
西北早粳稻核心种质资源蛋白质含量全基因组关联分析
2023-09-24 07:40:26孙志勇杨小丽陈思怡李培富
种子 2023年7期
马 伟, 孙志勇, 刘 阳, 徐 强, 杨小丽, 陈思怡, 李培富
(宁夏大学农学院/宁夏优势特色作物现代分子育种重点实验室, 银川 750021)
随着生活质量的提高,人们对稻米品质的要求也越来越高,亟需培育出更加优质的水稻新品种满足市场需求。蛋白质作为稻米中含量仅次于淀粉的第二大干物质,一般含量在8%左右,对稻米的外观品质、加工品质和食味品质都有很大影响[1]。因此,通过分子辅助标记方法挖掘出与稻米蛋白质含量相关的基因,对稻米品质改良具有重要意义。
分子辅助标记方法在植物分子育种中被广泛应用。奉杰等[2]以玉米骨干自交系ZNC442和SCML0849为亲本构建的131份F2∶3家系为材料,基于简化基因组测序方法对该群体进行基因型鉴定,利用ICIM软件的完备区间作图法对粒长、粒宽、百粒重等性状进行QTL定位,定位到23个与粒长相关QTLs,29个与粒宽相关QTLs,20个与百粒重相关QTLs。彭强等[3]以V20B/CPSLO17遗传背景衍生的150份重组自交家系作为作图群体,在3种环境下分别进行糙米粒重性状QTL检测及其遗传效应分析,定位到6个与糙米粒重相关的QTLs。
目前,已克隆多个与蛋白质含量相关的基因。OsGS2为谷氨酰胺合成酶基因,Singh等[4]研究发现,OsGS2和OsGS1;1的表达可以提高水稻植株的耐旱性。Lgc-1为低谷蛋白含量基因,Kusaba等[5]研究发现,Lgc1通过RNA沉默抑制glutelin基因表达来降低稻米中谷蛋白含量。OsHT1为调控赖氨酸和组氨酸转运蛋白合成的基因,Guo等[6]研究发现,OsLHT1在营养器官向生殖器官的氨基酸转运过程中对籽粒产量、营养品质和功能品质具有重要作用。Wakasa等[7]研究发现,OsBip1是参与内质网腔中种子贮藏蛋白等分泌蛋白折叠的关键伴侣蛋白的调控基因,OsBiP1的严重抑制或过表达不仅改变了种子表型和胚乳细胞的胞内结构,而且降低了种子贮藏蛋白含量、淀粉积累和粒重。Fang等[8]研究发现,LNUE1编码丙氨酸转氨酶OsAlaAT1基因的合成,调控水稻的氮素利用率、产量和稻米品质。OsGluA2为谷蛋白A2前体蛋白基因,Yang等[9]研究发现,OsGluA2通过增加籽粒存储蛋白含量和氨基酸总量,正调控籽粒的蛋白含量,导致蛋白体Ⅱ的数量和体积都升高,提高了稻米的营养品质。因此,通过GWAS(Genome Wide Association Study)发掘与蛋白质含量相关的基因,研究其表达机制,对水稻分子育种具有重要意义。
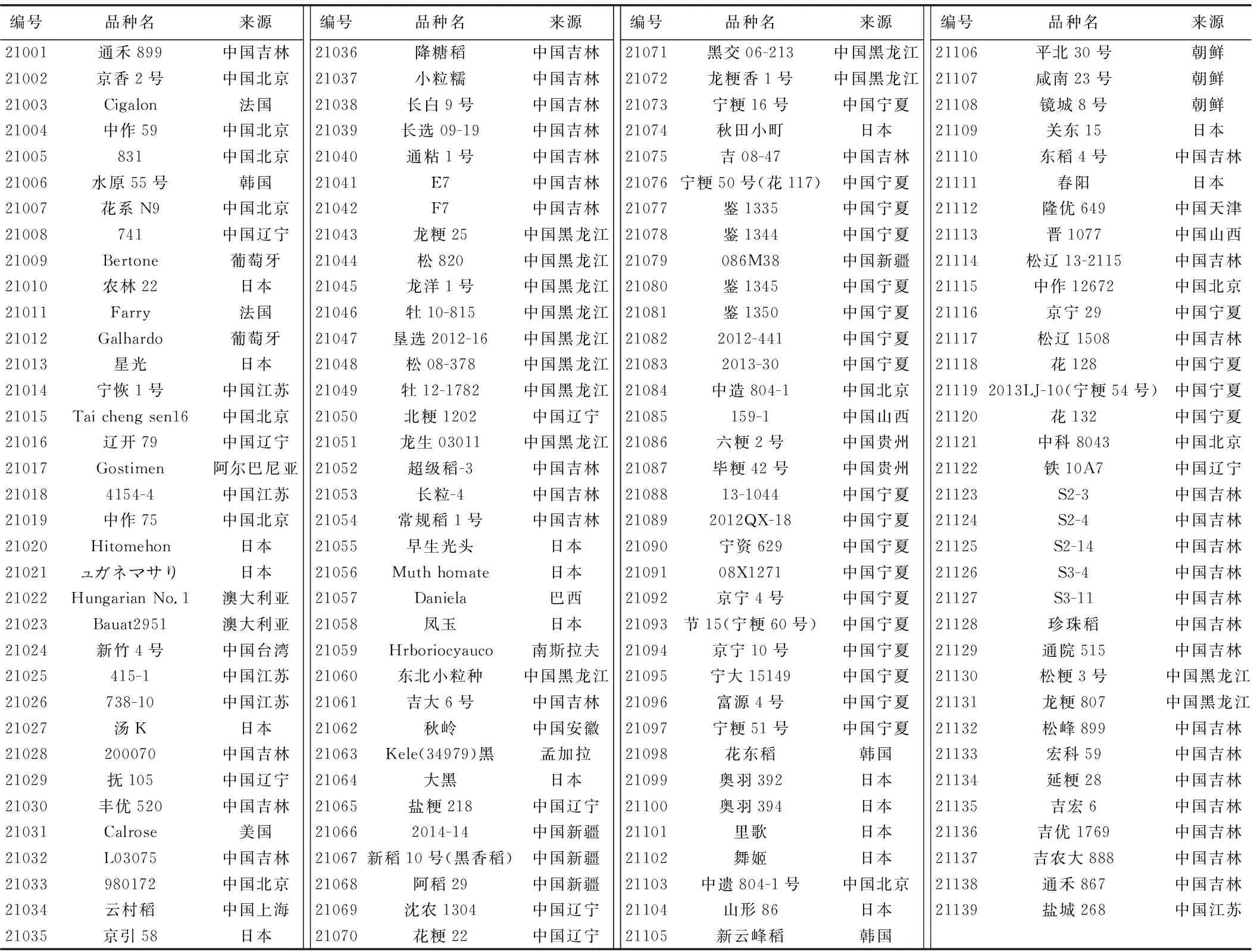
表1 参试水稻种质名称及来源信息Table 1 Name and source information of tested rice germplasm
1 材料与方法
1.1 试验材料
本试验以139份西北早粳稻核心种质资源为材料,2021年于海南种植,每份材料种植两行,单株移栽,每行10株,行长1.2 m,行距0.3 m,株距0.1 m,进行常规田间管理。
1.2 试验方法
1.2.1表型测定
采用北京东孚久恒JDMZ100稻米出米率检测仪检测出米率、LTJM-2099精米机加工稻谷得到精米,采用北京东孚久恒JSWL200大米食味计测定稻米蛋白质含量,3次重复。
1.2.2SNP标记方法
对139份水稻种质资源进行DNA提取和质控,建库测序,测序深度为10×,用基因组比对软件BWA将所有139份水稻过滤后的Clean Reads比对到日本晴参考基因组上,通过突变分析软件GATK检测全基因组中所有的潜在多态性SNP(Single Nucleotide Polymorphism)位点,再根据最大缺失率(0.05)、最小等位基因频率(0.01)等条件进一步过滤,筛选出具有高可信度的SNP位点。
1.3 数据处理
使用Origin2021b软件对139份水稻种质资源的蛋白质含量进行表型数据分析。
2 结果分析
2.1 描述性统计
对139份种质资源的蛋白质含量进行描述性统计,结果表明,139份种质资源蛋白质含量均值为10%,变异系数为16.86%,范围在7%~17%之间。
2.2 方差分析
对139份种质资源的蛋白质含量进行方差分析,结果(表2)表明,在0.05水平下,不同品种间的蛋白质含量有显著差异。

表2 139份种质资源蛋白质含量方差分析Table 2 Variance analysis of protein content of 139 germplasm resources
2.3 频率分布
使用Origin 2021b软件分析139份种质资源蛋白质含量频率分布,作频率分布直方图(图1),结果表明,139份种质资源的蛋白质含量主要分布在8%~12%之间。其中,蛋白质含量在9%~10%之间的有39个品种,蛋白质含量在10%~11%之间的有37个品种。

注:从上到下,从左到右,依次为FarmCPU曼哈顿图、QQ图,GLM曼哈顿图、QQ图,MLM曼哈顿图,QQ图。图2 FarmCPU、GLM、MLM模型Fig.2 FarmCPU,GLM,MLM model
2.4 全基因组关联分析
使用R包MVP进行全基因组关联分析[10],包含了三种模型:FarmCPU、一般线性模型(GLM)和混合线性模型(MLM)。将亲缘关系矩阵和主成分作为协变量加入模型进行校正,减少亲缘关系、群体结构的影响。全基因组水平显著阈值设置为1/markers数[11],对稻米蛋白质含量进行GWAS分析。设置阈值-log10(P)为5.7,筛选与稻米蛋白质含量显著相关的SNPs位点。
本试验共定位到3个与蛋白质含量显著相关的SNPs位点(表3),分布在1号、3号、6号染色体上,-log10(P)在5.79~6.36之间,贡献率在15%~17%之间。其中1号染色体上的位点在LOC_Os01g19600(11105219~11108548)上游,该基因是一个转座子蛋白基因;3号染色体上的位点在LOC_Os03g28120(16163989~16167050)下游,该基因是一个钾离子通道蛋白基因;6号染色体上的位点在LOC_Os06g48440(29302715~29312084)上,该基因是一个反转录转座子蛋白基因。

表3 全基因组关联分析显著SNP位点Table 3 Significant SNP loci in genome-wide association analysis
2.5 候选基因预测
根据全基因组水平显著阈值筛选出与表型显著相关的位点,LD(连锁不平衡),将位点向上下游延伸50 kb的范围定义为候选区间,挖掘候选基因。由表4可知,共关联到32个与蛋白质含量相关的基因,分布在1号、3号、6号染色体上。其中包括1个已克隆的基因[OsCMO(LOC_Os06g48510)];31个未克隆的基因,包括17个反转录转座子蛋白基因,3个转座子蛋白基因,3个三角状五肽重复结构域基因,Kelch基序家族蛋白基因(LOC_Os01g19480),锌离子结合蛋白基因(LOC_Os03g28140),钾离子通道蛋白基因(LOC_Os03g28120),果胶酯酶基因(LOC_Os03g28090),Ring-H2锌指蛋白基因(LOC_Os03g28080),OsFBX94-F-box构域蛋白基因(LOC_Os03g28130),RPM1抗病蛋白基因(LOC_Os06g48520)等,这些候选基因的功能有待进一步验证。
3 结果与讨论
本试验通过对139份核心种质资源的蛋白质含量表型数据进行全基因组关联分析,共定位到3个与蛋白质含量显著相关的SNP位点,关联到32个候选基因。关联到1个已克隆的基因OsCMO(LOC_Os06g48510),该基因是一个胆碱单加氧酶基因,对水稻耐盐性至关重要,过表达OsCMO会增加水稻对盐胁迫的耐受性,转基因株系中甜菜碱含量增加[12]。潘丽娟等[13-14]研究发现,OxCMO基因在水稻幼苗组织中的表达受高盐、低温和干旱等胁迫的上调诱导表达,表明该基因可能在抵御非生物胁迫中发挥重要作用。关联到31个未克隆的基因,包括17个反转录转座子蛋白基因,3个转座子蛋白基因,3个三角状五肽重复结构域基因,Kelch基序家族蛋白基因,锌离子结合蛋白基因,钾离子通道蛋白基因,果胶酯酶基因,Ring-H2锌指蛋白基因,OsFBX94-F-box结构域蛋白基因,RPM1抗病蛋白基因等。转座子对基因的表达和功能有重要影响,林悦龙等[15]研究发现,反转录转座子是许多植物基因组中的重要成分,反转录转座子的复制和移动可以作为沉默子或终止子让插入区域的正常基因发生表达沉默或者翻译提前终止,进而使基因功能失活;也可以直接作为邻近基因的启动子、增强子或者通过改变基因的剪接模式以及产生甲基化等来调控相应基因的表达与功能行使。何虎翼等[16]研究发现,转座子是植物产生变异的重要来源,不同的转座子通过不同的转座机制插入宿主基因组,影响基因表达和功能。Wang等[17]研究发现,五肽重复序列蛋白参与叶绿体或线粒体基因的转录后调控,包括RNA成熟,编辑,内含子剪接,转录物稳定和翻译起始,对植物的光合作用,呼吸作用和发育及其环境响应具有深远的影响。Yousefi等[18]研究发现,锌离子结合蛋白参与调控植物的高温、干旱和盐胁迫。Haque等[19]研究发现,钾离子通道蛋白基因过表达植株在盐胁迫下始终表现出生长优势。Nie[20]研究发现,RPM1抗病蛋白在白粉病病原体感染的先天免疫机制中起重要作用。综上所述,本试验关联到的32个与蛋白质含量相关的基因,对水稻的生长发育有重要影响,为今后稻米品质改良提供参考信息。
猜你喜欢
今日农业(2022年13期)2022-09-15 01:18:00
美食(2022年5期)2022-05-07 22:27:35
林业科学(2022年1期)2022-03-23 06:56:24
少儿科学周刊·儿童版(2021年21期)2021-12-11 01:45:00
中国蜂业(2021年5期)2021-05-22 02:59:26
中国粮食经济(2018年5期)2018-12-27 08:57:26
中国麻业科学(2018年6期)2018-04-09 11:22:12
浙江农林大学学报(2016年6期)2016-12-12 12:01:32
西南农业学报(2016年5期)2016-05-17 05:42:21
广西林业科学(2016年3期)2016-03-16 05:43:21
