
高粱的玉米黄质环氧化酶基因SbZEP 表达与DNA 变异分析
2023-10-14 19:04柴文婷李华天赵珊珊杨博慧刘卓君王新宇郝昱宁史小霞郭子浩吕晋慧张春来
山西农业科学 2023年10期
柴文婷 ,李华天 ,赵珊珊 ,杨博慧 ,刘卓君 ,王新宇 ,郝昱宁 ,史小霞 ,郭子浩 ,吕晋慧 ,张春来
(1.山西农业大学 农学院,山西 太谷 030801;2.山西农业大学 林学院,山西 太谷 030801)
玉米黄素(Zeaxanthin,Zea)在叶黄素循环的脱环氧化反应中形成,是一种具有强抗氧化活性的类胡萝卜素,广泛应用于饲料、化妆品、食品和医疗等方面[1]。在陆地植物中它参与光系统II(PSII)中多余光能的非光化学淬灭,将多余的能量作为热量耗散来防止光抑制[2]。玉米黄素可催化转化为紫黄质(Violaxanthin),这是ABA 生物合成和叶黄素循环的关键反应,这一反应不仅对于ABA 生物合成有重要作用,而且有助于类胡萝卜素的生物合成。
玉米黄质环氧化酶(Zeaxanthin epoxidase,ZEP),是一种参与玉米黄素积累和转化的酶,在植物抗渗透压和干旱胁迫、种子发育和休眠等次生代谢过程和逆境胁迫响应中发挥重要作用[3]。ZEP 由类囊体腔中的紫黄质脱环氧酶(Violaxanthin deepoxidase,VDE)催化。VDE 活性受到类囊体腔pH 值的严格调节,确保玉米黄素形成和NPQ 诱导仅在使光合电子传输饱和的光强度下发生[4]。
目前关于ZEP基因研究已获得大量成果,ZEP基因主要参与类胡萝卜素的合成代谢。LEE等[5]将橙辣椒品种与黄辣椒品种杂交进行遗传分析,并将成熟水果中的类胡萝卜素含量进行HPLC分析,结果表明,辣椒果实中的橙色由ZEP中单个隐性突变控制。CRUET-BURGOS 等[6]对345 种高粱种质进行的全基因关联分析,挖掘出玉米黄质环氧化酶基因是控制叶黄素和β-胡萝卜素变异的主要基因,高粱类胡萝卜素存在寡源遗传。SUEMATSU 等[7]通过比较黄肉栽培品种及2 个白肉突变体的转录组,检测到38 个与贮藏根中类胡萝卜素生物合成有关的基因,并对根中类胡萝卜素成分进行分析,发现ZEP旁系同源物参与类胡萝卜素的积累。此外,ZEP基因在植物响应各种环境胁迫的反应中起重要作用,如小麦ZEP1基因可以增强小麦对条锈病的防御能力[8]。
高粱属于杂粮作物的一种,营养成分高于玉米,是有较高的叶绿素、蛋白质和花青素含量[9-10]的杂粮。但较其他粮食作物,其类胡萝卜素等维生素含量较低,而通过调控玉米黄质环氧化酶的活性,提高籽粒类胡萝卜素含量是当前功能农业研究的重点。山西农业大学特色作物基因组学与遗传改良团队积累基因组、转录组、表型组资源,熟练掌握了生物信息分析与分子生物学实验技术,解析高粱抗逆和耐瘠性状形成机制,已经对高粱HSF[11]、ClC[12]、NRT1[13]、NRT2/3[14]和CDL[15]进行了全基因组鉴定、表达和DNA 变异分析,而在高粱种质中有关ZEP基因相关分子机制的研究较少。
本研究采用生物信息学方法筛选和鉴定SbZEP基因,并进行初步预测和分析,对SbZEP基因进行蛋白及基因分析和DNA 变异分析,旨在为研究SbZEP基因的功能提供必要的理论基础与试验依据。
1 材料和方法
1.1 高粱ZEP 基因成员鉴定
以拟南芥(Arabidopsis thaliana)的ZEP氨基酸序列作为参考,在Phytozome(https://phytozomenext.jgi.doe.gov)与NCBI(http://www.ncbi.nlm.nih.gov/)的数据库中完成Blast 查找和对比,获得ZEP基因蛋白序列、基因序列、CDS 序列。拟南芥(Arabidopsis thaliana)、玉米(Zea mays)、胡萝卜(Daucus carota)、向日葵(Helianthus annuusL.)、谷子(Setaria italica)、小果咖啡(Coffea arabica)、大豆(Glycine max)、水稻(Oryza sativax)、蓖麻(Ricinus communisL)和藜麦(Chenopodium quinoa)的ZEP蛋白序列来自Phytozome 数据库。
1.2 高粱ZEP 基因生物信息学分析
采用ExPASY-ProtParam tool 数据库(https://web.expasy.org/protparam/)对相应基因的蛋白质理化性质完成解析;用GSDS 在线工具(http://gsds.cbi.pku.edu.cn/)绘制高粱ZEP基因结构图;在NCBI-CDD(https://www. ncbi. nlm. nih. gov/Structure/bwrpsb/bwrpsb.cgi)上下载SbZEP 保守结构域相关数据,用TBtools 进行可视化。运用SignalP-5.0 Server数据库(https://www.services.healthtech.dtu.dk/services/SignalP-5.0/)检验SbZEP 信号肽,在PSORT 数据库(https://www.genscript.com/psort.html)对SbZEP 进行亚细胞定位。利用SOPMA 数据库(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl)完成SbZEP 二级结构预测,在Phyre2数据库(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi)完成蛋白质的三级结构模型的建立与分析,在SAVES v6.0 数据库(https://saves.mbl.ucla.edu/)检验三级结构是否正确。
NCBI 上下载SbZEP基因启动子序列,利用在线软件Plant CARE(https://bioinformatics. psb.ugent.be/webtools/plantcare/html/)进行顺式作用元件分析。使用邻接法(Neighbour joining,NJ)构建高粱ZEP基因系统进化树。在Phytozome 中下载相关基因表达量数据,采用TBtools 软件绘制热图。同时在Phytozome 数据库下载高粱DNA 重测序数据,完成SbZEP基因的SNP 的鉴定与注释。通过STRING 数据库(https://cn.string-db.org),构建高粱ZEPs互作蛋白网络图。
1.3 SbZEP 基因表达分析
高粱品系R111 的茎、胚珠、种子和SSA1 的种子转录组,在百迈客公司完成获取表达量,并采用Exce 11 软件绘图。
2 结果与分析
2.1 SbZEP 基因家族基本信息分析
利用高粱各组织的转录组测序数据,得到2 个SbZEP基因。将Sobic.006G097500和Sobic.010G1 81100分别编号为SbZEP1和SbZEP2;2 个基因分别位于6 号染色体46715530—46722401 和10 号染色体52039617—52041596 处,基因长度分别为6 873、1 980 bp(表1)。

表1 SbZEP 基因基本信息Tab.1 Basic information of SbZEP genes
2.2 SbZEP 基因结构和蛋白结构域分析
SbZEP基因结构显示(图1),SbZEP1和SbZEP2分别有15、3 个内含子,且SbZEP1在1~3 kb 处存在比SbZEP1更长的内含子,推测SbZEP1行使更多的基因功能。

图1 SbZEP 基因结构Fig.1 Structure of SbZEP genes
SbZEP 蛋白结构域分析可知(图2),SbZEP 蛋白共有6 种结构域,SbZEP1 和SbZEP2 均含有UbiH 和FAD_binding_3 这2 种保守结构域,除共有结构域外,SbZEP1 还包含PLN02927、FHA、Ubi-OHases、COQ6,其中,FHA 结构域控制叶黄素循环和脱落酸生物合成;SbZEP2 还含有Ubi-OHases,参与有氧泛醌生物的合成途径。推测SbZEP基因与结构域功能密切相关。

图2 SbZEP 蛋白结构域Fig.2 SbZEP protein domains
2.3 SbZEP 蛋白质理化性质分析
从表2 可以看出,SbZEP1 和SbZEP2 蛋白质氨基酸数目分别为672、436 个;分子质量大小分别为73 907.68、47 149.42 u;理论等电点分别为8.02 和5.45,SbZEP1、SbZEP2 均为碱性蛋白;不稳定系数(>40%)的解析显示,SbZEP 蛋白均是不稳定蛋白;SbZEP 蛋白的脂肪溶解系数较低,说明该蛋白的流动性较差;SbZEP1、SbZEP2 蛋白的平均亲水性分别为-0.371 和-0.094,表明SbZEP 蛋白质均为亲水性蛋白。

表2 SbZEP 蛋白质理化性质Tab.2 The physicochemical properties of SbZEP proteins
2.4 SbZEP 蛋白质信号肽及亚细胞定位分析
根据蛋白质信号肽分布(表3),SbZEP1 和SbZEP2蛋白质信号肽分值分别为0.004 5和0.264 3,小于0.5 为非分泌性蛋白。蛋白质的亚细胞定位预测证实,SbZEP1 蛋白定位在内质网上,SbZEP2 蛋白定位在叶绿体上。
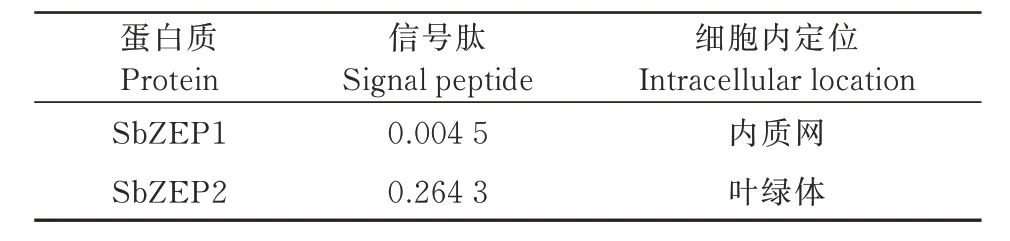
表3 SbZEP 蛋白质亚细胞定位Tab.3 Subcellular localization of SbZEP proteins
2.5 SbZEP 蛋白质二级结构分析
SbZEP 蛋白的二级结构解析结果如表4 所示。

表4 SbZEP 蛋白质的二级结构分析Tab.4 Secondary structure analysis of SbZEP protein %
从表4 可以看出,高粱ZEP 家族各成员中均含有4 种二级结构,其中,α-螺旋和无规则卷曲占比最高,这2 种形式涵盖的氨基酸比例>70%,由此证明,该基因蛋白主要由这2 种构成;而β-折叠和延伸链含量较少。以上结果表明,α-螺旋与无规则卷曲为SbZEP 蛋白的主要结构成分,延伸链与β-折叠则分散在蛋白质里。
2.6 SbZEP 蛋白质三级结构模型构建
SbZEP 蛋白的三级结构组成部分如图3 所示,其中,α-螺旋与无规则卷曲所占比例较大,且SbZEP1和SbZEP2 蛋白的三级结构模型有所差异;使用Ramachandran plot analysis 软件建立SbZEP 蛋白三级结构模型,可信度>90.00%。

图3 SbZEP 蛋白质三级结构Fig.3 Tertiary structure of SbZEP proteins
2.7 高粱ZEP 基因系统进化树分析
对高粱、拟南芥、胡萝卜、玉米、向日葵、谷子、小果咖啡、大豆、水稻、蓖麻和藜麦共24 个的ZEP基因家族进行聚类分析(图4)。

图4 ZEP 基因家族成员的系统进化树Fig.4 Phylogenetic tree of ZEP gene family members
图4 表明,ZEP基因家族可分为4 个亚族,其中,第Ⅰ亚族内分布有3 个藜麦和2 个小果咖啡,均为双子叶植物,拟南芥分布在第Ⅳ亚族,高粱分布在第Ⅱ、Ⅳ亚族中;SbZEP1与Rc30115.m001209、SbZEP2与Seita.7G116800互为直系同源基因,证明高粱和谷子的ZEP基因在演化过程中同源性差异更小;试验结果中单子叶植物如玉米、高粱、水稻、谷子与向日葵、藜麦、蓖麻、拟南芥等双子叶植物对比得知,二者大多不能聚合在同一分支下,说明ZEP基因在进化过程中出现单子叶和双子叶分化的现象。
2.8 SbZEP 顺式作用元件分析
由图5 可知,SbZEP家族启动子共预测到14 种顺式作用元件,其中,SbZEP1和SbZEP2均有参与激素响应(TGACG-motif、CGTCA-motif 茉莉酸甲酯、ABRE脱落酸)、光响应(G-box、GATA-motif、TCT-motif)、MBS 干旱诱导、ARE 厌氧诱导表达和O2-site 玉米醇溶蛋白代谢调节元件。此外,SbZEP1还含有CAT-box 与分生组织表达相关调控元件,表明SbZEP基因可能参与光反应和防御反应过程,且SbZEP1可能参与诱导分生组织表达过程。

图5 SbZEP 基因启动子区顺式作用调控元件的功能注释Fig.5 Functional annotation of the cis-acting regulatory elements of gene promoters of SbZEP
2.9 SbZEP 基因表达分析
SbZEP基因表达热图分析显示(图6),SbZEP基因在高粱不同生育期不同器官内表达水平存在差异;SbZEP1在生育期内部分部位的表达水平相对高一些,普遍在茎部和叶片表达,在叶轮生中间时期表达水平最高,同时在叶上部和叶下部区域表达水平较高。通过SbZEP1基因的完整表达时期结果可以得出,表达水平由高到低依次为叶轮生中间时期、叶上部和叶下部部位、上轮生叶;在根部和花序中表达水平较低。SbZEP2表达总体低于SbZEP1,且在各部位表达水平均较低。

图6 在高粱不同发育时期不同器官SbZEP 的表达Fig.6 Expression of SbZEP at various tissues at different developmental stages
SbZEP在高粱种子、胚珠和茎的表达如图7 所示,在茎、胚珠中的基因表达水平较高,2 个基因间表达水平有差异,SbZEP1的整体表达水平高于SbZEP2,SbZEP1的表达水平20 d 种子高于10 d。SbZEP2的表达水平SSA1 10 d 种子明显高于20 d。

图7 SbZEP 在高粱种子、胚珠和茎的表达水平Fig.7 Expression levels of SbZEP in sorghum seed,ovary,and stem
2.10 SbZEP 基因的DNA 变异分析
根据Phytozome 数据库内重测序结果得到24 个高粱品种SbZEP的SNP(表5),共筛选出9 个SNP 变异位点,不同品系对应不同位点的突变;其中,Std-Broomcorn 品系的SbZEP1在SBI-06的46716082、46716091、46716132、46720862 非同义编码SNP 变异,分别由甘氨酸Gly 变为谷氨酰胺Gln,由精氨酸Arg 变为赖氨酸Lys,由酪氨酸Tyr 变为丝氨酸Ser,赖氨酸Lys 变为苏氨酸Thr;E-Tian 和Keller 品系中SbZEP1在SBI-06的46717731 和46717733 上的突变,分别由苏氨酸Thr 变为丙氨酸Ala;赖氨酸Lys 变为苏氨酸Thr;在Ji2731 品系中,SbZEP1在SBI-06的46716260 处的突变,由天冬氨酸Asp 变为谷氨酸Glu。Tx642 和Segaolane 品系中SbZEP2在SBI-10的52040014、52040426 位点处,分别由组氨酸His 变为天冬酰胺Asn;丝氨酸Ser 变为丙氨酸Ala。这些氨基酸的改变,会导致翻译后做得到的蛋白质与之前有差异,将有一定概率影响到蛋白结构与功能。高粱籽粒的DNA 变异数据,在之后的试验中可被用在基因克隆和分子标记筛选。
2.11 SbZEP 蛋白互作分析
SbZEP全基因组的蛋白互作显示如图8所示,平均聚类系数为0.924;PPI富集P值为2.43×10-5。其中,SbZEP1分别与Sobic.06G049200(VDE domaincontaining protein)、Sobic.06G182200(sterol desaturase family)和Sobic.04G074000(Lycopene betacyclase)蛋白互作;SbZEP2分别与Sobic.06G049200和Sobic.10G276400(Chloroplast phytoene synthase 1)蛋白互作;根据基因功能预测显示(表6),Sobic.06G049200参与类胡萝卜素代谢过程。其中,Sobic.04G074000、Sobic.06G188200 和Sobic.10G2764 00 参与类胡萝卜素的生物合成与代谢。由此证明,SbZEP基因可能与类胡萝卜素的含量变化有关。

图8 SbZEP 蛋白互作网络Fig.8 Network of SbZEP protein interaction

表6 STRING 预测SbZEP 互作蛋白功能注释Tab.6 Function annotation of SbZEP interaction protein by STRING prediction
3 结论与讨论
高粱是抗逆性较强的作物之一,且抗逆境的能力比玉米、小麦等更强,提高高粱籽粒类胡萝卜素含量变成了当前功能农业研究的重点。目前,在烟草[16]和万寿菊[17]、拟南芥、水稻、玉米等植物中已经对其ZEP基因进行克隆和解析[18]。通过从全基因组水平上对SbZEP基因进行分析研究,完成了高粱种质中籽粒类胡萝卜素的全基因组关联分析[19-20]。
本研究在高粱全基因组中鉴定出2 个SbZEP基因,定位于高粱6 号与10 号染色体上,与大洋南绿藻中鉴定的ZEP基因个数相同[21]。高粱ZEP 蛋白含有UbiH 和FAD_binding_3 这2 种保守结构域,且ZEP1比ZEP2存在更多的结构域和内含子,表明SbZEP1基因存在更多的功能。系统进化分析表明,高粱与玉米的亲缘关系更近,ZEP基因进化过程中存在单双子叶分化现象。研究表明,NoZEPs基因的过表达可增加大洋南绿藻中紫黄质及其下游类胡萝卜素的含量,而导致玉米黄质含量的减少[21]。SbZEP 蛋白互作分析表明,ZEP 蛋白调控类胡萝卜素生物合成与代谢,但在高粱中ZEP 蛋白参与调控胡萝卜素和玉米黄素合成代谢相关具体分子机制还有待进一步研究。
SbZEP基因启动子多数含有响应光反应和防御反应的元件,包括茉莉酸甲酯响应元件(TGACGmotif、CGTCA-motif)和脱落酸响应元件ABRE。已有研究表明,玉米黄素在叶绿体中具有重要的光保护功能,强光条件下在类囊体膜中积累[22],也是SbZEP2在叶绿体内表达的原因之一。且脱落酸(ABA)是一种重要的类胡萝卜素衍生植物激素,是环境胁迫条件下调节生理过程的植物胁迫信号生物合成的关键酶[13],在植物对生物和非生物胁迫的反应以及各种生理和发育过程中起着至关重要的作用[23]。GaZEPs调节抗病性和对非生物胁迫的响应,如干旱、热冲击、低温、盐胁迫等[24]。这些结果预示了ZEP基因在高粱非生物胁迫的响应及光反应中的作用。
热图分析表明,SbZEP基因都在叶中高表达,在其他部位表达水平较低或不表达,这与LOU等[25]的分析结果基本一致。拟南芥ZEP 蛋白水平变化受干旱胁迫影响,ZEP 蛋白在ABA 生物合成和叶黄素循环中的不同功能,导致ZEP 蛋白的组织和应激特异性积累,最高水平的ZEP 存在于叶和根中[26]。在亚洲棉花中,叶中GaZEP表达水平与脱落酸水平呈正相关[23]。由此预测,SbZEP的表达水平可能受ABA 调控影响。
SbZEP基因的DNA 变异分析表明,SbZEP在多个位点存在不同的突变,不同突变位点引起氨基酸的改变,这种突变可能与类胡萝卜素的合成及代谢相关。玉米黄质环氧化酶不仅是玉米黄质变异的主要基因,也是叶黄素和β-胡萝卜素变异的主要基因,类胡萝卜素为单基因及多基因变异[27]。拟南芥和高粱野生型OR 中单个氨基酸的改变大大增强了其促进类胡萝卜素积累的能力[28]。因此,推测高粱不同突变位点对类胡萝卜素的合成产生不同作用,类胡萝卜素含量存在遗传多样性。
高粱ZEP基因表达分析显示,高粱在茎、胚珠表达水平较高,结合热图分析可以看出,与SCHWARZ 等[26]和CRUET-BURGOS 等[29]的研究结果具有较高的一致性。SCHWARZ 等[26]通过对AtZEP 蛋白寡肽的特异性抗体进行蛋白质印迹分析,研究了AtZEP 蛋白在野生型植物不同组织中的分布,在叶片中的蛋白质含量最高,其次是茎,根和种子中含量略低。类胡萝卜素生物合成和降解途径的大多数候选基因在高粱种子发育阶段之间存在差异表达。本研究发现,在不同高粱品系中,SbZEP基因在种子内表达水平不同。AtZEP基因表达在种子发育过程中受到发育调节,mRNA 水平在种子发育的后期强烈增加,然后在成熟后的干燥种子内降低[30]。在高粱SSA1和R111品系中,SbZEP1的表达水平20 d 种子高于10 d,可能是种子发育过程中受发育调节引发的结果。
本研究中确定的SbZEP基因与类胡萝卜素含量显著关联,可用于生物强化工作,以提高高粱的营养质量。以上研究成果为玉米黄质环氧化酶基因功能研究提供了基础理论,也为研究该基因在叶黄素循环中的作用机制提供了相关理论成果。
猜你喜欢
古今农业(2022年2期)2022-08-15
南方水产科学(2022年2期)2022-04-22
青年文学家(2022年1期)2022-03-11
今日农业(2021年20期)2021-11-26
食品工业(2021年11期)2021-11-25
食品研究与开发(2016年16期)2016-04-10
中国果菜(2016年9期)2016-03-01
食品科学(2013年22期)2013-03-11
食品科学(2013年19期)2013-03-11
食品科学(2013年13期)2013-03-11
