料
1.1.1 样本 2017年9月至2018年9月,从云南省怒江州怒江流域的鸠门当村、亚左洛村、古泉村、茨开镇及独龙江流域的独龙江乡5个采样点,在春、夏、秋、冬四个季节采集到625份新鲜独龙牛粪便样本,其中怒江流域557份,独龙江流域68份,-20 ℃保存。内阿米巴原虫阳性DNA样本由大理大学病原生物学寄生虫实验室保存。
1.1.2 主要试剂 10×PCR Buffer(AM21651A),MgCl2(AM21652A),dNTPs Mixture(AM21653A),Ex-TaqDNA聚合酶(AM21653A),DL-2 000 Marker(AM11312A),6×DNA Loading Buffer(AGY0007A),宝日医生物技术(北京)有限公司产品;DNA提取试剂盒(D4015020000K24U001),美国OMEGA Bio-Tek公司产品;4S Green核酸染色剂(A616694),生工生物工程(上海)有限公司产品。
1.1.3 主要仪器 冷冻离心机(5427R),艾本德中国有限公司产品;PCR仪(Applied Biosystems 2720),美国Bio-Rad公司产品;电泳仪(DYY-12)、琼脂糖水平电泳槽(DYCP-31DN),北京六一生物科技有限公司产品;全自动凝胶成像系统(Syngene GeneGenius),美国Syngene公司产品。
1.2 方法
1.2.1 粪便DNA提取 独龙牛粪便样本DNA的提取按照粪便DNA提取试剂盒所述步骤进行操作,提取的DNA放置于-20 ℃保存。
1.2.2 引物设计与PCR扩增 扩增内阿米巴原虫18S rRNA基因位点,引物序列参照VERWEIJ J J等的报道[10]进行设计(表1),由生工生物工程(上海)股份有限公司进行合成。预扩增产物大小约为600 bp。

表1 扩增内阿米巴原虫18S rRNA基因位点的引物序列、目的片段、退火温度Table 1 Primer sequences,fragment length,annealing temperature for amplification of the 18S rRNA gene of Entamoeba spp.
PCR扩增反应体系:10×PCR buffer 2.5 μL,MgCl21.5 μL,dNTPs Mixture 2 μL,Ex-TaqDNA聚合酶0.25 μL,引物Entam1、Entam2均为0.5 μL,模板DNA 1 μL,加ddH2O补至25 μL。
1.2.3 PCR产物检测 配置10 g/L琼脂糖凝胶,加入4S Green核酸染色剂混匀,取PCR产物5 μL与1 μL的6×DNA Loading buffer混合上样,电泳的电压为120 V,时间约为30 min。PCR阳性样本送生工生物工程(上海)股份有限公司进行测序。
1.2.4 遗传进化分析 使用BLAST将测序获得的样本序列在GenBank上与内阿米巴原虫参考序列进行相似性比对。参考序列主要包括比利时的猴源Entamoebasp.MG107/BEL(登录号:GU437825)、中国的牛源Entamoebasp.MG107/BEL(登录号:MT734309)、中国的牛源E.bovis(登录号:MT734187)、冰岛的鹿源E.bovis(登录号:FN666252)、瑞典的羊源E.bovis(登录号:FN666250)、瑞典的牛源E.bovis(登录号:FN666249)、瑞典的鹿源Entamoebasp.RL1(登录号:FN666253)、瑞典的牛源Entamoebasp.RL2(登录号:FN686363)、瑞典的龟源Entamoebasp.RL5(登录号:FR686365)、德国的猴源Entamoebasp.RL3(登录号:FN686358)、英国的牛源Entamoebasp.RL8(登录号:KR025406)和利比亚的牛源Entamoebasp.RL4(登录号:FR686361)。以Entamoebasp.RL5(登录号:FR686365)作为外群,使用MEGA-11计算序列之间的遗传距离,采用邻接法(NJ)构建系统进化树。
1.2.5 数据的统计分析 使用SPSS 18.0软件对数据进行统计分析,采用卡方检验分析内阿米巴原虫感染率的差异,当P<0.05时,差异显著;当P>0.05时,差异不显著;当P<0.01时,差异极显著。
2 结果
2.1 常规PCR扩增结果
利用常规PCR方法对内阿米巴原虫18S rRNA基因位点进行扩增,625份样本中共有61份样本扩增出阳性条带,条带大小约为600 bp(图1)。

M.DNA标准DL 2 000;1~4.阳性样品;5.阳性对照;6.阴性对照M.DNA Marker DL 2 000; 1-4.Positive samples; 5.Positive control; 6.Negative control图1 内阿米巴原虫18S rRNA基因PCR扩增结果Fig.1 PCR amplification results of 18S rRNA gene of Entamoeba spp.
2.2 测序与分子鉴定结果
对本研究获得的61份内阿米巴原虫阳性样本测序结果进行比对分析,其中51份鉴定为E.bovis,与瑞典的牛源E.bovis(FN666249)序列相似性为95.19%~99.26%;10份鉴定为Entamoebasp.MG107/BEL,与比利时的猴源Entamoebasp.MG107/BEL(GU437825)序列相似性为95.45%~98.83%,两者均不属于人畜共患寄生虫。E.bovis在5个采样点的独龙牛中均有感染,是当地独龙牛感染内阿米巴原虫的优势虫种,所占比例为83.61%(51/61),以茨开镇独龙牛的E.bovis感染数量最多。独龙江流域未检测到Entamoebasp.MG107/BEL,怒江流域范围内除鸠门当村,其余采样地点均检测到Entamoebasp.MG107/BEL,所占比例为16.39%(10/61),以茨开镇独龙牛的Entamoebasp.MG107/BEL感染数量最多(表2)。将阳性序列上传至GenBank获得登录号为:OP563753~OP563813,基于内阿米巴原虫18S rRNA构建系统进化树,本研究中的E.bovis序列OP563782与冰岛的鹿源E.bovis(FN666252)位于同一进化分支,E.bovis序列OP563771与瑞典的羊源E.bovis(FN666250)位于同一进化分支,E.bovis序列OP563778与瑞典的牛源E.bovis(FN666249)位于同一进化分支;Entamoebasp.MG107/BEL序列OP563811和OP563805与比利时的猴源Entamoebasp.MG107/BEL(GU437825)位于同一进化分支(图2)。
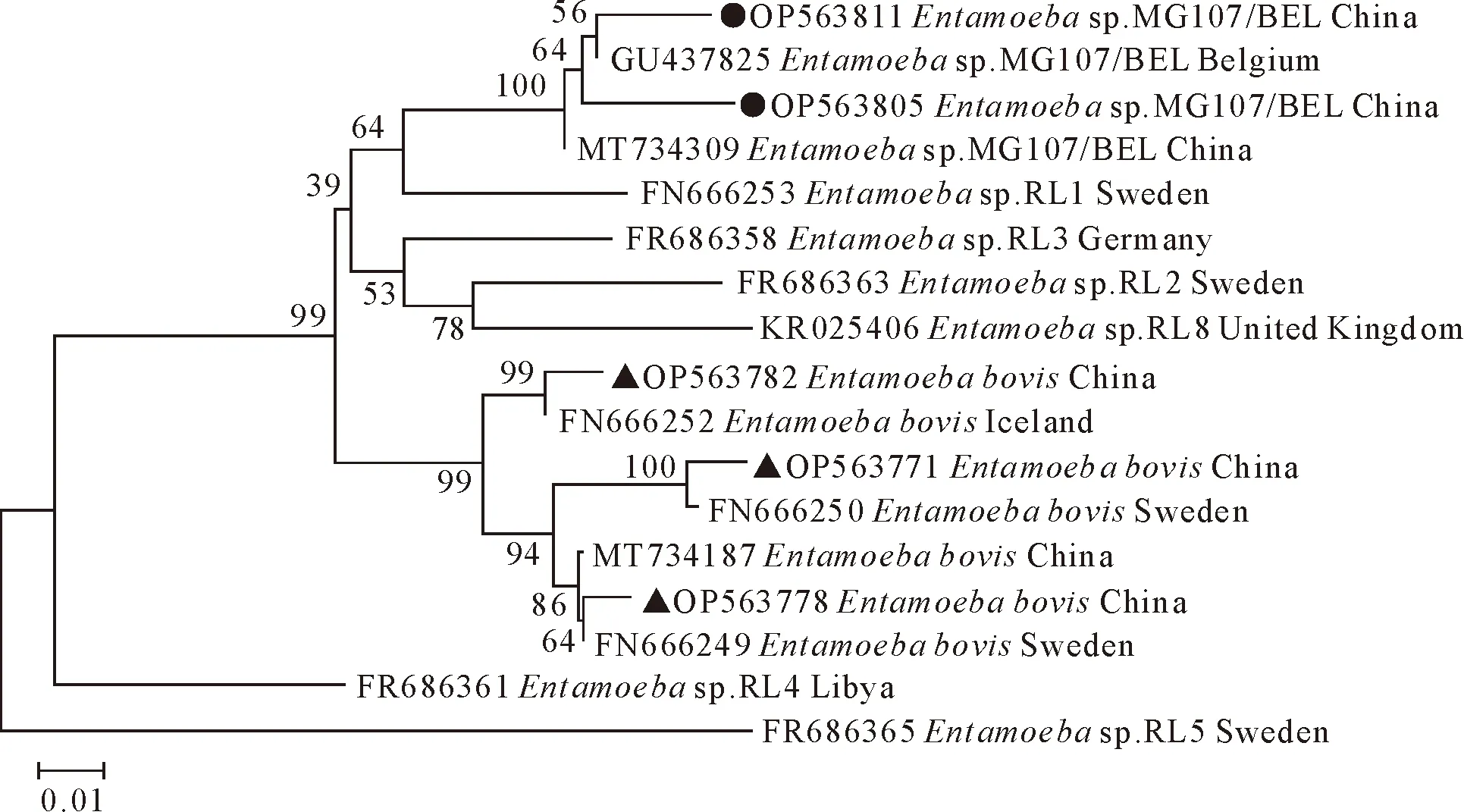
▲为本研究中鉴定的部分Entamoeba bovis;●为本研究中鉴定的部分Entamoeba sp.MG107/BEL▲Part of Entamoeba bovis identified in this study;●part of Entamoeba sp.MG107/BEL identified in this study图2 内阿米巴原虫系统进化树Fig.2 Phylogenetic tree of Entamoeba spp.
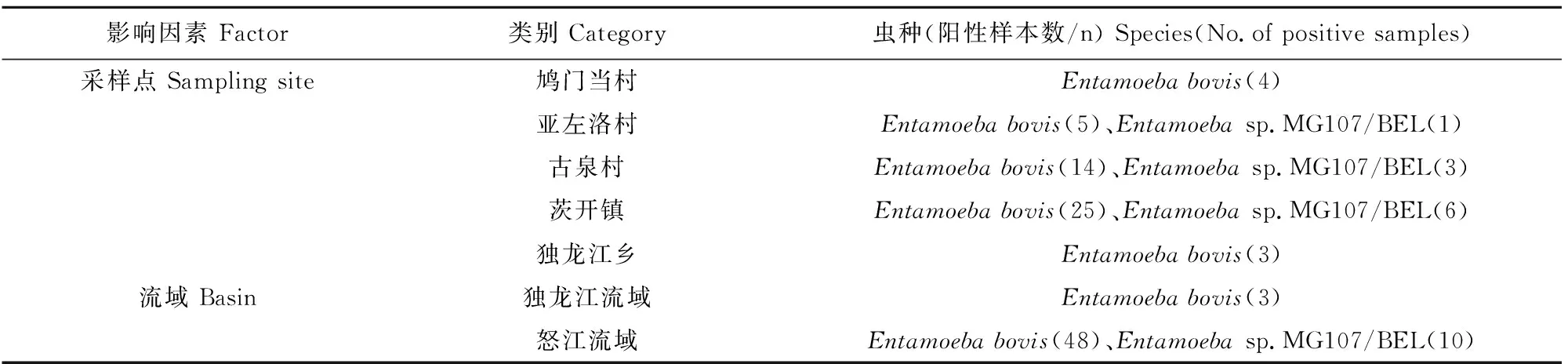
表2 云南省怒江州独龙牛内阿米巴原虫分布情况Table 2 Distribution of Entamoeba spp.in Bos frontalis in Nujiang prefecture,Yunnan province
2.3 独龙牛内阿米巴原虫感染情况
625份独龙牛粪便样本中共检测出61份内阿米巴原虫阳性样本,总感染率为9.76%。5个采样点中,鸠门当村、亚左洛村、古泉村、茨开镇、独龙江乡独龙牛的内阿米巴原虫感染率分别为4.12%(4/97)、7.59%(6/79)、11.18%(17/152)、13.54%(31/229)和4.41%(3/68),不同地点之间,其感染率差异显著(P<0.05)。怒江流域与独龙江流域独龙牛的内阿米巴原虫感染率分别为10.41%(58/557)和4.41%(3/68),两流域之间,其感染率差异不显著(P>0.05)(表3)。春、夏、秋、冬4个季节中,独龙牛内阿米巴原虫的感染率分别为11.93%(26/218)、23.66%(22/93),3.33%(4/120)和4.64%(9/194),不同季节之间,其感染率差异极显著(P<0.01)(表4)。

表3 不同采样点和流域内阿米巴原虫的感染情况Table 3 The infections of Entamoeba spp.in different sampling sites and basins
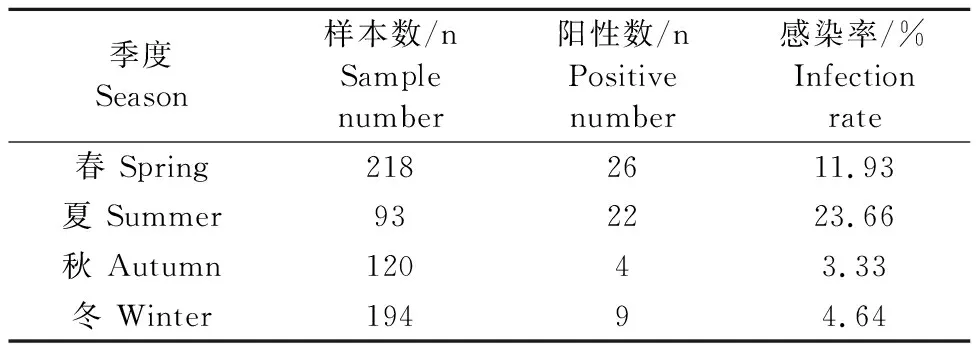
表4 不同季度内阿米巴原虫的感染情况Table 4 The infections of Entamoeba spp.in different seasons
3 讨论
独龙牛是印度野牛的亚种,除了分布在我国云南省怒江州,还分布在孟加拉国、印度、缅甸和不丹[11]。近年来印度和不丹已报道的研究显示,独龙牛感染的寄生虫包括指形长刺线虫、牛弓首蛔虫、弓形虫和绦虫[12-14],国内已报道的研究显示,独龙牛感染了环孢子虫、毕氏肠微孢子虫和芽囊原虫等[15-17]。国内外尚未有独龙牛内阿米巴原虫的相关调查研究,故本研究对云南省怒江州独龙牛内阿米巴原虫的感染情况进行调查,结果显示内阿米巴原虫的感染率为9.76%,茨开镇的感染率最高,鸠门当村的感染率最低,不同地点之间差异显著(P<0.05),分析其原因可能与各个地点独龙牛的养殖模式有关,集体管理模式可能导致内阿米巴原虫更加容易传播,而将独龙牛散养在深山中,牛群活动范围更大,聚集的规模更小,不容易互相传播,怒江流域的茨开镇和古泉村应加强对其内阿米巴原虫的防治。国外的研究报道显示,日本奶牛内阿米巴原虫的感染率为72%,但该研究牛粪便样本数为25份[3];乌干达牛内阿米巴原虫的感染率为80%,采集样本数为45份[18],国外感染率比本试验感染率高的原因可能是样本采集数量、地理位置、气候条件不同造成的。国内青海省牦牛内阿米巴原虫的感染率为36.32%[19],云南大理奶牛内阿米巴原虫的感染率为30.09%[20],河北和天津地区奶牛内阿米巴原虫的感染率为33.27%[21],均高于云南省独龙牛的感染率,其差异可能与样本采集数量、地理位置、生态环境、品种、自身免疫状况等因素有关。本调查结果还显示,4个季节中独龙牛内阿米巴原虫的感染率在夏季最高,春季和冬季次之,秋季最低,差异显著(P<0.05)。青海省牦牛内阿米巴原虫的感染率在夏季也是最高的[19],夏季可能是牛感染内阿米巴原虫的高危季节,夏季应特别加强对内阿米巴原虫的检测。
本研究基于内阿米巴原虫18S rRNA基因,鉴定出E.bovis和Entamoebasp.MG107/BEL两个种的内阿米巴原虫,均不属于人畜共患寄生虫种类,其中,E.bovis是独龙牛感染的优势虫种。国外的研究显示,Stensvold C R等建立了一种新的内阿米巴原虫命名方式——核糖体谱系(RL)[22]。反刍动物可感染的内阿米巴原虫主要有E.bovis、Entamoebasp.RL1、Entamoebasp.RL2、Entamoebasp.RL4(灵长类动物的Entamoebasp.RL3)和Entamoebasp.RL8[3,23]。本研究结果中未鉴定出核糖体谱系的内阿米巴原虫,Entamoebasp.MG107/BEL目前可能被认为是内阿米巴原虫的暂定型,并未正式归为某一个谱系之中,通过进化树分析推测其与E.bovis具有较高的同源性。Entamoebasp.MG107/BEL的发现也是继衡昭君和Ren M等之后再一次从牛中鉴定出该内阿米巴原虫[19-20]。
独龙牛作为怒江州独龙族牧民长期驯养的一种半野生畜种,具有很好的经济前景,但因分布于偏远山区,增加了防治其人畜共患寄生虫工作的难度。E.bovis在牛中据有普遍传染性,但其致病性很低[3],Matsubayashi M等研究发现,波列基内阿米巴(Entamoebapolecki)在猪体内具有较低的致病性,当其与胞内劳森菌混合感染时能够诱导引发回肠炎[24]。当牛内阿米巴原虫和其他病原体混合感染时,是否会加重疾病严重程度尚未可知,还需进一步的研究来证实。当地牧民应改善养殖环境的卫生条件,做好牛粪便的无害化处理,建议当地有关部门加强对独龙牛内阿米巴原虫感染的检测检疫,加强对内阿米巴原虫病的防治宣传教育。
猜你喜欢
中国动物保健(2022年2期)2022-05-05
晋中学院学报(2022年2期)2022-02-03
畜牧与兽医(2021年10期)2021-10-09
种子(2021年2期)2021-03-31
矿产勘查(2020年8期)2020-12-25
民族翻译(2020年4期)2020-02-27
浙江化工(2019年5期)2019-06-04
山西农经(2019年21期)2019-01-02
动物医学进展(2018年11期)2018-12-05
现代畜牧兽医(2018年7期)2018-08-25
