
青稞酸性磷酸酶基因 HvnACP2克隆和亚细胞定位研究
2023-10-17 06:13安立昆任晴雯姚晓华姚有华吴昆仑
西北农业学报 2023年10期
安立昆,任晴雯,姚晓华,姚有华,吴昆仑
(1.青海大学 农林科学院,西宁 810016;2.青海省青稞遗传育种重点实验室,西宁 810016;3.国家麦类改良中心青海青稞分中心,西宁 810016)
青稞(HordeumvulgareL. var.nudumHook. f.)是大麦的变种,由于其颖壳与籽粒分离,也称为裸大麦。青稞籽粒富含纤维素、蛋白质、维生素、β-葡聚糖,是重要的粮食、饲料和食品加工原料,青稞是最具有青藏高原地区特色的作物之一[1-4]。青藏高原地区气候极为恶劣,尤其是土壤极为贫瘠,长期的进化和筛选使得青稞对贫瘠的土壤具有较强的适应性[5-6]。随着青稞新品种的不断推广,种植面积逐年增大,已成为青海省农业独具地方特色的作物,在青海省农业经济中占有重要地位[7-9]。
磷是植物三大营养素之一,其重要性仅次于氮元素,磷元素的缺乏对作物的产量和品质有着严重的影响。农业生产中的磷肥主要源于磷矿开采,作为不可再生资源全球磷矿储量都在不断下降,尤其在中国由于农业生产中磷肥的利用效率不高,施入土壤中的磷不能得到有效利用,流失后反而对生态环境造成影响[10]。在土壤中大部分有效磷会被土壤吸附和固定,极难溶解和转移,即使磷含量较高的土壤中也只有少量磷元素可被植物吸收和利用,在土壤中植物对磷的利用效率远低于其他元素[11-13]。青藏高原地区是青稞的主产区,土壤极为贫瘠,生态极为脆弱,过量施用磷肥会导致更严重的生态灾难,不利于青藏高原地区农业的可持续发展。所以研究青稞磷吸收和利用相关基因,提高其对土壤中的磷利用率对于减少青稞生产中的磷肥施用,促进青稞生产的可持续发展具有重要意义。
酸性磷酸酶(Acid Phospoatase,ACP)是植物体中广泛存在的一种水解酶,可以水解磷酸酯并释放出磷酸,在低磷条件下植物体内尤其是根系会分泌大量酸性磷酸酶促进有机磷的水解,在植物磷吸收和利用方面有着重要作用[13-14]。酸性磷酸酶基因属于一类高度保守的基因家族,不同家族成员通过不同的表达模式调控在植物体中发挥作用,维持植物体中磷元素的吸收和利用[15]。Zhang等[16]采用全基因组关联GWAS鉴定到了一个大豆磷高效利用基因GmACP1,在大豆中过表达GmACP1基因可以明显提高大豆对磷的利用效率,并认为GmACP1P基因为通过分子育种手段准确培育磷高效利用大豆品种提供了基础。李靖怡[17]对水稻OsACP1进行研究发现OsACP1在水稻中只受低磷的诱导表达,在水稻中过表达OsACP1会使正常生长条件下的水稻出现磷中毒症状,在缺磷培养条件下恢复正常,OsACP1定位在内质网和高尔基体膜上,认为OsACP1的功能可能是在内质网和高尔基体膜中代谢磷酸乙醇胺和磷酸胆碱等有机磷,维持缺磷条件下细胞中的磷稳态。
目前关于酸性磷酸酶的研究主要集中在水稻、玉米、小麦、大豆等主要作物中,关于青稞中酸性磷酸酶基因尚未见研究报道。本试验从青稞‘肚里黄’中克隆得到酸性磷酸酶基因HvnACP1对其基因和蛋白进行生物信息学分析,并对其亚细胞定位进行研究。为青稞磷高效利用相关基因研究和利用分子育种手段进行磷高效利用品种培育提供一定基础。
1 材料与方法
1.1 试验材料
青稞品种‘肚里黄’和本氏烟草(Nicotianabenthamiana)由青海省青稞遗传育种重点实验室保存,青稞和烟草于2021年4月种植于青海大学农林科学院实验温室中。摘取生长10 d后青稞的第3叶片进行DNA和RNA提取。采用生长1月后的烟草第3或第4叶片进行试验。
PCR及其产物连接、cDNA合成、大肠杆菌感受态细胞分别采用北京全式金有限公司的TransTaq○RDNA Polymerase High Fidelity(AP131-01)试剂盒,pEASY○R-Blunt Cloning Kit(CB101-01)试剂盒,TransScript○RFirst-Strand cDNA Synthesis SuperMix(AT301-02)试剂盒以及Trans1-T1大肠杆菌感受态细胞。BP和LR反应试剂盒购自Invitrogen公司。引物合成和测序由北京擎科生物科技有限公司完成。
1.2 试验方法
1.2.1 青稞DNA、RNA提取和cDNA合成 采用尿素法[18-19]提取青稞叶片DNA,TRIzol法提取RNA。cDNA合成参照TransScript○RFirst-Strand cDNA Synthesis SuperMix(AT301-02)试剂盒说明完成,所有样品置于-20 ℃保存。
1.2.2HvnACP2基因和启动子区域序列克隆 将已报道的谷子酸性磷酸酶基因SiACP序列输入NCBI(https://www.ncbi.nlm.nih.gov/)中进行搜索获得大麦的HvACP基因序列和结构信息[14]。根据大麦HvACP2基因序列及起始密码子ATG前2 000 bp左右序列设计引物。基因克隆正向引物:F:5′-ATGGAGGTGCTCGCGG-3′,反向引物:R:5′-CTAGTTGGTGTGGGTT-3′。启动子区域序列克隆正向引物:F:5′-ATGTAATGTTGTAGTTTATTGC-3′反向引物:R:5′-GGCGTGCGCCGCTAGGAG-3′。采用PCR以cDNA为模板克隆HvnACP2cDNA序列,以基因组DNA为模板克隆HvnACP2启动子区域序列。按以下程序进行扩增:94 ℃ 2 min;98 ℃ 10 s, 58 ℃ 30 s,68 ℃ 1 min(扩增启动子区域序列为2 min),35个循环。反应完毕后对PCR产物进行琼脂糖凝胶电泳,确定PCR产物片段大小无误后,参照pEASY-Blunt Cloning试剂盒说明书,将PCR产物进行连接并转化大肠杆菌,筛选阳性克隆并送测序。获得cDNA序列后采用DNAMAN7对cDNA序列中的ORF区域进行翻译获得HvnACP蛋白 序列。
1.2.3 青稞HvnACP2生物信息学分析 采用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对启动子区域元件进行预测;采用Protparam(https://web.expasy.org/protparam/)对蛋白理化性质进行预测分析;采用TMHMM Server v. 2.0(http://www.cbs.dtu.dk/services/TMHMM/)对蛋白跨膜结构进行预测分析;采用kinasephos2(http://kinasephos2.mbc.nctu.edu.tw/)对蛋白磷酸化位点进行预测分析;采用SignalP(http://www. Cbs. Dtu. Dk /services /SignalP/)对蛋白信号肽进行预测分析;采用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)对蛋白二级结构进行分析;采用蛋白同源建模软件Swiss-Model(http://swissmodel.expasy.org/)进行蛋白三级结构同源建模。
在植物基因组数据库Gramene(http://www.gramene.org/)和NCBI(https://www.ncbi.nlm.nih.gov/)中进一步搜索得到其他植物的同源蛋白序列:象草(Cenchruspurpureus)CpACP2、狗尾草(Setariaviridis)SvACP2、黍(Setariaviridisvar.hallii)PhACP2、糜子(Panicummiliaceum)PmACP2、福尼奥小米(Digitariaexilis)DeACP2、南荻(Miscanthus lutarioriparius)MlACP2、玉米(ZeamaysL.)ZmACP2、康藏嵩草(Carexlittledalei)ClACP2、番茄(Solanumlycopersicum)SlACP2、蓝星睡莲(Nymphaeacolorata)NcACP2、油菜(Brassicanapus)BnACP2、烟草(Nicotianaattenuata)MaACP2、粳稻(OryzasativaL.subsp. Japonica)OsACP2、籼稻(OryzasativaL.subsp. Indica)OsACP2、二型花(Dichantheliumoligosanthes)DoACP2、小米(Setariaitalica)SiACP2、弯叶画眉草(Eragrostiscurvula)EcACP2、二穗短柄草(Brachypodiumdistachyon)BdACP2、小麦(Triticumaestivum)TaACP2、野甘蓝(Brassicaoleraceavar.oleracea)BoACP2、萝卜(Raphanussativus)RsACP2、盐芥(Eutremasalsugineum)、EsACP2、小菥蓂(Microthlaspierraticum)MeACP2、筷子芥(Arabisnemorensis)AnACP2、荠蓝(Camelinasativa)CsACP2、荠菜(Capsellarubella)CrACP2。采用DNAMAN 7.0将青稞HvnACP2蛋白序列与大麦、水稻、二穗短柄草、烟草、番茄的同源蛋白进行序列比对,并根据NCBI中提供的相应蛋白信息分析结构域和关键位点。采用MEGA7对所有植物的同源蛋白序列以最大似然法构建系统进化树。
1.2.4 亚细胞定位表达载体构建 采用载体pSATN1-mkate利用gatway技术构建亚细胞定位载体。设计带有attB位点的引物对HvnACP2CDS序列进行PCR克隆,回收PCR产物后,按照BP和LR反应试剂盒说明书完成载体构建,并对载体进行测序验证。将构建好的载体通过冻融法转化入农杆菌EHA105中。制备好农杆菌重悬液后将菌液注射入烟草叶片下表皮,放入完全黑暗环境,温度22 ℃,培养3 d后将叶片切成1 cm2大小,放置于Nikon C2-ER激光共聚焦显微镜下观察并拍照。
2 结果与分析
2.1 青稞 HvnACP2基因生物信息分析
2.1.1 基因结构和启动子功能元件预测分析 测序结果表明青稞HvnACP2和大麦HvnACP2基因和启动子区域DNA序列完全一致。根据植物基因组数据库Gramene中获得的大麦HvACP2基因结构信息分析,青稞HvnACP2含有2个外显子和1个内含子(图1)。
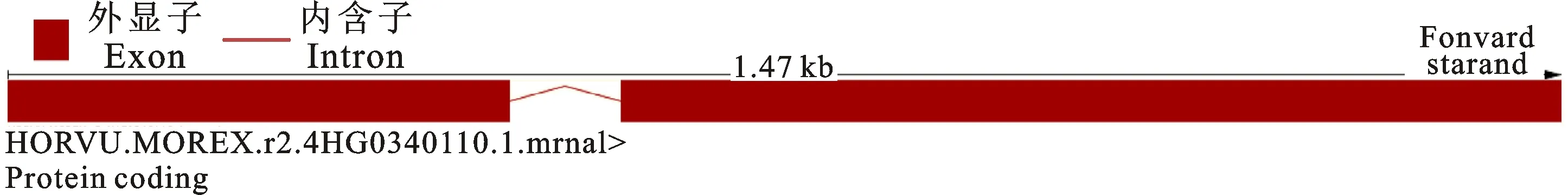
图1 青稞 HvnACP2基因结构Fig.1 Gene structure of HvnACP2
将克隆得到的HvACP2基因启动子区域序列输入启动子元件分析网站PlantCARE中进行分析(表1)。发现HvnACP2启动子区域具有大量的TATA-box和CAAT-box启动子核心元件,3个脱落酸响应元件2个茉莉酸响应元件,此外还有1个分生组织表达顺式作用元件,8个光响应元件,1个厌氧响应元件,2个MYB转录因子结合位点。
2.1.2 青稞HvnACP2蛋白理化性质和结构分析 通过生物信息学软件预测分析发现 HvnACP2由454个氨基酸组成,为亲水性稳定蛋白(表2)。HvnACP不具有跨膜结构(图2),有丝氨酸(Ser)5个,苏氨酸(Thr)2个,酪氨酸(Tyr)5个共12个磷酸化位点(图3)。具有信号肽序列:MEVLAVLFLLAAHAASA,切割位点在第17至18位氨基酸之间(图4)。HvnACP2二级结构蛋白结构α螺旋共85个、无规则卷曲共245个、延长链共107个,β转角共17个分别占氨基酸总数的18.72%、53.96%、23.57%、3.74%,其三级结构和二级结构预测分析结果基本一致(图5和图6)。

表2 青稞HvnACP蛋白质理化性质分析Table 2 Physical and chemical properties of HvACP2 protein

图2 青稞HvnACP2蛋白跨膜结构预测分析Fig.2 Transmembrane structure prediction of HvnACP2 protein

图3 青稞HvnACP2蛋白磷酸化位点预测Fig.3 Prediction of HvnACP2 phosphorylation sites

图4 青稞HvnACP2蛋白信号肽预测Fig.4 Prediction of HvnACP2 signal peptide

蓝色.α螺旋;红色.延伸链;橙色.无规则卷曲;绿色.β转角

图6 青稞 HvnACP2三级结构预测分析Fig.6 Tertiary structure prediction of HvnACP2
2.1.3 HvnACP2蛋白氨基酸序列对比和同源进化分析 蛋白序列分析发现大麦 HvACP2和HvnACP2蛋白序列完全一致,进一步与水稻、二穗短柄草、烟草、番茄的同源蛋白序列分析发现其都具有信号肽结构、紫色酸性磷酸酶结构域、金属磷酸酶结构域和相同的金属离子结合位点(图7)。将青稞HvnACP2与28种植物的ACP2蛋白序列构建系统进化树,发现大麦HvACP2和青稞HvnACP2与小麦TaACP2亲缘关系最近(图8)。

图7 HvnACP2蛋白氨基酸序列对比分析Fig.7 Amino acid sequence alignment of HvnACP2 with homologous genes of other species

图8 青稞HvnACP2与其他植物同源蛋白系统进化树Fig.8 Phylogenetic tree of HvnACP2 with other species
2.2 HvnACP2亚细胞定位研究
将带有载体pSATN1- HvnACP2::mkate的农杆菌EHA105注射入烟草叶片中,对HvnACP2进行亚细胞定位研究发现红色荧光信号分布于细胞膜上,说明HvnACP2定位于细胞膜上(图9)。
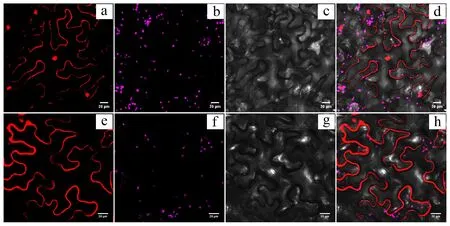
a~d.分别为35::mkate空载体荧光场、叶绿体自发荧光场、明场和叠加场;e~h.分别为35:HvnACP2-mkate荧光场、叶绿体自发荧光场、明场和叠加场
3 结论与讨论
酸性磷酸酶家族是植物利用磷元素重要的基因家族之一。酸性磷酸酶按功能可分为细胞内和分泌型的两类。细胞内的酸性磷酸酶可以将衰老组织中的磷素进行再利用,分泌型的酸性磷酸酶可分泌到细胞外活化被土壤吸附的磷,将土壤中的有机磷转化为无机磷,然后供植物体的利用,酸性磷酸酶对植物在应对低磷胁迫应答中具有重要的作用[20]。有一些研究报道酸性磷酸酶在调控植物碳代谢、细胞壁合成和抗病等方面有重要的作用[21]。
本研究克隆了酸性磷酸酶HvnACP2基因和启动子区域序列,对其基因和蛋白进行生物信息学分析,并对其亚细胞定位进行研究。很多研究表明大部分植物酸性磷酸酶都具有信号肽结构,但不同的植物酸性磷酸酶亚细胞定位位置不同,有定位于细胞质、质膜、细胞壁甚至细胞外[22-26]。侯立江等[27]对西尔斯山羊草紫色酸性磷酸酶PAP1进行分析也发现其具有信号肽结构,PAP1蛋白包含金属离子结合中心,预测它属于金属蛋白,亚细胞定位预测分析其定位于质膜的概率最高。本研究发现青稞HvnACP2同样具有信号肽、金属磷酸酶结构域和金属离子结合位点,并且定位于细胞质膜上。通过与其他物种的同源蛋白序列进行对比并构建系统进化树发现其系统进化树并没有分为典型的单双子叶两支,与大麦和青稞亲缘关系最近的是小麦TaACP2。赵雄伟等[14]对谷子酸性磷酸酶ACP家族基因进行鉴定并对其中的SiACP1耐低磷单倍型进行分析共发现了谷子中的13个ACP家族基因,生物信息学研究表明其蛋白都具有高度保守的磷酸酶结构域,谷子ACP家族基因与其他植物构建进化树分析发现,单双子叶植物ACP基因呈现明显的分化,通过转录组数据分析发ACP基因家族中不同成员的表达模式各有差异,通过基因关联和单倍型分析发现SiACP1启动子上的一个SNP与耐低磷相关性状显著相关。李睿等[15]对苹果紫色酸性磷酸酶相关基因MdPAP10进行克隆和研究发现MdPAP10基因包含5个外显子和4个内含子,其蛋白具有信号肽并包含一个磷酸酶结构域,MdPAP10在苹果根中表达量最高,叶片次之,果实中表达量最低,低磷培养条件下根中的MdPAP10受诱导表达明显,叶片中的表达量没有明显变化,在苹果愈伤组织中过表达MdPAP10可以提高其对低磷胁迫的耐受性,并且可以促进其他磷吸收利用基因MdPHT1.1、MdPT2和MdPHO1等相关基因表达。
青稞作为青藏高原地区的特有作物,减少青稞生产中的化肥施用是保障青藏高原地区农业生态和青稞产业可持续发展的关键之一。目前关于青稞中磷高效利用的基因研究报道极少。本研究克隆了青稞酸性磷酸酶HvnACP2基因,并进行了生物信息学分析和亚细胞定位研究,为青稞磷高效利用基因研究和分子育种提供了一定基础。
猜你喜欢
星星·诗歌原创(2022年1期)2022-04-16
翠苑(2019年4期)2019-11-11
中国组织化学与细胞化学杂志(2017年1期)2017-06-15
浙江农业学报(2017年3期)2017-04-08
中国烟草学报(2016年1期)2016-11-16
西藏科技(2016年8期)2016-09-26
广西林业科学(2016年1期)2016-03-20
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
医学研究杂志(2015年8期)2015-06-22
中国医学科学院学报(2013年6期)2013-03-11
