
多浪羊CB1基因表达分析与脂肪沉积的关联性研究
2023-10-19 08:33张永浩郭丽君闫伟王玉涛
畜牧与兽医 2023年4期
张永浩,郭丽君,闫伟,王玉涛*
(1. 喀什大学生命与地理科学学院,新疆 喀什 844000;2. 江苏农牧科技职业学院动物科技学院,江苏 泰州 225300)
内源性大麻素系统(endogenous cannabinod system,ECS)是一个多功能的信号系统,由酰胺、酯类和长链不饱和酸组成的脂质分子[1]。直接或间接参与多种生理过程,由内源性大麻素、大麻素受体及与之相关的合成、水解酶共同构成[2]。大麻素受体主要包括CB1和CB2,且两者均属于具有7次跨膜结构的G蛋白偶联受体家族。CB1分布广泛,在大脑皮层、下丘脑、肝脏、脂肪组织、骨骼肌、胃肠道均有分布[3],为7次跨膜G蛋白偶联受体,能耦联并活化Gi蛋白,抑制腺苷环化酶活性,进而降低cAMP水平;还能抑制钙离子通道、活化钾离子通道及丝裂原活化蛋白激酶[4]。CB2基因主要表达于外周免疫器官,参与免疫调节过程。CB1在结构上一个重要特征是:激动剂与CB1结合比拮抗剂与CB1结合体积会发生变化,这种可塑性使CB1能够对各种不同的配体做出反应,这与CB1调节不同生理活动或心理状态的功能是一致的[5]。CB1基因的高表达是对慢性循环的内源性大麻素的反应[7],且CB1基因在治疗肥胖[8],减缓药物成瘾[9],调节抑郁[10],缓解帕金森综合征[11],乃至化妆品研发[12]和生殖方面[13]均有一定效果。
如今在羊肉产量满足人们需求的同时,人们对羊肉口感的要求越来越高。肌内脂肪含量(IMF)是评价羊肉性状的一个重要指标,直接影响羊肉的风味、嫩度、适口性、多汁性[14]。此外,因羊肉价格远远高于猪肉和鸡肉[15],这间接影响着人们需求,直接和间接因素均会影响羊肉商品特性和经济效益。多浪羊是新疆的一个优良品种,是由新疆喀什市麦盖提县当地长期培育而成的地方品种,具有生长迅速,肉质鲜美的特点,深受南北疆牧民的喜爱,又称“麦盖提羊”。动物脂肪不仅作为人体重要的能源物质,而且为人类供给了许多必需脂肪酸,协助脂溶性维生素的吸收。作为肉食品来讲,脂肪组分和加工原材料均很重要,因此调节脂肪含量成为了研究热点问题[16-17]。为了满足消费者对高质量羊肉制品的需求及挖掘其巨大的经济潜力,研究绵羊脂肪沉积的生物学机制已成为近年来研究热点。本研究旨在分析绵羊CB1基因对肌内脂肪沉积的影响,为进一步揭示动物肉质调控的生物学机制提供理论基础和技术支持,具有重要的理论意义和应用价值。
1 材料与方法
1.1 试验动物及样品
样品取自喀什德吾苒屠宰场。采集不同月龄(3、6、12月龄)多浪羊各3只,每只采其背最长肌、皮下脂肪、腹部脂肪、肠系膜组织用于索氏抽提、脂肪代谢酶活测定、石蜡切片、RNA和DNA提取。
1.2 主要试剂
TRIzol提取试剂盒、反转录试剂盒、荧光定量试剂盒、2×TaqPCR Mix均购自中科瑞泰(北京)生物科技有限公司;A067总脂酶(脂蛋白脂肪酶/肝脂酶、LPL/HL)测试盒、绵羊激素敏感性甘油三酯脂肪酶(HSL)酶联免疫检测试剂盒均购自南京建成生物工程研究所。
1.3 荧光定量PCR(RT-qPCR)
按照TRIzol试剂使用说明书方法进行操作。对不同月龄多浪羊胴体不同部位组织(背最长肌、皮下脂肪、腹部脂肪、肠系膜)的总RNA进行提取。提取方法为:取组织进行液氮研磨至粉状,加入TRIzol摇匀,冰上静置,弃去上层油脂,加入氯仿离心取上清液,加入预冷的异丙醇,离心使总RNA沉淀,并用75%酒精洗涤。使用琼脂糖检测和紫外分光光度计对符合要求的总RNA进行反转录[18]及荧光定量分析,具体操作步骤为:95 ℃初始变30 s;95 ℃变性50 s,60 ℃退火延伸60 s,共计40个循环,熔解曲线遵循7500 Real Time PCR System机器自带程序绘制。
1.4 CB1扩增
由改进过的酚氯仿法[19]提取总DNA,并参考刘贤勋[30]设计引物,扩增片段1 400 bp左右的CB1编码区,引物由金唯智生物技术有限公司合成,PCR扩增后经琼脂糖凝胶电泳检测,产物送至艾康健武汉生物技术有限公司测序。

表1 CB1基因引物信息
1.5 脂肪测定
参照食品安全国家标准GB 5009.6—2016《食品中脂肪的测定》索氏抽提法进行操作。
1.6 脂肪代谢相关酶活测定
用比色法测定LPL和HL酶活性。用酶联免疫吸附试验测定HSL酶活性。
1.7 脂肪细胞面积测定
将采回的样品立即放入4%中性甲醛溶液中浸泡10 h,将固定好的样品切成2.5 cm×2.5 cm×1 cm的样品在自来水下洗涤固定液10 h,用梯度酒精(70%、80%、90%、95%、100%)各40 min脱水;酒精二甲苯、二甲苯各透明30 min;在63 ℃条件下二甲苯石蜡、石蜡各浸蜡2 h;包埋切片,采用HE染色法染色,置于10×40倍镜下进行切片图像采集,将每个组织做2个载玻片的切片,每张切片随机选4个视野,拍照导入Image Pro Plus 6.0图像分析系统中,每个视野均匀选取9个细胞,共计72个细胞,进行平均面积的计算。在Image Pro Plus 6.0中进行空间校正并设置好标尺,用自动绘图法并人工校正,对随机选取的9个细胞进行面积的测量。
1.8 相关性分析
SPSS 22.0、Image Pro Plus 6.0、MEGA等软件对细胞和测序数据进行整理分析,对各处理间的差异采用LSD法和邓肯法进行0.05水平上的显著性检验;并进行不同月龄差异性分析。
2 结果与分析
2.1 CB1基因分析
2.1.1 CB1基因表达量分析
提取不同月龄绵羊背最长肌、皮下脂肪、腹部脂肪、肠系膜组织的总RNA,对其完整性、纯度进行检测。通过绵羊CB1、GAPDH基因溶解曲线图可知,得到的样本mRNA表达量准确性高(图略)。经过2-ΔΔCt法测得CB1基因mRNA相对表达量,测定结果如图1所示。CB1基因表达量随着年龄的增长而逐步提高,且均表现为在6~12月龄的增长速率高于3~6月龄的增长速率。背最长肌3~6月龄CB1表达量增长了128.07%,6~12月龄CB1表达量增长了107.69%;腹部脂肪在3~6月龄CB1表达量增长了160%,在6~12月龄增长了143.65%;皮下脂肪3~6月龄CB1表达量增长了352.9%,6~12月龄CB1表达量增长了88.3%;肠系膜组织3~6月龄CB1表达量增长了135.69%,6~12月龄CB1表达量增长了74.29%。

图1 CB1基因mRNA相对表达量
2.1.2 CB1基因CDS序列分析
将不同月龄的多浪羊CB1基因进行PCR扩增并测序。用MEGA软件对多浪羊CB1基因与绵羊(XM 004011281.4)、山羊(XM 018053156.1)、大羚羊(XM 040238815.1)、加拿大盘羊(CP 011893.1)、野牦牛(XM 005889296.1)的CB1基因构建NJ进化树。由图2可知,多浪羊与绵羊、加拿大盘羊聚为一支,与山羊的关系比大羚羊、野牦牛的近。

注:1A~1C为3只12月龄多浪羊,2A~2C为3只6月龄多浪羊,3A~3C为3只3月龄多浪羊
用DNASP软件分析结果见表2。共发现了6个单倍型,4个变异位点,核苷酸差异系数(Pi)为0.001 16,单倍型多样性(Hd)为0.778,核苷酸平均差异度(K)为1.4,同义替换率(ds)为0.003 24,非同义替换率(dn)为0.000 39,dn/ds=0.12,可知基因进化比较平缓稳定,中性测试不显著(P>0.1),因此,根据中性进化模型受环境人为影响较少。A+T平均比例为42.87%,G+C平均比例为57.13%。G+C含量高于A+T含量,说明存在1定的碱基偏倚性。仅存在一个氨基酸替换,且经过在线软件分析(http://genetics.bwh.harvard.edu/pph2/)得出氨基酸替换对蛋白质影响概率为0.188,可见氨基酸对蛋白质的影响较为温和(表3、图3)。
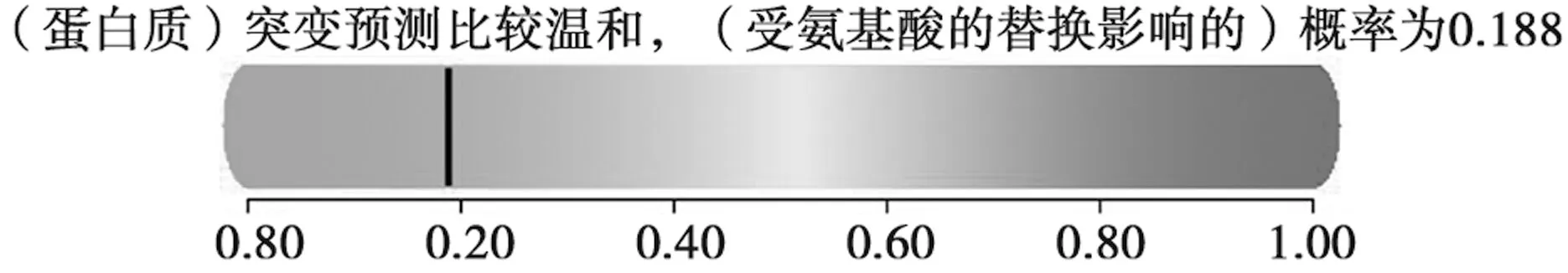
图3 CB1基因编码区SNP预测

表2 CB1基因编码区变异位点

表3 CB1氨基酸变异位点
由图4可知,H_1(与NCBI中XM_004011281.4序列一致)为优势单倍型。

图4 多浪羊CB1基因单倍型network网络图
2.2 脂肪含量测定
经索氏抽提测得脂肪含量,测定结果见图5。背部脂肪含量显著低于其他部位的脂肪含量,且不同部位皆随着月龄的增加,上升趋势明显。背最长肌3~6月龄提高了103.45%,6~12月龄提高了84.26%。腹部脂肪3~6月龄提高了314.47%,6~12月龄提升了1.21%。皮下脂肪3~6月龄提高了60.4%,6~12月龄提高了-2.19%。肠系膜组织3~6月龄提高了23.23%,6~12月龄提高了3.84%。由图5可知,在1年内背最长肌一直以比较均匀的速度增长;皮下脂肪、腹部脂肪、肠系膜组织的脂肪在3~6月份增长速度高于6~12月份的增长速度。

图5 脂肪含量变化
2.3 脂肪代谢酶活测定
本试验采取比色法、酶联免疫吸附法对多浪羊背最长肌、皮下脂肪、腹部脂肪、肠系膜组织的酶活性进行测定,测定结果见图6。用比色法对肝脂酶酶活性进行测定,测定背最长肌在3~6月龄肝脂酶的活性增长了289.38%,在6~12月龄肝脂酶的活性增长了-32.697%;腹部脂肪在3~6月龄肝脂酶的活性增长了501.523%,在6~12月龄肝脂酶的活性增长了52.15%;皮下脂肪在3~6月龄肝脂酶的活性增长了120.56%,在6~12月龄肝脂酶的活性增长了-8.22%;肠系膜组织在3~6月龄肝脂酶的活性增长了230.23%,在6~12月龄肝脂酶的活性增长了21.78%。
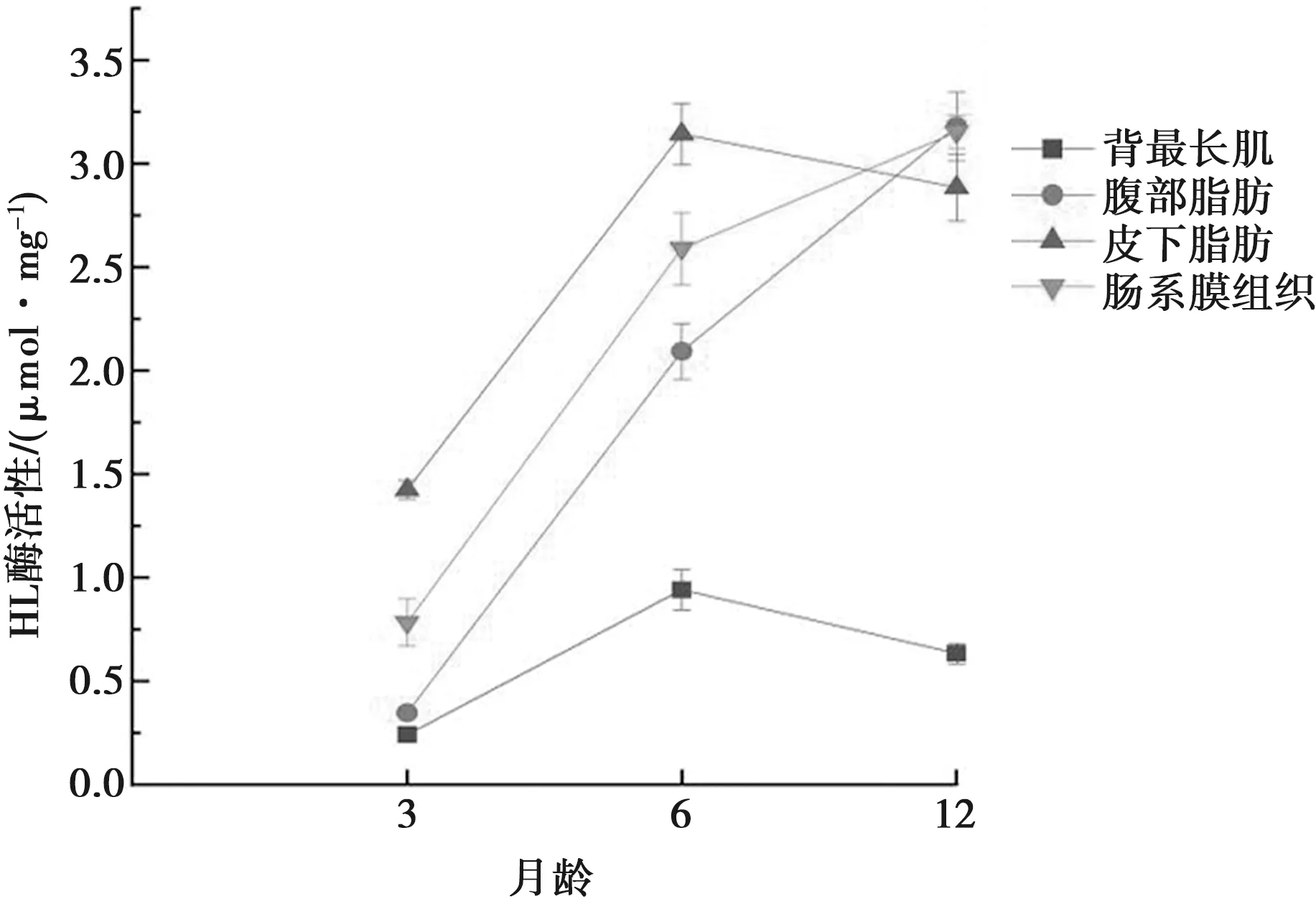
图6 肝脂酶活性变化
不同月龄HSL活性变化见图7。皮下脂肪、腹部脂肪、肠系膜组织中的HSC均在6月龄达到最高,背最长肌在6月龄达到最低。背最长肌在3~6月龄HSL酶活性增长了-11.84%,在6~12月龄HSL酶活性增长了29.1%;腹部脂肪在3~6月龄HSL酶活性增长了132.52%,在6~12月龄HSL酶活性增长了-12.87%;皮下脂肪在3~6月龄HSL酶活性增长了254.3%,在6~12月龄HSL酶活性增长了-18.99%;肠系膜组织在3~6月龄HSL酶活性增长了6.868%,6~12月龄HSL酶活性增长了-36.8%。

图7 激素敏感性甘油三酯脂肪酶含量变化
不同月龄LPL酶活性变化见图8。脂蛋白脂肪酶在整个生长阶段,酶活随着月龄的增加而提高,其中皮下脂肪、腹部脂肪、肠系膜组织中酶活在6~12月龄的增长速率高于3~6月龄的增长速率;背最长肌中酶活在3~6月龄的增长速率高于6~12月龄的增长速率。背最长肌在3~6月龄脂蛋白脂肪酶活性增长了127.69%,在6~12月龄脂蛋白脂肪酶活性增长了31.07%;腹部脂肪在3~6月龄脂蛋白脂肪酶活性增长了140.23%,在6~12月龄脂蛋白脂肪酶活性增长了143.9%。皮下脂肪在3~6月龄脂蛋白脂肪酶活性增长了-23.02%,在6~12月龄脂蛋白脂肪酶活性增长了330.1%。肠系膜组织在3~6月龄脂蛋白脂肪酶活性增长了13.94%,在6~12月龄脂蛋白脂肪酶活性增长了72.76%。

图8 脂蛋白脂肪酶活性变化
2.4 组织学观察
采取石蜡切片法对多浪羊背最长肌、皮下脂肪、腹部脂肪、肠系膜组织的脂肪细胞大小进行测定。由图9、图10可知,不同月龄4个部位的脂肪组织进行HE染色后,各个部位组织切片上几乎都能观察到大小均一的脂肪细胞,脂肪细胞饱满,细胞形态呈圆形、椭圆形或多边形,胞浆中存在大型脂滴,细胞核和细胞器被压迫到边缘、形态完整无破损。

图9 不同月龄背最长肌、腹部脂肪形态(400×)

图10 不同月龄皮下脂肪、肠系膜组织形态(400×)
由图11可见,在整个生长阶段背最长肌、皮下脂肪、腹部脂肪、肠系膜组织中脂肪细胞大小随着月龄的增加而逐步变大,且均表现为在3~6月龄的增长速率高于6~12月龄的增长速率。背最长肌在3~6月龄脂肪细胞面积增长了158.06%,在6~12月龄脂肪细胞面积增长了22.4%;腹部脂肪在3~6月龄脂肪细胞面积增长了260.6%,在6~12月龄脂肪细胞面积增长了33.87%;皮下脂肪在3~6月龄脂肪细胞面积增长了128.7%,在6~12月龄脂肪细胞面积增长了19.28%;肠系膜组织在3~6月龄脂肪细胞面积增长了136.8%,在6~12月龄脂肪细胞增长了50.43%。

图11 细胞面积变化
2.5 CB1基因与性状的关联性分析
如表4所示,CB1基因表达量与各组织的脂肪含量呈显著正相关(背最长肌相关系数为0.989,P<0.01;皮下脂肪相关系数为0.813,P<0.01;腹部脂肪相关系数为0.736,P<0.05;肠系膜组织相关系数为0.889,P<0.01)。
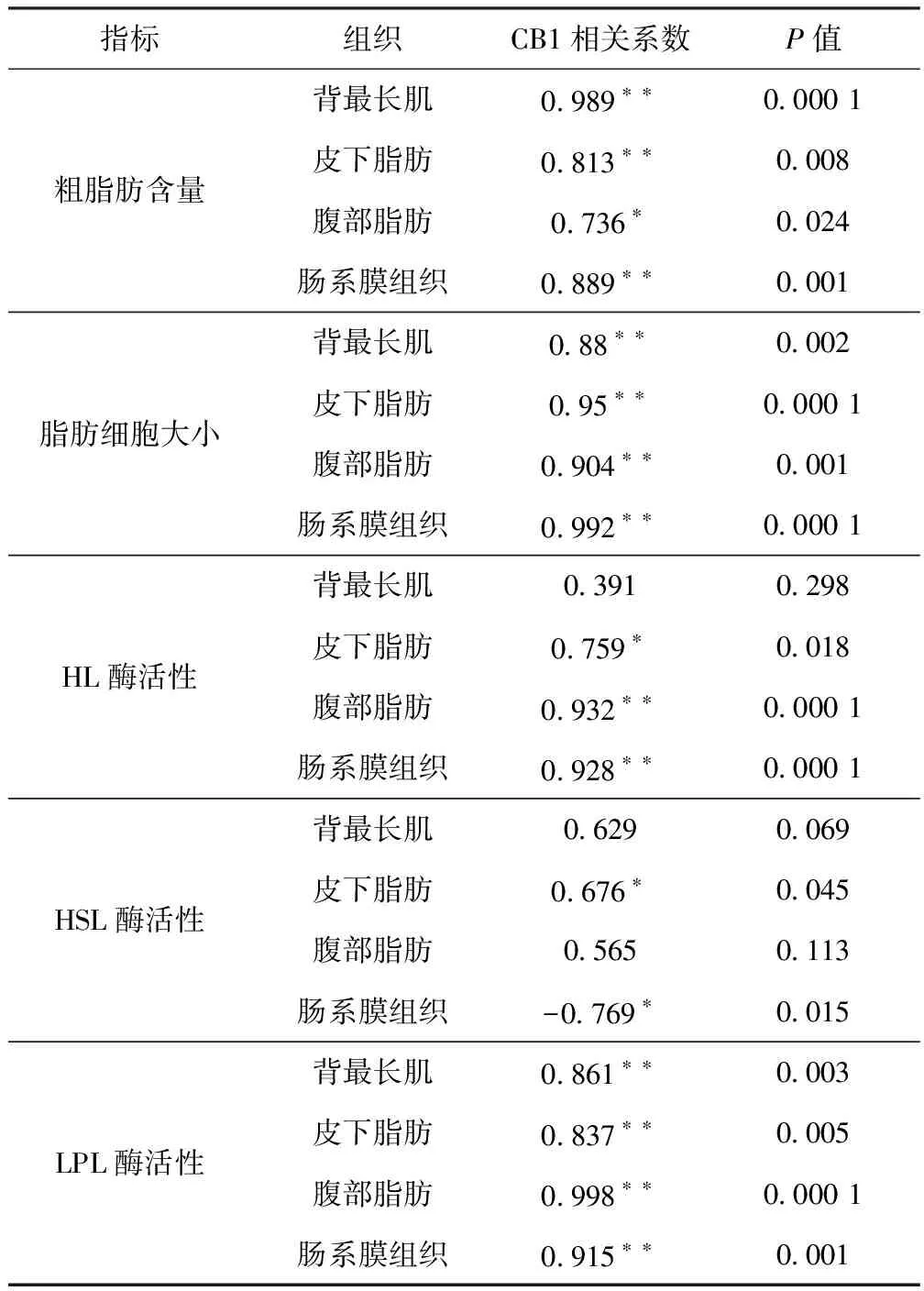
表4 不同日龄CB1基因相对表达量与各项指标的相关性分析
CB1基因表达量与各组织中脂肪细胞大小呈显著正相关(背最长肌相关系数为0.88,P<0.01;皮下脂肪相关系数为0.95,P<0.01;腹部脂肪相关系数为0.904,P<0.01;肠系膜组织相关系数为0.992,P<0.01)。
CB1基因表达量与HL酶活性相关性在不同部位呈现出不同的结果,在皮下脂肪,腹部脂肪,肠系膜组织中呈显著正相关(皮下脂肪相关系数为0.759,P<0.05;腹部脂肪相关系数为0.932,P<0.01;肠系膜组织相关系数为0.928,P<0.01),在背最长肌中无显著相关性。
CB1基因与HSL酶活性相关性在不同部位呈现出不同的结果,在皮下脂肪和肠系膜组织中呈显著正相关(皮下脂肪相关系数为0.676,P<0.05;肠系膜组织相关系数为-0.769,P<0.05)在背最长肌和腹部脂肪中无显著相关性。
CB1基因表达量与各组织中LPL酶活性呈显著正相关(背最长肌相关系数为0.861,P<0.01;皮下脂肪相关系数为0.837,P<0.01;腹部脂肪相关系数为0.998,P<0.01;肠系膜组织相关系数为0.915,P<0.01)。
2.6 月龄与脂肪沉积相关指标的差异性分析
如表5所示,背最长肌粗脂肪含量与年龄呈现出显著的差异性,皮下脂肪、腹部脂肪和肠系膜组织粗脂肪含量与年龄相关性差异性不显著。
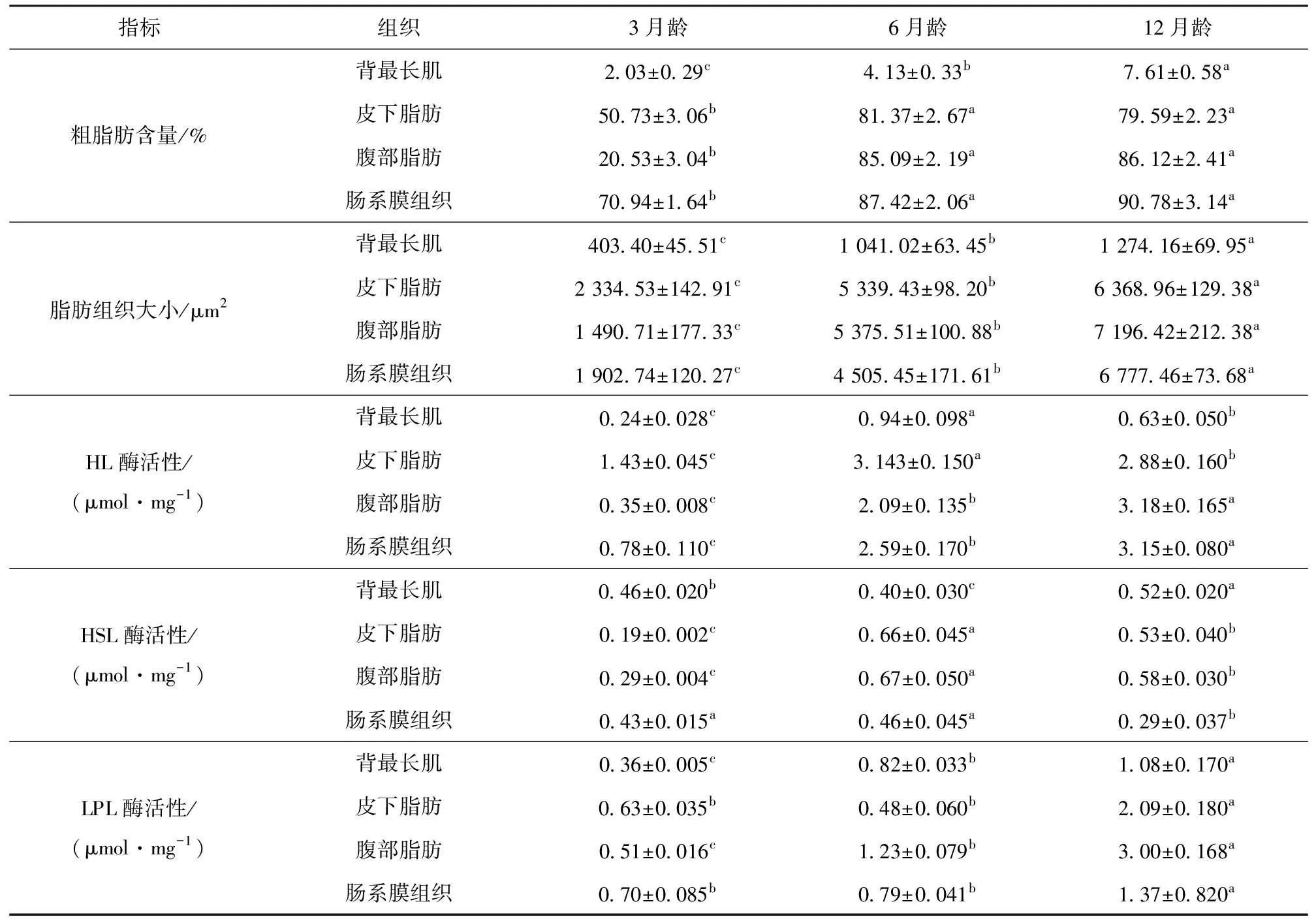
表5 不同月龄各项指标的差异性分析
不同部位脂肪组织大小皆与年龄呈现出显著的差异性(P<0.01)。不同部位CB1表达量皆与年龄呈现出显著的差异性(P<0.05),HL酶活性皆与年龄呈现出显著的差异性(P<0.05)。
背最长肌、皮下脂肪和腹部脂肪部位HSL酶活性与年龄呈现出显著性差异(P<0.05),对于肠系膜组织中HSL酶活性3月龄与6月龄差异性不显著(P>0.05),二者与12月龄酶活性呈现出显著性差异(P<0.05)。
背最长肌和腹部脂肪中LPL酶活性与年龄呈现出显著性差异(P<0.05),皮下脂肪和肠系膜组织中LPL酶活性3月龄与6月龄差异性不显著(P>0.05),但二者与12月龄酶活性呈现出显著性差异(P<0.05)。
3 讨论
魏宗友等[20]研究发现,随着日粮的增加,腹脂率会随HL活性增强呈现出不同程度的提高;涂玮等[21]在罗非鱼上也发现有这种趋势,随着饲料水平的升高,罗非鱼的HL活性呈现出先升高后下降的趋势;HL酶与LPL酶在功能上相似主要在中密度脂蛋白和高密度脂蛋白代谢中起作用是重要的参与者,但不是唯一主要因素[22]与本研究结论相一致。王刚等[23]、乔永等[24]研究发现,LPL表达与脂肪含量呈正相关关系,在发育早期哈萨克羊LPL酶活性随着年龄的增加而增加,在后期保持在一个稳定的水平,与本研究结果一致。徐小春等[25]研究结果显示,滩羊尾部脂肪中HSL酶活性总体随着月龄的增加呈现上升→下降→上升→下降的趋势,且在尾部脂肪、皮下脂肪、背最长肌中差异明显;Carole等[26]研究HSL基因表达水平在一定程度上与甘油三酯含量呈负相关的结果与本研究相一致。杨雪等[27]研究认为皮下脂肪、内脏脂肪的形成速率比肌内脂肪要高;罗鑫等[28]试验中肉羊在8月龄之前生长速度比较快,尤其在5月龄左右生长最快,此结论与本研究结果相一致。本研究结果表明,腹部脂肪细胞面积>皮下脂肪细胞面积>肠系膜组织细胞面积>背最长肌细胞面积。王金泉等[29]研究发现阿勒泰大尾羊在3月龄期间肾周脂肪细胞面积为2 000 μm2,皮下脂肪含量较少。这一结论与本试验多浪羊的研究结果一致,9月龄期间多浪羊脂肪细胞面积略高于阿勒泰羊脂肪细胞面积,可能由于阿勒泰羊生活在山地运动较多原因所致。本试验CB1基因在各个组织中均有表达,在脂肪组织中表达量最高,极显著高于其他组织,其次在肺脏、肝脏、肾脏、背最长肌中表达,在心脏和脾脏中表达量最低,与刘贤勋等[30]的研究结果相一致。CB1基因编码区分析可知H1为多浪羊主要单倍型,且CB1基因进化较为保守,受环境影响较小。魏星灿等[31]发现猪的CB1基因同源性较高,且在进化中受到纯化选择的作用;刘贤勋等[30]发现,布莱凯特黑牛的CB1基因在生物进化过程中非常保守,显示出高度保守的同源性。由CB1相关性分析可知,CB1基因表达量与腹部脂肪含量显著相关,与背最长肌、皮下脂肪、肠系膜组织脂肪含量极显著相关,与各部位脂肪细胞面积大小极显著相关,与脂肪代谢酶相关性随着部位不同表现出不同的相关性。
CB1基因编码区相对保守,环境对其影响较小。且多浪羊CB1基因表达量与脂肪细胞面积呈极显著正相关,与脂肪含量成显著正相关。与脂肪代谢酶相关性在不同部位、不同月龄表现出了不同的相关性。整体表现为CB1基因对LPL的影响高于HL的影响,高于对HSL的影响。
综上可知,多浪羊CB1基因可能不仅限于通过改变酶活性代谢的方式调控脂肪沉积,具体调控机制有待进一步研究。本研究为进一步揭示动物肉质调控的生物学机制提供理论和技术支持。
猜你喜欢
现代畜牧兽医(2022年3期)2022-12-06
散文诗世界(2019年10期)2019-09-10
浙江临床医学(2018年8期)2018-10-22
中国兽医杂志(2018年12期)2018-04-25
水动力学研究与进展 B辑(2017年2期)2017-04-26
国外医药(抗生素分册)(2016年2期)2016-07-12
中国塑料(2016年7期)2016-04-16
中国继续医学教育(2016年3期)2016-03-28
湛江文学(2015年1期)2015-12-18
家电科技(2014年5期)2014-04-16
