
全国部分地区H9亚型禽流感病毒分子流行病学和遗传变异分析
2023-10-19 08:28马利芳牛登云段宝敏史秋颖陈卓冯敬敬鲍恩东
畜牧与兽医 2023年4期
马利芳,牛登云,段宝敏,史秋颖,陈卓,冯敬敬,鲍恩东
(1. 南京农业大学动物医学院,江苏 南京 210095;2. 天津渤海农牧产业联合研究院有限公司,天津 300308)
禽流感病毒(avian influenza virus,AIV)属于正黏病毒科A型流感病毒属,为单股负链RNA病毒,是引起家禽呼吸道疾病的主要病因之一。HA蛋白是禽流感病毒最主要的2种表面糖蛋白之一,在AIV复制过程中起关键作用,直接参与AIV的致病过程,是AIV的主要毒力因子和保护性抗原。根据HA抗原性不同可将AIV分为18种HA亚型(H1~H18)[1-2],其中H9亚型禽流感病毒(H9-AIV)是其重要成员,不仅能感染禽类、也能感染猪和人类等哺乳动物。但由于其低致病性特点,往往被人们所忽视,而且H9-AIV也没有被列入政府强制免疫计划。近年来,H9-AIV检出在各地不断得到报道[3-8],如从1992年H9N2首次在中国境内出现并开始报道以来,H9-AIV在短短几年内便发展成为全国大流行,且其HA经历了从BJ/94 Like、F/98 Like、Y280 Like的逐渐演化[1,9]。在当前中国家禽广泛接种禽流感疫苗的大环境下,H9-AIV正不断发生变异以逃避疫苗的选择性压力。因此,对于H9-AIV的全面监测,不仅对中国家禽的养殖、疫病防控具有重要意义,而且由于H9-AIV是高致病性AIV的重要基因供体,监测其变异方向对人类健康也同样重要。本文就2020—2021年间中国家禽间H9-AIV的流行特点和变异情况进行了调研。
1 材料与方法
1.1 样品来源
本研究收集的临床样本,主要包括拭子(咽喉拭子或泄殖腔拭子)、组织样本、死鸡等。检测样品主要为2020—2021年收集到的来源于山东、辽宁、河北、河南等省份的10 336份疑似H9亚型禽流感发病肉鸡(如AA、科宝、817等)和蛋鸡(如海兰灰、海兰褐)的临床样本。
1.2 仪器设备及材料
PCR仪(型号T100)为美国伯乐公司产品;核酸提取仪(型号NP968)为天隆公司产品;高通量组织研磨仪(型号SCIENTZ-48)为宁波新芝生物科技有限公司产品;低温冷冻离心机(型号5427R)为eppendorf公司产品;RNA酶抑制剂(RRI)、逆转录酶(M-MLV)等均购自宝生物工程(大连)有限公司;RT-PCR/PCR相关试剂均购自北京全式金生物技术有限公司;检测病毒分离用SPF鸡胚均购自梅里亚或济南赛斯公司;检测及测序引物由北京擎科新业生物技术有限公司合成。

M. DL1000 DNA Marker;1~6. 检测样品;7. 阳性对照
1.3 引物设计与合成
检测引物参照NY/T 772—2013提供的H9-AIV基因的引物序列:H9-487-F为5′-CTCCACACAGAGCAYAATGG-3′;H9-487-R为5′-GYACACTTCTTCTTCTRTCG-3′,引物扩增长度为487 bp。
在NCBI中查找到HA基因的参考核苷酸序列,设计HA测序引物:H9-F为5′-AGCAAAAGCAGGGGAATG-3′;H9-R为5′-TTATATACAAATGTTGCATCTGCAAG-3′,引物扩增长度为1 683 bp。引物委托北京擎科新业生物技术股份有限公司合成。
1.4 样品处理
针对不同样本,将采集到的拭子溶解在灭菌PBS中待检,或将采集到的组织样本或病死鸡样本(重点采集气管、脾脏、肺脏、肝脏、肾脏等)用剪刀剪碎,加入等体积PBS,反复冻融3次,用组织研磨仪打碎,10 000 r/min离心10 min,取上清液至4 ℃待检。
1.5 核酸提取和RT-PCR鉴定
用移液器吸取200 μL上清液用核酸提取仪提取RNA,进行反转录。反转录体系为:随机引物(25 μmol/L)4 μL、RNase free H2O 20 μL、模板RNA 4 μL,总体积为28 μL。反应程序为:70 ℃ 10 min后迅速在冰上急冷2 min以上。在上述管中加入下列反转录反应液:5×M-MLV Buffer 8 μL、dNTP Mixture(各10 mmol/L)2 μL、RNase Inhibitor(40 U/μL)1 μL、M-MLV(200 U/μL)1 μL,总体积为40 μL。反应程序为:42 ℃ 1 h,70 ℃ 15 min后冰上冷却,得到的cDNA溶液用于PCR扩增。
20 μL检测用扩增体系:ddH2O 11.6 μL、10×Buffer 2 μL、dNTP Mixture 1.6 μL、引物H9-487-F/R 1.0/1.0 μL,TaqDNA酶 0.3 μL、cDNA 模板2.5 μL,总体积20 μL。反应程序为:95 ℃预变性5 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 35 s,扩增34个循环;72 ℃延伸10 min。扩增结束后,进行琼脂糖凝胶电泳。
1.6 病毒分离
将RT-PCR检测阳性的病料经双抗处理后,接种9~10日龄SPF鸡胚,每胚100 μL,弃去24 h内死亡的鸡胚,以后每隔8 h照胚1次,记录鸡胚死亡时间,接种后72 h收获尿囊液,进行血凝和血凝抑制试验。
1.7 测序和分析
将病毒分离结果为阳性的尿囊液提取核酸,进行RT-PCR,胶回收产物送测序公司进行测序。拼接后的序列与GenBank中的参考序列(表1),用MegAlign软件进行同源性比对,利用MEGA 5.0软件进行遗传进化分析。

表1 参考毒株序列在NCBI上的登录信息
50 μL测序用扩增体系:ddH2O 20.5 μL、10×Buffer 5 μL、dNTP Mixture 4 μL、引物H9-F/R 2.5/2.5 μL、rTaqDNA酶0.5 μL、cDNA模板2.5 μL,总体积为50 μL。反应程序:95 ℃预变性5 min;95 ℃ 30 s,50~52 ℃ 30 s,72 ℃ 1 min,扩增34个循环;72 ℃延伸10 min。12 ℃保存。
2 结果
2.1 H9-AIV检测率
统计H9-AIV核酸检测结果发现,2020—2021年期间送检的10 336份样品中,有1 665份样品为H9-AIV检测阳性,图1显示的是其中部分样品的RT-PCR鉴定电泳结果,H9-AIV阳性率为16.1%。
从检测样品的发病鸡类型分析,商品肉鸡的检出率最高,其次为商品黄鸡,父母代比商品代检出率低表2。从送检样本的月份检出率来看,2020年秋季到2021年春季期间,H9-AIV阳性检出率较高(表3)。

表2 2020—2021年不同类型鸡H9-AIV检出统计

表3 2020—2021年H9-AIV检出统计
2.2 病毒分离
选择RT-PCR检测阳性的部分样品进行鸡胚接种以分离病毒,共获得920株H9-AIV,分离率为92.2%(920/998),其中12份是尿囊腔接种呈阴性后,通过卵黄囊接种或绒毛尿囊膜接种呈阳性。病毒接种鸡胚后,弃掉24 h内死亡鸡胚,剩余鸡胚出现胚体表面有针尖大小的红点、全身发红、弱胚等病变;部分样品在48~72 h内死亡,死亡鸡胚剖检出现内脏出血等病变(图2)。
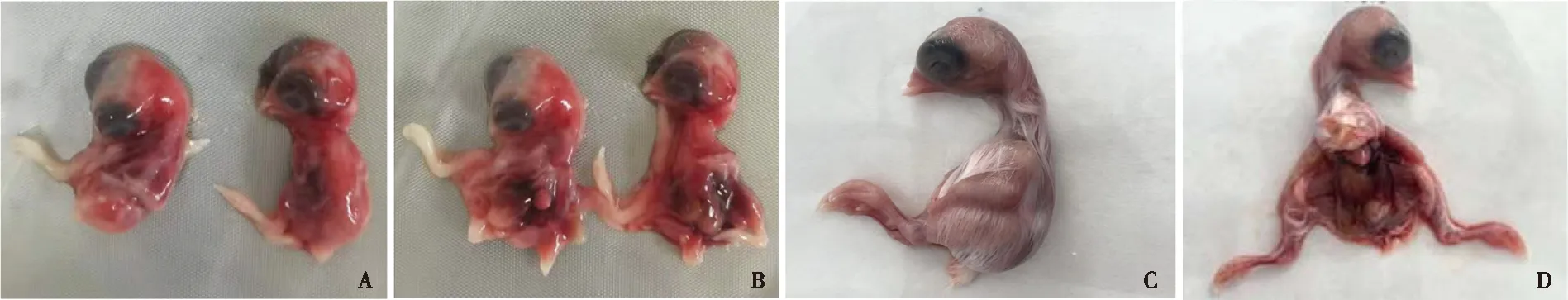
A、B. 鸡胚病料接种后的病变;C、D.正常鸡胚
2.3 HA基因同源性及遗传进化分析
对代表性分离毒株进行HA基因测序后,共获得464条HA序列,对上述序列进行基因同源性和遗传进化分析。结果显示,分离株与欧亚谱系F98 like代表毒株A/Chicken/Shanghai/F/98(H9N2)HA基因核苷酸和氨基酸同源性分别在85.5%~91.0%和84.5%~93.8%,而与Y280 like代表毒株A/Duck/Hong Kong/Y280/97(H9N2)核苷酸和氨基酸同源性分别在86.8%~92.0%和85.6%~94.6%,与WJ57核苷酸和氨基酸同源性分别在92.2%~99.9%和89.6%~99.8%。
遗传进化分析结果显示,2020—2021年全部采集后分离的毒株都属于h9.4.2.5分支,与其他的参考毒株相比,本研究中所分离的毒株与WJ57毒株的遗传关系较近。但在此分支上,根据遗传距离的远近,选取部分有代表性的分离毒株序列进行遗传进化分析,结果见图3。进化树共分为4个亚分支,其中21株分离毒株(4.5%,21/464)属于h9.4.2.5.1分支;101株分离毒株(21.8%,101/464)属于h9.4.2.5.2分支;22株分离毒株(4.7%,22/464)属于h9.4.2.5.3分支;320株分离毒株(69.0%,320/464)属于h9.4.2.5.4分支。
2.4 关键氨基酸位点分析
HA基因裂解位点对于AIV的致病力至关重要,本研究对测得的H9-AIV HA基因氨基酸序列进行分析,发现464株分离毒株中共有457株氨基酸裂解位点(335~338位氨基酸,本研究中全文采用H9编码序号)为R-S-S-R,3株为R-A-S-R,4株为R-S-N-R,均为典型的低致病性禽流感病毒特征性序列,这与之前的研究结果[13-14]一致。
本研究分析比较了H9-AIV分离毒株HA蛋白的11个受体结合位点(109、146、161、92、191、198、202、203、234、235、236位氨基酸),发现共有459株(459/464)第234位受体结合位点由Q突变为L,其他分离毒株分别突变为F/G/V;有92株(92/464)第198位受体结合位点为A,211株为T,154株为V,其余分离毒株分别突变为(H/K/M/Y)。其余受体结合位点均相对保守。此前有报道称,HA基因第381位氨基酸(对应H3第363位氨基酸)是影响H9-AIV的气溶胶特性的关键位点[16],本研究中427株分离毒株(92.0%,427/464)的该位点氨基酸由R突变为K,表明大多数分离毒株可能具有气溶胶的传播能力。
分析潜在糖基化位点结果显示,共计434株分离毒株与WJ57毒株的潜在糖基化位点一致,均为7个糖基化位点(29、141、298、305、313、492、551);7株含有8个潜在糖基化位点(其中6株由于在145位氨基酸S突变为N,由此增加1个潜在糖基化位点);14株分离毒株含有6个潜在糖基化位点,主要原因是缺少第29位或313位潜在糖基化位点。与F98 like和Y280 like相比,2020—2021年送检的H9-AIV已缺失了218位的潜在糖基化位点,新增加了313位潜在糖基化位点,这与资料报道的我国华东地区(2016—2020年)的H9N2变异基本一致[9,17],而与古小兵[18]所报道(2011—2012年)的研究结果不同,说明H9-AIV已经逐步发生变异,且变异趋势非常显著。
3 讨论
在以往的研究中,人们普遍认为分离AIV的最好是通过尿囊腔途径接种鸡胚。但有些毒株分离失败时,也可通过卵黄囊或绒毛尿囊膜途径接种分离[19],这一方法在本研究中也得到了有效应用。
本研究对分离毒株的HA基因进行了遗传进化分析,结果显示送检的所有H9-AIV分离毒株全部属于h9.4.2.5分支,且逐步演化出新的4个亚分支,而且主要位于h9.4.2.5.4分支上。本试验中共有459个分离毒株的受体结合位点第234位氨基酸由Q突变为L,占总样本的98.9%(459/464)。这与此前所报道的结果基本一致,预示H9-AIV进一步增强了对哺乳动物的α-2,6-Gal受体的结合能力;本研究中427株分离毒株(92.0%,427/464)的HA蛋白第381位氨基酸由R突变为K,但H9N2亚型AIV必须同时在PA的672位氨基酸为L,且HA的363位为K时才能进行气溶胶传播感染,另有其他次要因素也可影响病毒的气溶胶传播特性[16]。因此,本研究中的分离毒株是否具有气溶胶传播特性还需进一步研究才能确定。本研究中的分离毒株发生了HA基因第218位糖基化位点缺失和313位潜在糖基化位点增加。此前有报道称,这可能会影响H9-AIV对鸡和鸡胚的感染力及病毒在组织中的复制能力,但并不会影响病毒的抗原特性[20]。这对进一步分离和筛选H9-AIV疫苗候选毒株而言具有重要指导意义。
相比于其他亚型,H9亚型AIV的致病性低,但长期带毒会导致鸡只出现呼吸道症状和蛋鸡产蛋量下降的现象[21-22]。鸡感染H9亚型AIV后容易引起免疫抑制,进而继发感染其他病原,死亡率高,造成较大经济损失。Hu等[4]在研究2013—2017年从中国湖北分离出的H9N2时发现,其分离毒株与人源H9N2、高致病性AIV H10N8、H5N2、H5N6和H10N6的内源基因高度同源。研究认为,H9N2型AIV可能会充当其他AIV的基因供体,或者可能与其他AIV发生基因重组,从而造成危害性更大的新病毒病的流行[7,18,23-24]。一旦高致病性AIV通过气溶胶形式传播,那对社会造成的威胁将是不可估量的。综上,全面监测H9亚型AIV的变异具有重大意义。
猜你喜欢
科学大观园(2022年2期)2022-01-23
现代畜牧科技(2021年6期)2021-07-16
中国病理生理杂志(2015年8期)2015-12-21
动物医学进展(2015年10期)2015-12-07
医学研究杂志(2015年12期)2015-06-10
中国当代医药(2015年30期)2015-03-01
特产研究(2014年4期)2014-04-10
癌变·畸变·突变(2014年2期)2014-03-01
郑州大学学报(理学版)(2014年3期)2014-03-01
中国粮油学报(2014年7期)2014-02-06
