
茶树地方群体种资源叶片表型及生化组分多样性分析
2023-08-30 14:19唐璐李长乐葛悦王璞赵华王明乐王郁郭飞倪德江
茶叶科学 2023年4期
唐璐,李长乐,葛悦,王璞,赵华,王明乐,王郁,郭飞,倪德江
茶树地方群体种资源叶片表型及生化组分多样性分析
唐璐,李长乐,葛悦,王璞,赵华,王明乐,王郁,郭飞*,倪德江
华中农业大学园艺林学学院果蔬园艺作物种质创新与利用全国重点实验室,湖北 武汉 430070
为保护茶树种质资源和促进茶树种质创新,对来自全国12个省份的32份茶树地方群体种资源表型性状及生化组分多样性进行了研究。结果表明,16个表型性状的遗传多样性指数变化范围为0.81~1.94,供试群体表型遗传多样性较高;叶面积、叶形、叶面、芽叶茸毛、叶尖、叶身6个表型性状变异的累计贡献率为71.78%,是造成供试茶树群体表型差异的主要因素;综合两年生化成分测定结果,供试样品咖啡碱含量变异系数(17.95%和14.55%)最大,其次是茶多酚含量变异系数(13.61%和8.11%),游离氨基酸变异系数(5.62%和7.52%)最小;基于生化组分含量测定结果进行聚类分析将供试茶树群体划分为3个类群,分别包含不同类型的种质。从供试茶树地方群体种资源的地理分布范围来看,供试种质生化成分含量呈现出地域差异。
茶树地方群体种;表型性状;生化组分;多样性
茶树[(L.)O. Kuntze],作为一种经济价值极高的叶用作物,被广泛栽培于国内多个省份。近年来,随着茶产业的蓬勃发展,对优质、特异、多元茶树种质资源的需求日益增长,发掘和保护优异茶树种质资源的重要性日益凸显。茶树地方群体种对其所在茶区生态条件和生产模式具有良好的适应性,经过长期的自然或人工选择育种,为茶树品种改良提供了丰富的资源。当前育成的无性系茶树品种多是从茶树地方群体种中采用单株系统选育而来,如龙井43[(L.)O. Kuntze cv.]是从浙江龙井群体种中单株系统选育而来。因此,茶树地方群体种既可作为育种材料,从中筛选出具有特异性状(表型性状特异、经济性状特异)的新品种应用于生产,也是研究不同地域茶树亲缘关系、传播方向、演化规律的重要材料,对其开展遗传多样性研究具有重要意义。
开展茶树遗传多样性研究是保护、开发、利用茶树种质资源的前提,是茶树创新育种和基因遗传改良的基础[1]。对茶树表型性状进行测量、统计是最早的遗传多样性分析方法,该方法具有直观、操作简单、成本低的优势,从表型上便可直接选择到与目标性状相关联的个体或群体[2]。茶树作为叶用经济作物,其叶片性状观测在茶树种质资源鉴定中具有重要地位[3]。植物在逆境胁迫下,出于自我保护会启动自身的抗逆机制来适应环境变化,长期的逆境胁迫会产生不可逆的植物表型变异,基于表型的遗传多样性研究对探究植物对环境的适应性及其进化方向都具有重要意义[4]。以植物主要代谢产物差异为基础进行遗传多样性研究的方法[5]有助于指导茶树育种,培育特异种质,运用的分析方法主要有光谱分析法和色谱分析法。茶多酚、咖啡碱、游离氨基酸是茶叶中主要的次生代谢产物,这些生化组分的含量与茶叶品质息息相关,各物质之间的比例差异使不同茶树品种适制性不同。刘彤[6]、莫岚等[7]分别对广西省内柳州及来宾野生茶树群体的主要生化特征进行了鉴定和遗传多样性分析,通过聚类分析,揭示了不同茶树群体的适制性和品质特征。对群体种资源的研究有利于发掘生化成分特异的植株。有学者从广东省茶树群体种中发现了具有高茶多酚、高氨基酸及高儿茶素含量的群体[8-9];对四川地区茶树资源的生化组分表型多样性研究发现,该地区茶树资源的水浸出物含量较高[10];在对广西省不同区域茶树群体种的研究中也发现桂南区域种质的茶多酚、水浸出物及咖啡碱含量较高,桂北区域种质的氨基酸含量较高[11]。因此,通过研究不同茶树品种间叶片表型与生化组分的差异,能从不同角度阐释茶树的遗传多样性水平[12]。
目前茶树种质资源方面的研究主要集中于某一区域(省份),对不同区域(省份)间茶树群体种资源的综合比较研究较少,不利于直观探究不同地域茶树群体种之间的相关性与差异性[13]。本研究搜集了来自全国12个省份的32份茶树群体品种资源,对其新梢芽叶、成熟叶片表型数据进行测量统计,采用主成分分析和聚类分析法对叶片表型性状的多样性展开研究,揭示了不同地区茶树群体种可能的遗传背景;测定样品的主要生化成分游离氨基酸、咖啡碱、茶多酚含量,并进行遗传多样性分析及聚类分析,揭示了不同地区茶树种质资源生化成分含量规律。基于表型性状分析和生化组分含量的分析开展茶树群体种资源遗传多样性研究,以期深入了解不同类型的茶树地方群体种,明确国内群体种质的区域差异与遗传结构,为其演化过程中茶树表型性状、生化成分含量的变化研究提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为在全国12个省份收集的32份茶树地方群体种资源,均为当地具有一定影响力的群体种资源。供试地方群体种茶树经实地考察、多人鉴定后于2015年统一收集种子并播种在营养钵内,随后于2016年3月定植于华中农业大学茶学试验基地茶园种质资源圃。供试茶树种质的名称、来源、编号等信息见表1。试验基地位于东经114°35′,北纬30°47′,海拔29.5 m,年平均温度17.9 ℃,年均降雨量2 012 mm,相对湿度70.3%,属亚热带季风气候。茶园种质资源圃行距1 m,地势平缓,立地条件和栽培管理措施基本一致。
1.2 茶树叶片表型性状观测方法
参照陈亮等[14]编著的《茶树种质资源描述规范和数据标准》,对32份供试群体种茶树的新梢芽叶及成熟叶片表型性状进行观测、统计。从每份群体种资源中随机选取10株作为观测材料,共计观测320株茶树种质。性状观测由2人同时进行,对茶树的芽叶色泽、芽叶茸毛、叶片着生状态、叶色、叶身、叶质等16个表型性状进行观测、记录,12个质量性状描述代码及4个数量性状统计方法如表2所示。
1.3 茶树种质生化成分检测方法
生化成分含量测定样品取样采用混样法,分别于2019年4月和2020年7月在供试茶树三分之二植株新梢发育为一芽二叶(平展)期采摘其新梢一芽二叶,为避免采摘样品受种植区域影响,每份种质分3个区域,每个区域内随机混合采摘1份样品,即进行3次生物学重复,采摘下的样品用液氮处理后于–80 ℃超低温冰箱预冷,随后真空冷冻干燥固样。
生化成分检测方法采用紫外吸光光度法,干物质含量测定参照GB/T 8303—2013;游离氨基酸总量测定参照GB/T 8314—2013;咖啡碱含量测定参照GB/T 8312—2013;茶多酚含量测定参照GB/T 8313—2018。每份样品均进行3次技术重复。
1.4 数据处理
根据田间统计结果及质量性状的赋值标准将质量性状数值化,使用Excel 2019整理并计算表型性状的分布频率、Shannon-weaver多样性指数、标准差、变异系数、平均值等;使用数据处理软件IBM SPSS Statistics 26进行主成分分析(PCA)及欧式遗传距离(Euclidean distance)的计算;基于IBM SPSS Statistics 26计算得出的欧式遗传距离矩阵,利用MEGA 7采用非加权组平均法(UPGMA)对供试材料进行聚类分析。其中变异系数CV=SD/MN×100%,SD为标准差,MN为平均值;Shannon-weaver指数H′=−Σln,其中为该性状第个描述值出现的频率;数值型性状经质量化处理后计算Shannon-weaver多样性指数,参照郭荣华等[15]的数值型性状质量化处理方法将每一性状的观测值赋予不同的等级。使用SPSS Statistics 26进行数据处理,使用TB tools绘制热图(Heat map)。
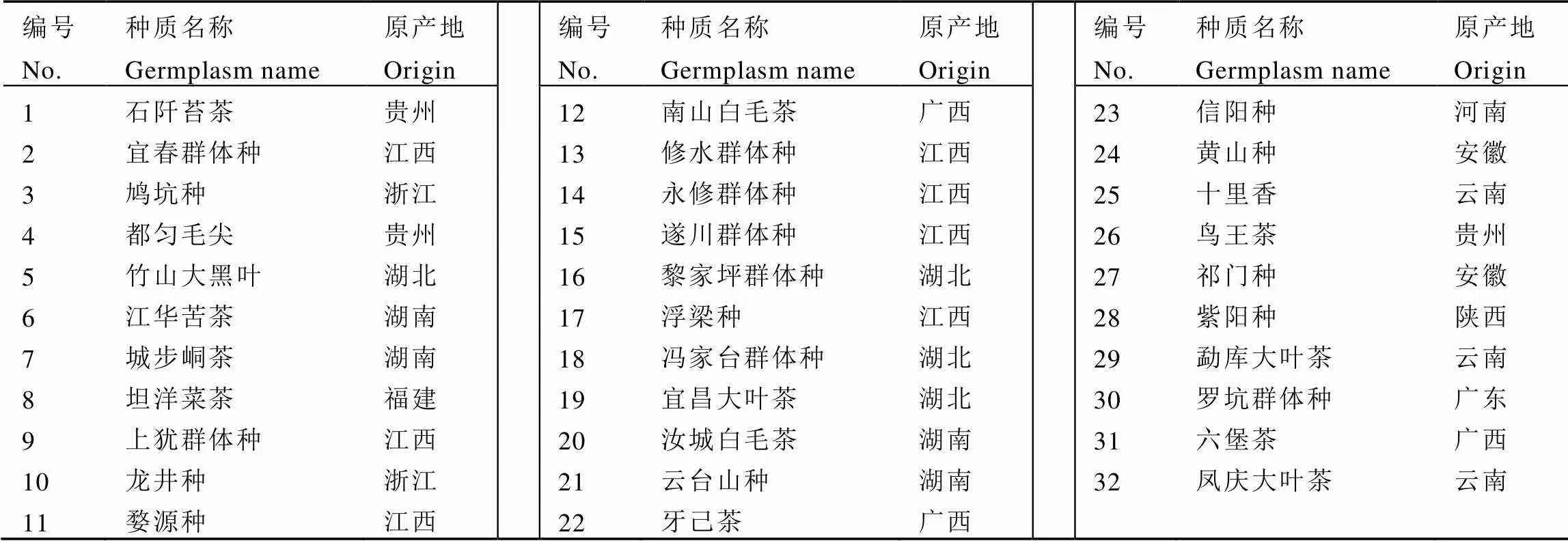
表1 供试材料的名称及原产地
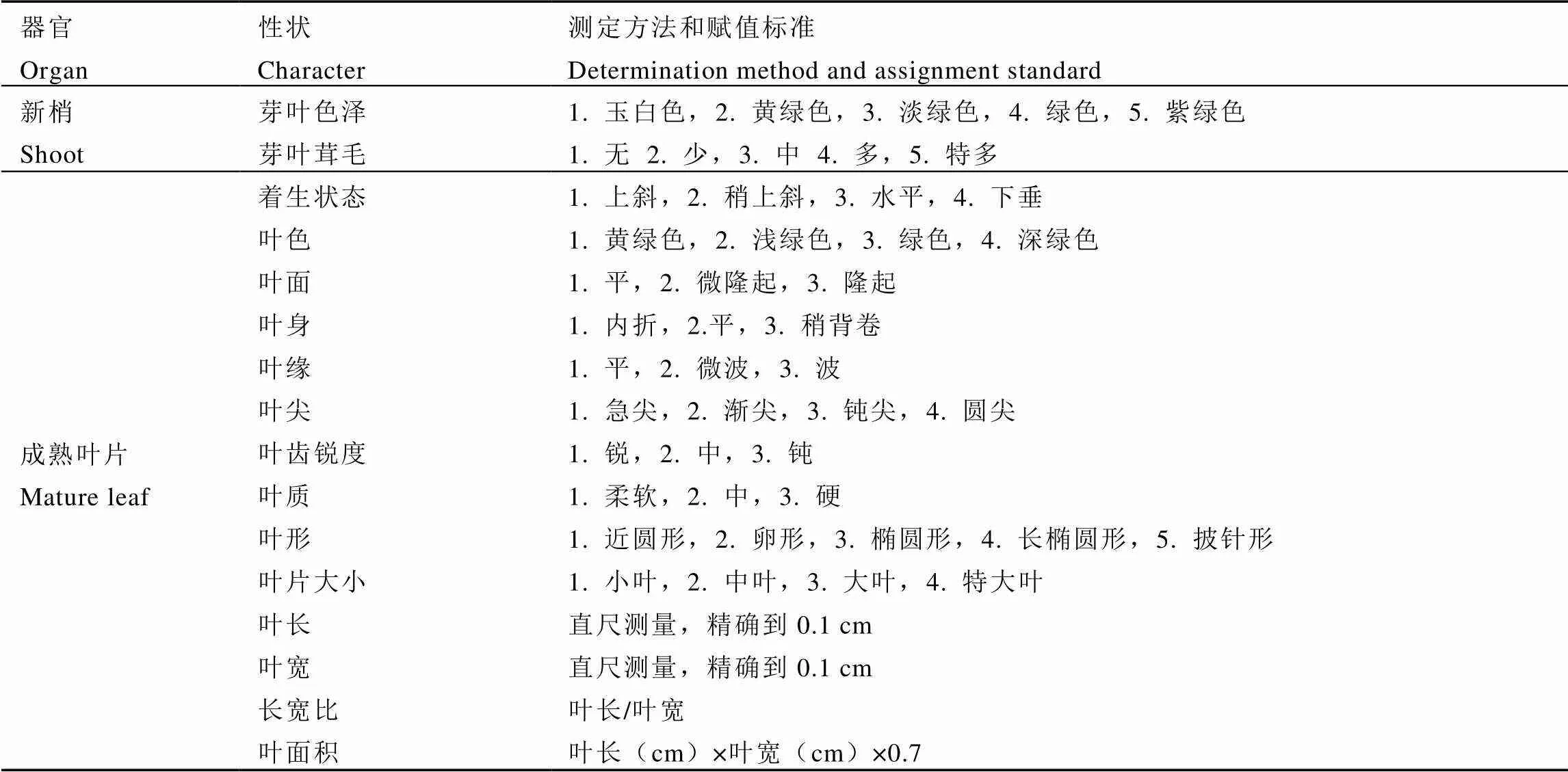
表2 新梢和叶片的质量性状描述代码及数量性状统计方法
2 结果与分析
2.1 茶树叶片表型性状的遗传多样性分析
2.1.1 茶树叶片形态学性状的遗传多样性分析
基于供试茶树表型性状观测结果,对描述型性状分布频率及遗传多样性指数进行统计,结果如表3所示。12个描述型性状中遗传多样性最小的是叶缘(0.81),最大的是芽叶色泽(1.27)。根据多样性指数从高到低排序,依次是芽叶色泽>叶片着生状态>叶形>叶色>叶质>叶身>叶齿>芽叶茸毛>叶面>叶尖>叶片大小>叶缘。供试群体的芽叶色泽以淡绿色为主,占比42.44%;大多数植株芽叶茸毛数量表现为中等,占比55.95%;成熟叶片着生状态以水平着生为主,占44.69%,其次是稍上斜着生,占27.97%,下垂着生占比最少,为8.68%;叶色大多表现为绿色及深绿色,分别占比58.84%和23.15%;叶面主要表现为平和微隆起,分别占比47.59%和44.70%;叶身以平为主,占比54.02%;叶缘微波占比最高,为63.67%,平其次,占30.87%;叶尖主要表现为渐尖,占比67.85%;叶齿锐度多表现为中等锐度,占54.98%;叶质中等硬度的占比最高,为51.77%;叶形以椭圆形居多,占52.09%,其次是长椭圆形,占28.62%;叶片大小以中、小叶为主,其中小叶占比62.06%,中叶占比31.51%。
对供试茶树数值型性状的遗传多样性进行统计,结果见表4。从整体来看,供试茶树的数量性状表现出丰富的多样性:供试材料叶长范围为4.00~18.00 cm,平均为8.12 cm;叶宽的变化范围为1.70~8.60 cm,平均为3.42 cm;长宽比最小值为0.93,最大值为4.00,平均值为2.41;叶面积指数从5.60 cm2到73.71 cm2不等,平均值为20.48 cm2。供试植株各数量性状遗传多样性指数范围为1.77~1.94,皆高于质量性状。变异系数以叶面积(53.33%)最大,其次是叶宽(26.71%),再次为叶长(26.12%),叶片长宽比的变异系数最小,为17.09%。
2.1.2 茶树叶片形态学性状的主成分分析
对供试茶树的表型性状进行主成分分析,得到各主成分特征值变化折线图(图1)。其中横坐标显示的是16个主成分,纵坐标显示的是各主成分对应的特征值,由图可知前6个主成分都包含较大的特征值(特征值>1)、折线较为陡峭,第7个主成分之后折线趋于平缓,说明前6个主成分包含的信息量大。对各主成分的特征值、贡献率、累计贡献率进行统计(表5),结果表明前6个主成分的累计贡献率达71.78%,能较好地概括16个表型性状的大部分信息。其中第一主成分贡献率为25.49%,主要受叶长、叶宽、叶面积和叶片大小的影响,载荷值分别为0.87、0.93、0.96和0.91,表现为高度正相关,其中占主导地位的是叶面积;第二主成分主要受到长宽比和叶形的影响,载荷值为0.83和0.88,贡献率为13.16%,其中占主导地位的是叶形;第三主成分贡献率为10.21%,与叶色、叶面、叶身、叶缘、叶质呈正相关,载荷值分别为0.47、0.55、0.52、0.44、0.48,与叶齿呈负相关,载荷值为–0.43,其中占主导地位的是叶面及叶身;第四主成分与芽叶茸毛高度正相关,载荷值为0.79,与叶缘高度负相关,载荷值为–0.60,贡献率为9.04%;第五主成分主要受叶面的影响,载荷值为0.60,贡献率为7.05%;第六主成分主要受叶尖性状的影响,载荷值为0.51,贡献率为6.83%。综合来看,叶面积、叶形、叶面、芽叶茸毛、叶尖、叶身这6个表型性状可以反映出16个表型性状71.78%的信息,是造成供试茶树群体表型差异的主要因素。
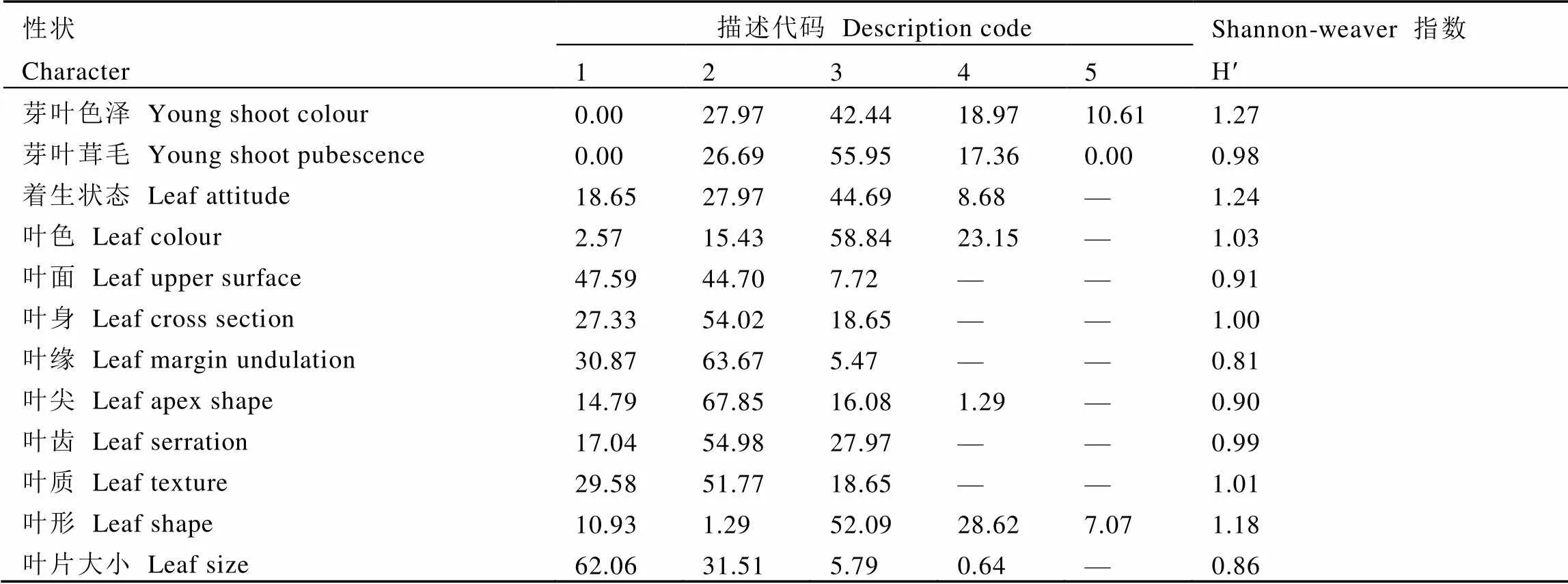
表3 描述型表型性状的分布频率和多样性指数
2.1.3 茶树叶片形态学性状的种质聚类分析
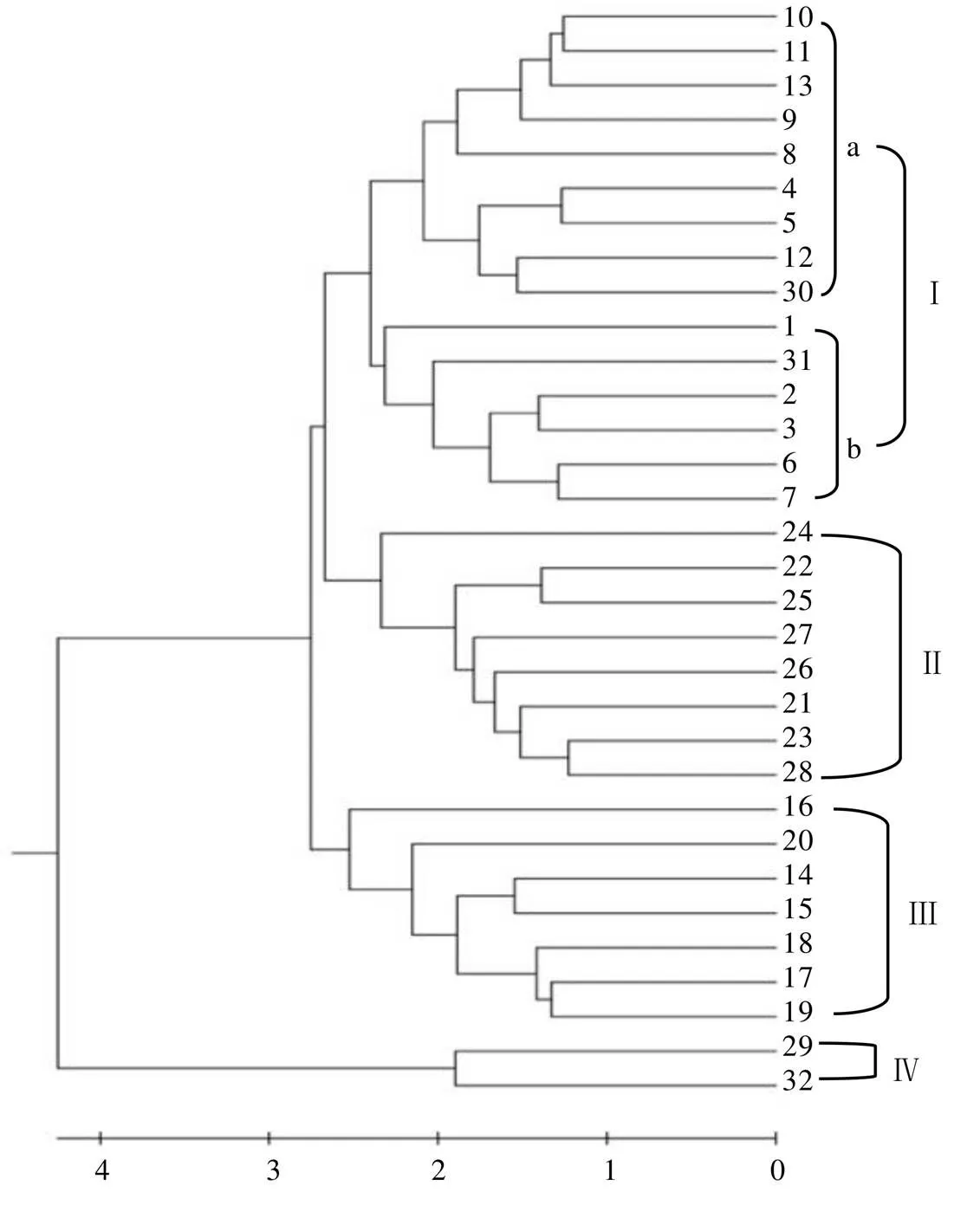
根据供试材料的16个表型性状信息,采用非加权平均法(UPMGA)基于欧式遗传距离对32份茶树群体种质进行聚类分析(图2)。聚类分析结果表明,供试茶树种质被划分为4个类群。类群Ⅰ包括2个亚群,亚群a包含龙井种等9个群体种,亚群b包括石阡苔茶等6个群体种。其主要特征是叶片以中小叶为主、叶质中等、叶缘微波、叶身内折、芽叶茸毛少、芽叶色泽绿。类群Ⅱ包括黄山种等8个群体种,其主要特征是叶片以中小叶为主、芽叶色泽黄绿及淡绿、芽叶茸毛多、叶缘平、叶形椭圆形。类群Ⅲ包括黎家坪群体种等7个群体种,其主要特征是叶片以小叶为主、芽叶茸毛中等、叶片着生状态上斜及稍上斜、叶色绿色、叶质中等偏硬。类群Ⅳ包括勐库大叶茶和凤庆大叶茶,主要特征是大叶种、叶质柔软、叶齿较钝、叶面微隆起、叶色淡绿色、芽叶茸毛中等。
2.2 茶树种质主要生化成分的遗传多样性分析
2.2.1 主要生化成分含量遗传多样性分布
春季样品测定结果显示(图3),供试茶树种质的游离氨基酸含量范围为34~49 mg·g-1(干重),其中氨基酸含量在37~40 mg·g-1的茶树材料有15份(占比46.88%),游离氨基酸含量在>40~43 mg·g-1的茶树材料有8份(占比25.00%);供试茶树材料的咖啡碱含量在20~45 mg·g-1,其中12份茶树材料咖啡碱含量在30~35 mg·g-1(占比37.50%),9份茶树材料咖啡碱含量在>35~40 mg·g-1(占比28.13%);供试茶树材料的茶多酚含量位于212~352 mg·g-1,其中19份茶树材料的茶多酚含量在268~296 mg·g-1(占比59.38%)。
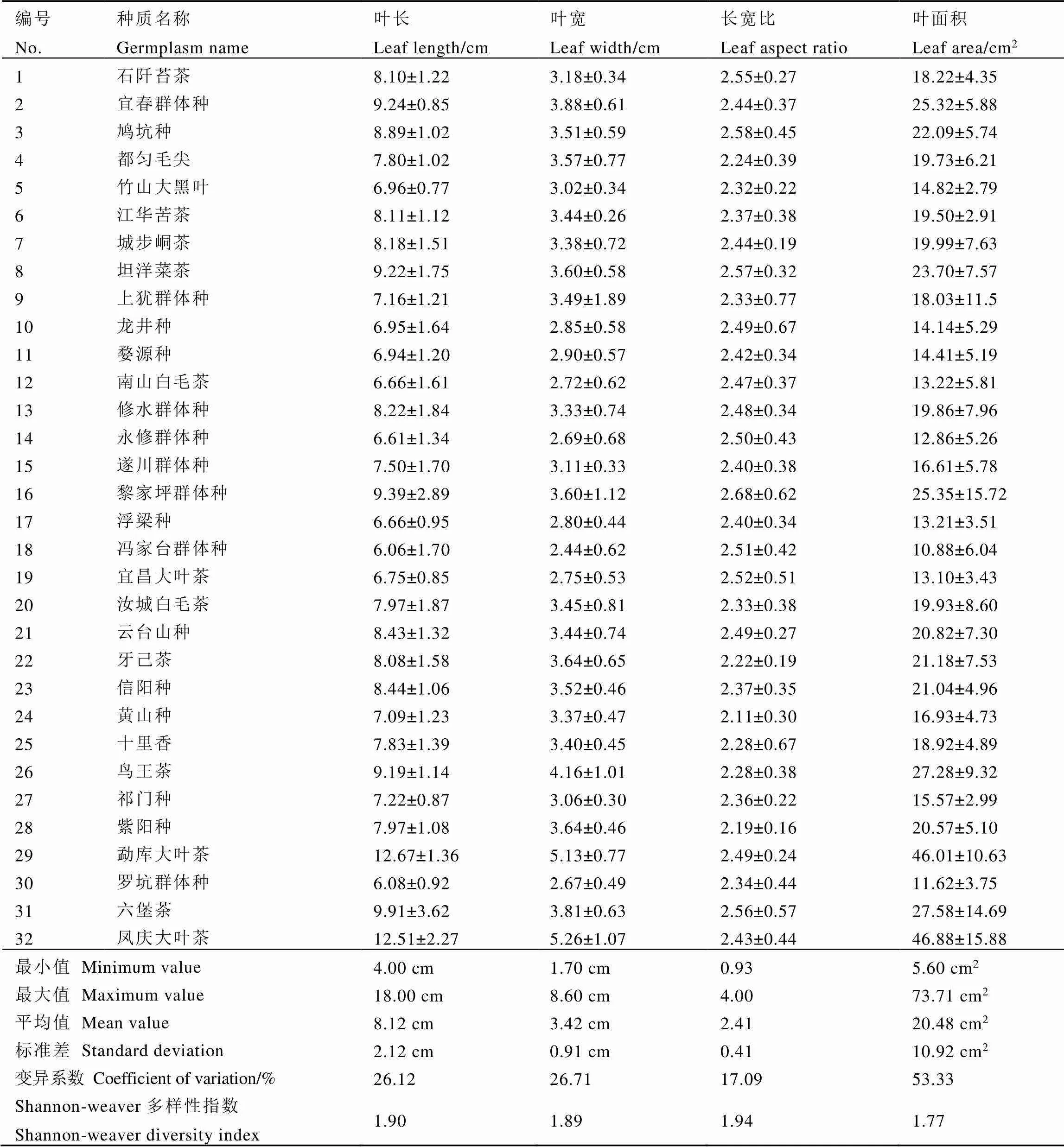
表4 数值型表型性状的变异情况和分布特征
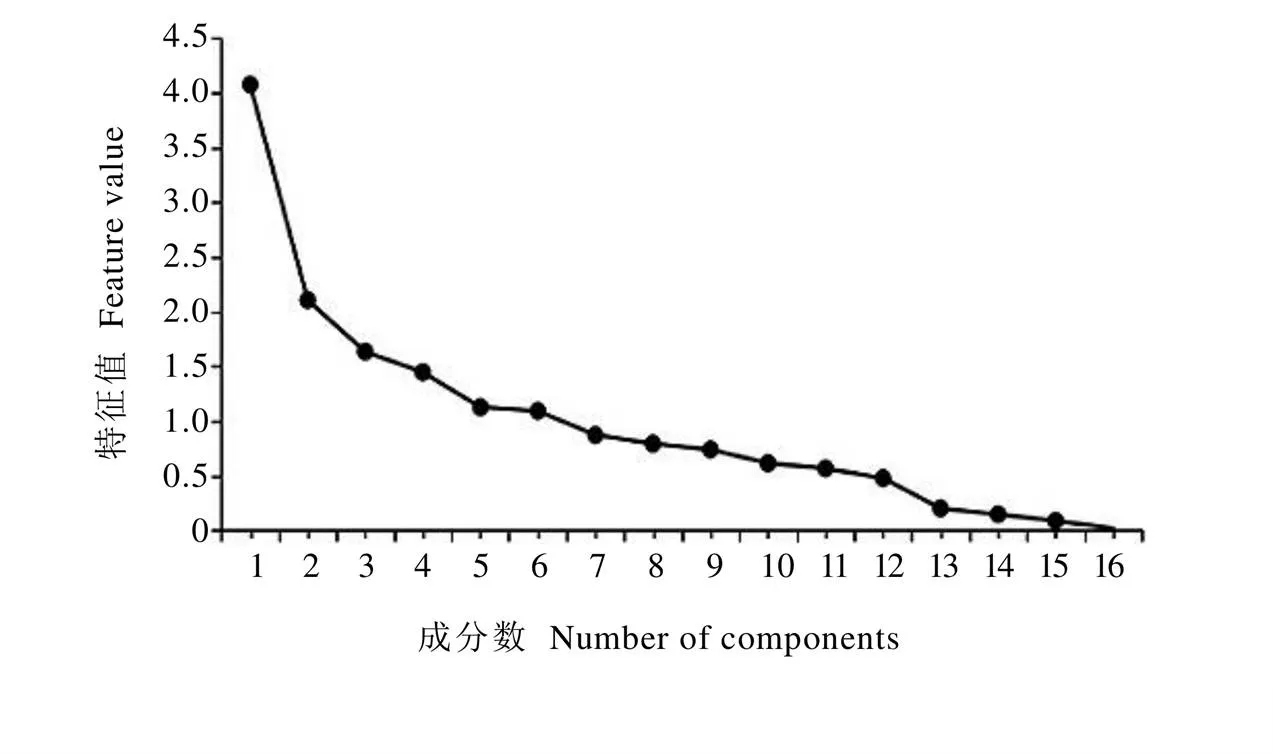
图1 主成分特征值变化折线图
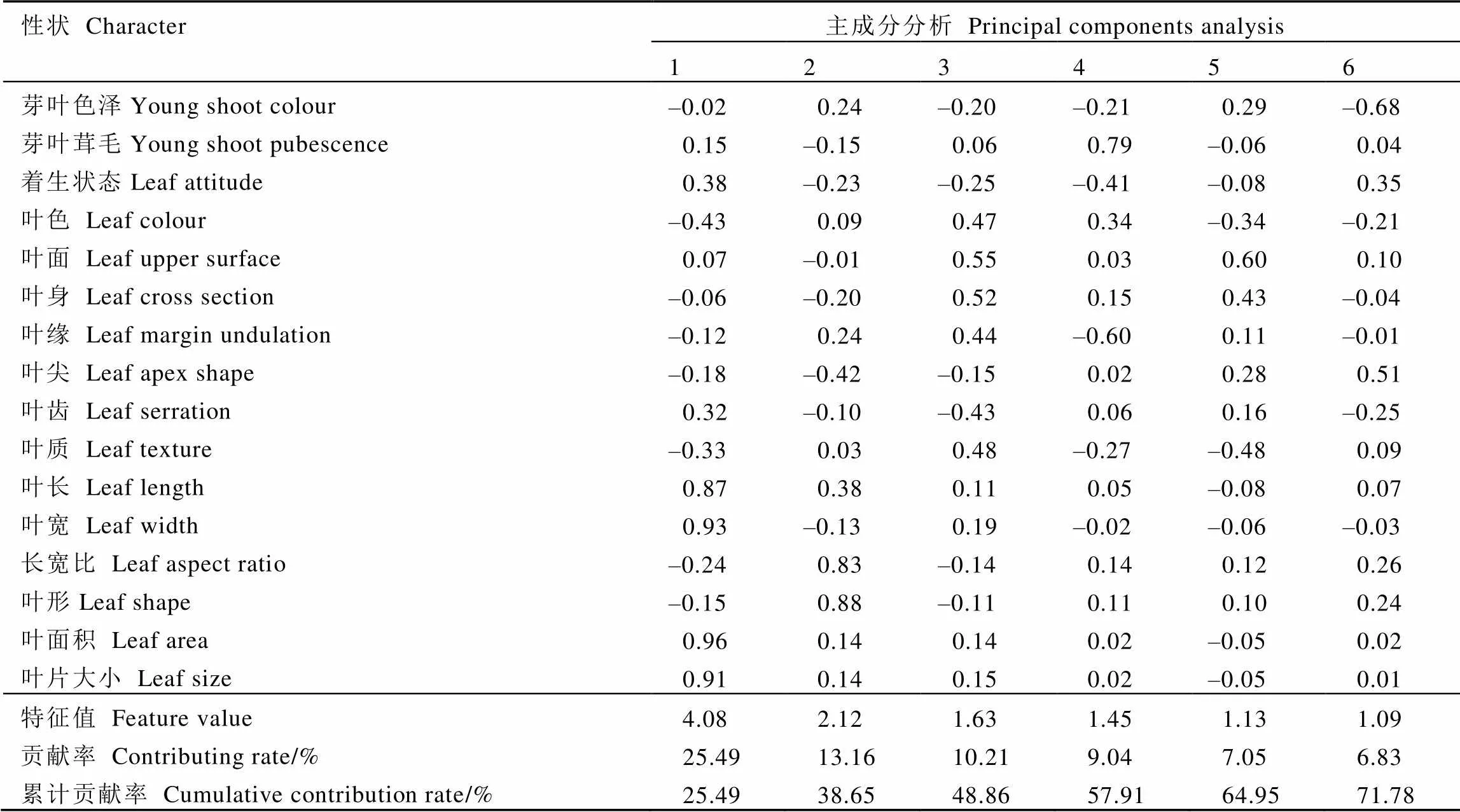
表5 基于16个表型性状的茶树群体种质主成分分析
2.2.2 主要生化成分含量遗传多样性分析
对32份茶树地方群体种资源的两年生化成分含量进行测定、统计,供试材料表现出丰富的遗传多样性,结果见表6。从整体上看,春季,供试材料的游离氨基酸含量平均值为32.28 mg·g-1(干重),变异系数为5.78%,其中鸟王茶含量最低(27.12 mg·g-1),宜春群体种含量最高(36.48 mg·g-1);咖啡碱含量平均值为21.54 mg·g-1,变异系数为18.63%,变化范围12.29~31.19 mg·g-1,含量最低的是鸟王茶,含量最高的是遂川群体种;茶多酚含量平均值为200.40 mg·g-1,变异系数为13.61%,含量最低的是宜春群体种(153.71 mg·g-1),含量最高的是江华苦茶(277.07 mg·g-1)。2020年夏季供试材料的游离氨基酸含量平均值为40.52 mg·g-1,变异系数为7.52%,竹山大黑叶含量最低(36.49 mg·g-1),十里香含量最高,为46.90 mg·g-1;咖啡碱含量平均值为33.81 mg·g-1,变异系数为14.55%,凤庆大叶茶含量最低(22.70 mg·g-1),遂川群体种含量最高,为42.11 mg·g-1);茶多酚含量平均值为282.00 mg·g-1,变异系数为8.11%,变化范围为214.18~349.93 mg·g-1,龙井种含量最低,江华苦茶含量最高。
注:1. 石阡苔茶,2. 宜春群体种,3. 鸠坑种,4. 都匀毛尖,5. 竹山大黑叶,6. 江华苦茶,7. 城步峒茶,8. 坦洋菜茶,9. 上犹群体种,10. 龙井种,11. 婺源种,12. 南山白毛茶,13. 修水群体种,14. 永修群体种,15. 遂川群体种,16. 黎家坪群体种,17. 浮梁种,18. 冯家台群体种,19. 宜昌大叶茶,20. 汝城白毛茶,21. 云台山种,22. 牙己茶,23. 信阳种,24. 黄山种,25. 十里香,26. 鸟王茶,27. 祁门种,28. 紫阳种,29. 勐库大叶茶,30. 罗坑群体种,31. 六堡茶,32. 凤庆大叶茶
综合两季生化成分含量的测定结果对含量特异的茶树种质材料进行挖掘,结果表明,春季样品中宜春群体种的茶多酚含量最低,夏季样品中其茶多酚含量亦低于大部分群体种;江华苦茶在春、夏两季中咖啡碱和茶多酚含量都较高;龙井种的茶多酚含量在春、夏两季中皆相对较低;遂川群体种春、夏两季咖啡碱含量皆为最高;黎家坪群体种多酚含量相对较高,且游离氨基酸含量在春季样品中最高,在夏季样品中亦相对较高;十里香游离氨基酸含量在夏季样品中最高,春季亦较高;鸟王茶在两季节的氨基酸和咖啡碱含量均极低;罗坑群体种在两季节中的氨基酸含量均极高;凤庆大叶茶在两季中的咖啡碱含量均极低。
2.2.3 茶树种质主要生化成分的聚类分析
使用TB tools对夏季茶树地方群体种资源的生化成分含量数据进行归一化处理后,基于欧式遗传距离采用最长距离法(Complete)聚类并绘制热图,结果如图4所示,供试茶树群体种被划分为3个类群。类群Ⅰ包含遂川群体种等9个群体种,其主要特征是氨基酸含量中等偏低,咖啡碱含量较高。类群Ⅱ包含3个亚群,亚群a包括江华苦茶、城步峒茶、黎家坪群体种和祁门种,其主要特征是咖啡碱和茶多酚含量较高,氨基酸含量中等,其中江华苦茶具有极高的茶多酚含量;亚群b包括石阡苔茶等6个群体种,其主要特征是氨基酸含量较低,咖啡碱和茶多酚含量居中;亚群c包括鸟王茶、宜昌大叶茶、云台山种和紫阳种,其主要特征是氨基酸含量偏低,咖啡碱和茶多酚含量低。类群Ⅲ包含凤庆大叶茶等9个群体种,除龙井种外氨基酸含量皆高于其他群体种,凤庆大叶种的咖啡碱含量在供试群体中最低。
从供试种质资源来源省份的地理分布范围来看(表7),来自同一辖区内的茶树种质资源多聚为一类,江西省有5份供试茶树群体种聚在一起,来自湖南、湖北、安徽的全部种质聚在类群Ⅱ中,来自云南、广西的全部种质聚在类群Ⅲ中。贵州省和江西省茶树种质资源表现出丰富的多样性,分别属于两种及以上类群:贵州省(Ⅰ、Ⅱ)、江西省(Ⅰ、Ⅱ、Ⅲ)。
3 讨论
3.1 茶树叶片表型性状的遗传多样性分析
表型性状是区分不同群体种茶树的重要依据[16],本研究对32份供试茶树种质资源的表型性状进行遗传多样性分析,16个表型性状的Shannon-weaver遗传多样性指数变化范围为0.81~1.94。其中12个描述型性状的遗传多样性指数最小值为0.81,最大值为1.27,远高于前人关于鸠坑茶树资源及贵州古茶树芽叶质量性状调查所得的遗传多样性指数[17-18],说明供试材料表型性状变异较为丰富。整体看,供试茶树种质的芽叶色泽以淡绿色为主,芽叶茸毛数量表现为中等,成熟叶片着生状态以水平为主,叶色大多表现为绿色,叶面主要表现为平,叶身以平为主,叶缘大多表现为微波,叶尖主要表现为渐尖,叶齿锐度多表现为中等锐度,叶质主要表现为中等硬度,叶形以椭圆形和长椭圆形为主,叶片大小以中小叶为主,其中小叶占比较多,与周炎花[19]关于国内茶树品种芽叶表型的调查结果较为一致。
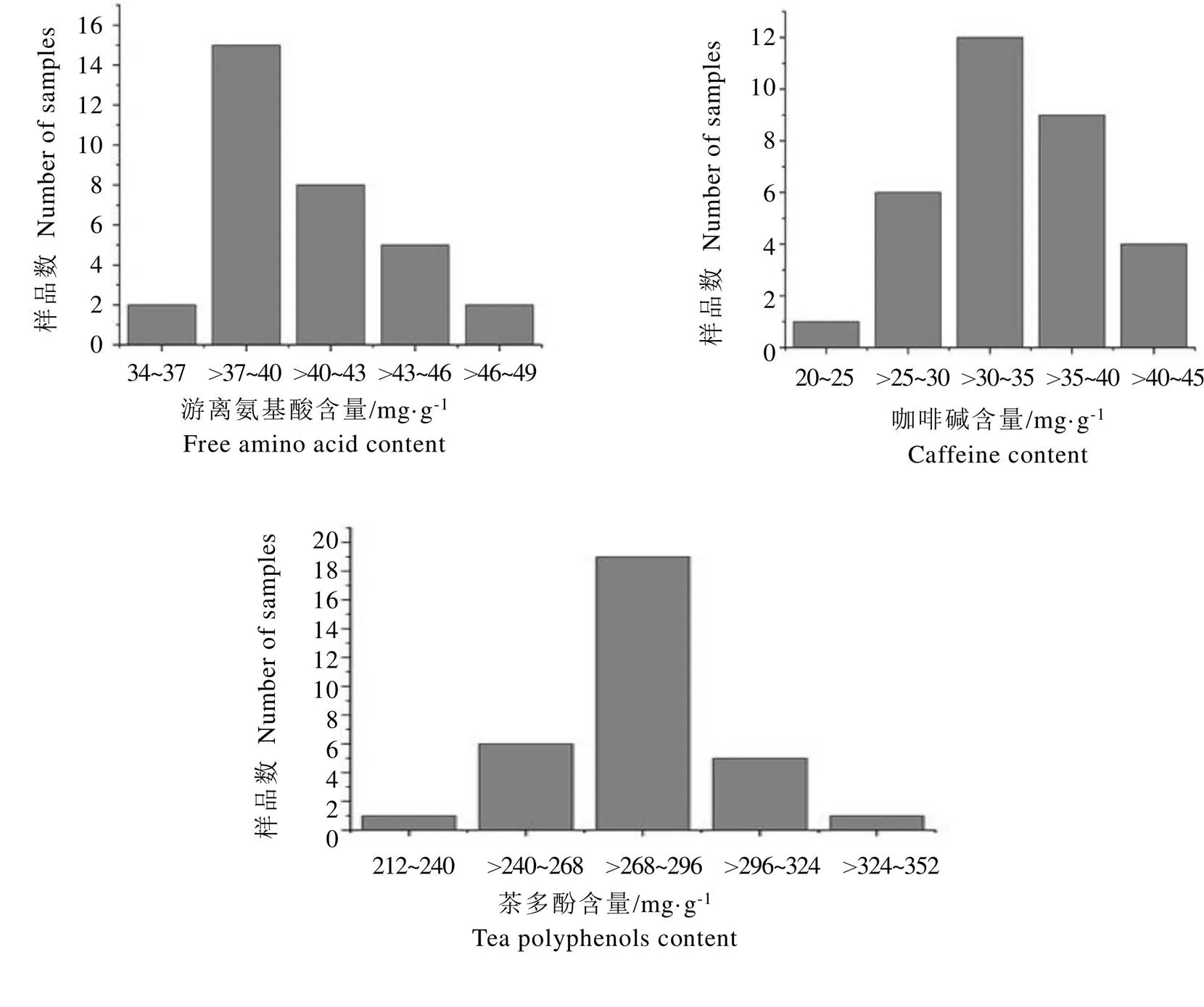
图3 夏季样品主要生化成分含量分布图
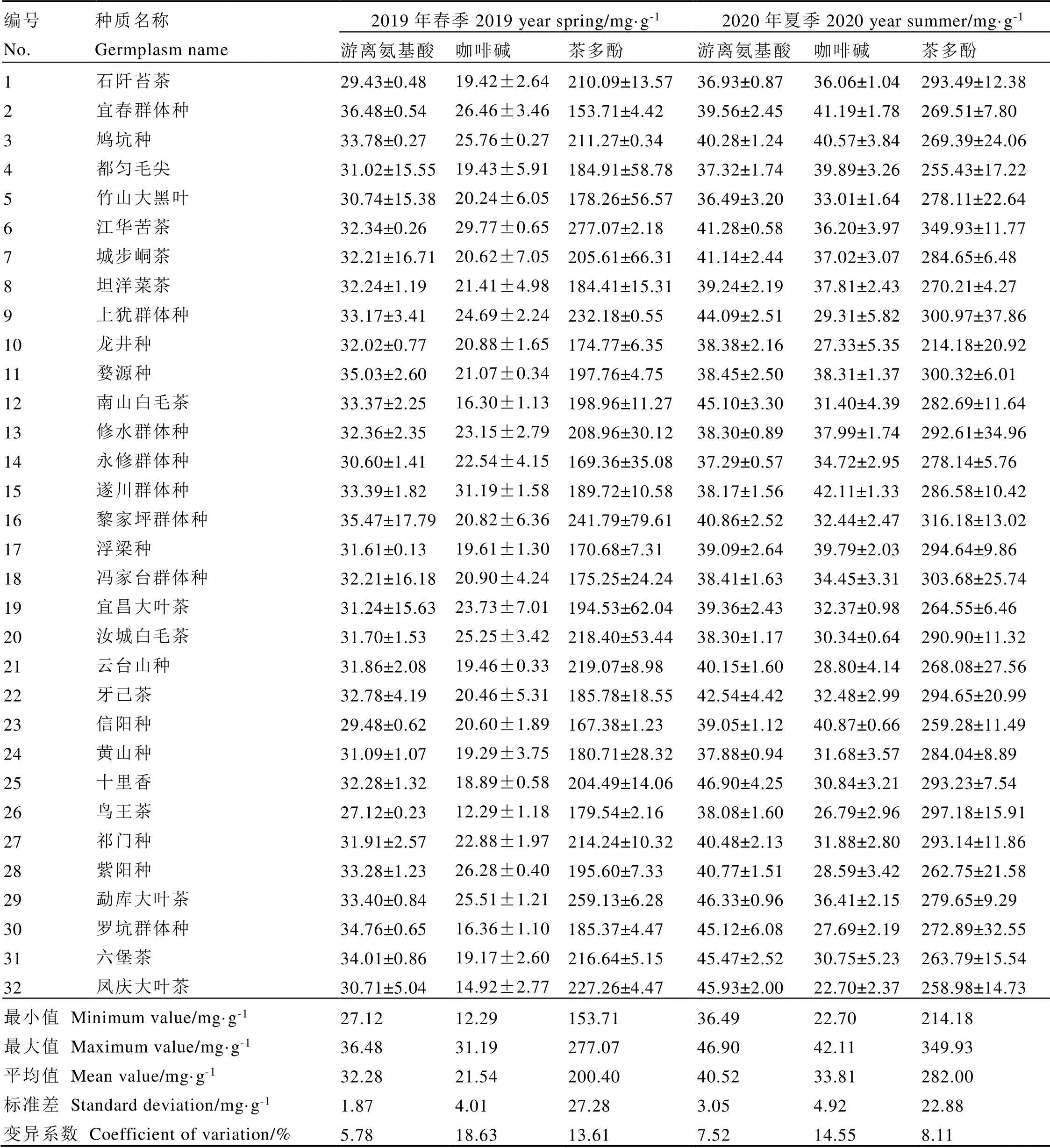
表6 32份种质资源主要生化成分含量
不同种质表型性状主成分分析也可作为种质评价的一种手段,目前已被广泛用于杨梅、猕猴桃、柑橘、苹果等种质资源的评价工作中[20-25],通过计算不同种质的综合得分,可以实现对优异种质资源的快速便捷筛选。茶树16个表型性状的主成分分析及聚类分析结果表明,叶面积、叶形、叶面、芽叶茸毛、叶尖、叶身这6个表型性状是造成供试茶树群体表型差异的主要因素;供试茶树除类群Ⅳ表现为大叶种外,其余3个类群皆表现为中、小叶种,其中类群Ⅲ叶片偏小;类群Ⅱ叶形以椭圆形为主,其余类群叶形多在椭圆、长椭圆间变异;类群Ⅳ叶面多表现为隆起,类群Ⅰ中亚群b多表现为平;芽叶茸毛以类群Ⅰ中亚群b最多,类群Ⅰ中亚群a最少;类群Ⅳ尖多表现为急尖和渐尖,其他类群多表现为渐尖;类群Ⅲ叶身包含平和稍背卷两大类型,类群Ⅰ和类群Ⅳ叶身多表现为内折、平。主成分分析筛选得到的6个性状基本上可以将聚类分析划分的四大类型区分开,且亲缘关系近、地域位置近的材料大多聚在一起。林子翔[26]对观赏辣椒种质进行了主成分分析,挖掘了影响观赏辣椒价值的主要农艺性状,并在此基础上进行聚类分析,39份供试材料被划分为3个类型,各类型之间特征显著、且差异较大,与本研究结果相似。本研究主成分分析中第四主成分与芽叶茸毛高度正相关(载荷值为0.79),与叶缘高度负相关(载荷值为–0.6),推测叶缘与芽叶茸毛存在相关性。
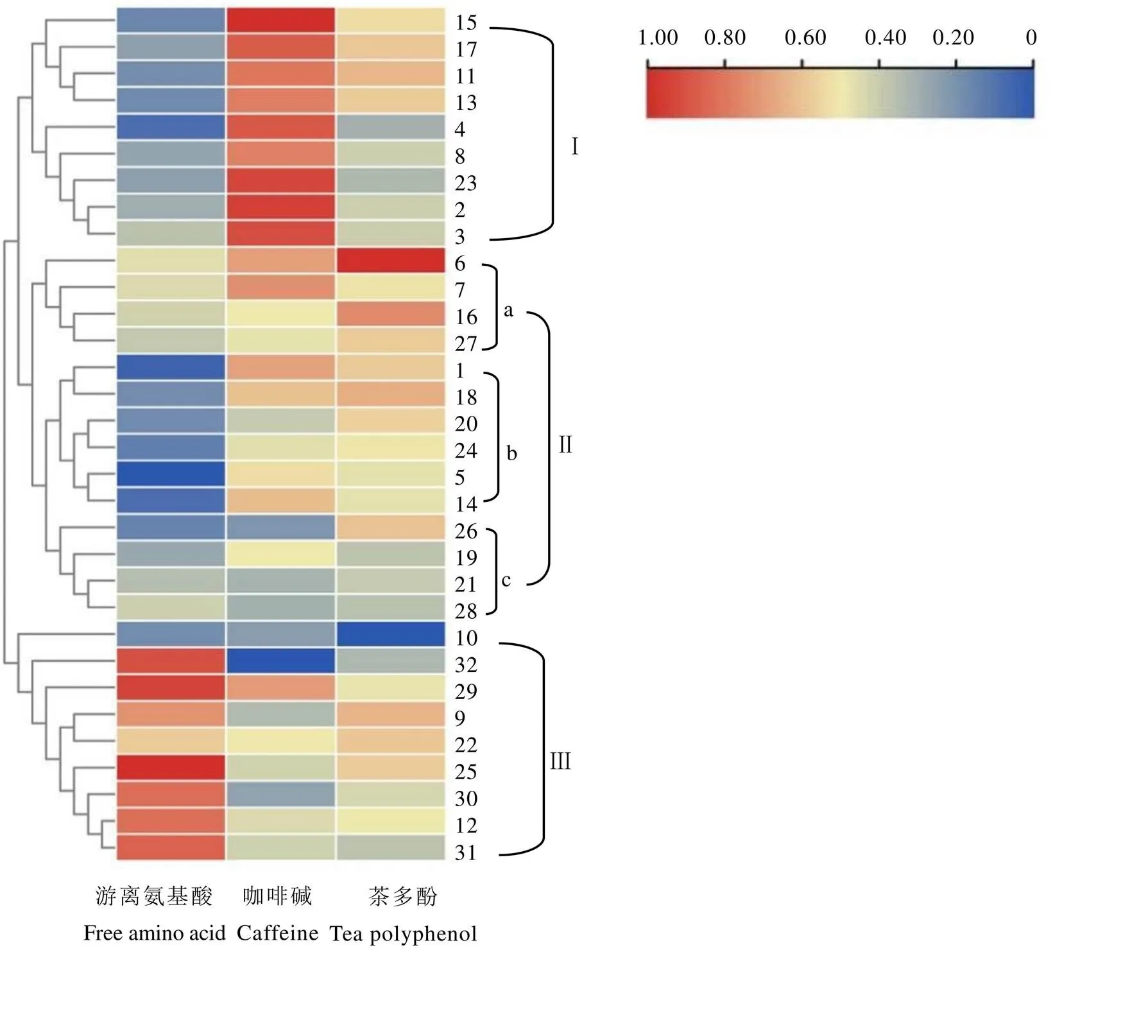
注:1. 石阡苔茶,2. 宜春群体种,3. 鸠坑种,4. 都匀毛尖,5. 竹山大黑叶,6. 江华苦茶,7. 城步峒茶,8. 坦洋菜茶,9. 上犹群体种,10. 龙井种,11. 婺源种,12. 南山白毛茶,13. 修水群体种,14. 永修群体种,15. 遂川群体种,16. 黎家坪群体种,17. 浮梁种,18. 冯家台群体种,19. 宜昌大叶茶,20. 汝城白毛茶,21. 云台山种,22. 牙己茶,23:信阳种,24. 黄山种,25. 十里香,26. 鸟王茶,27. 祁门种,28. 紫阳种,29. 勐库大叶茶,30. 罗坑群体种,31. 六堡茶,32. 凤庆大叶茶。
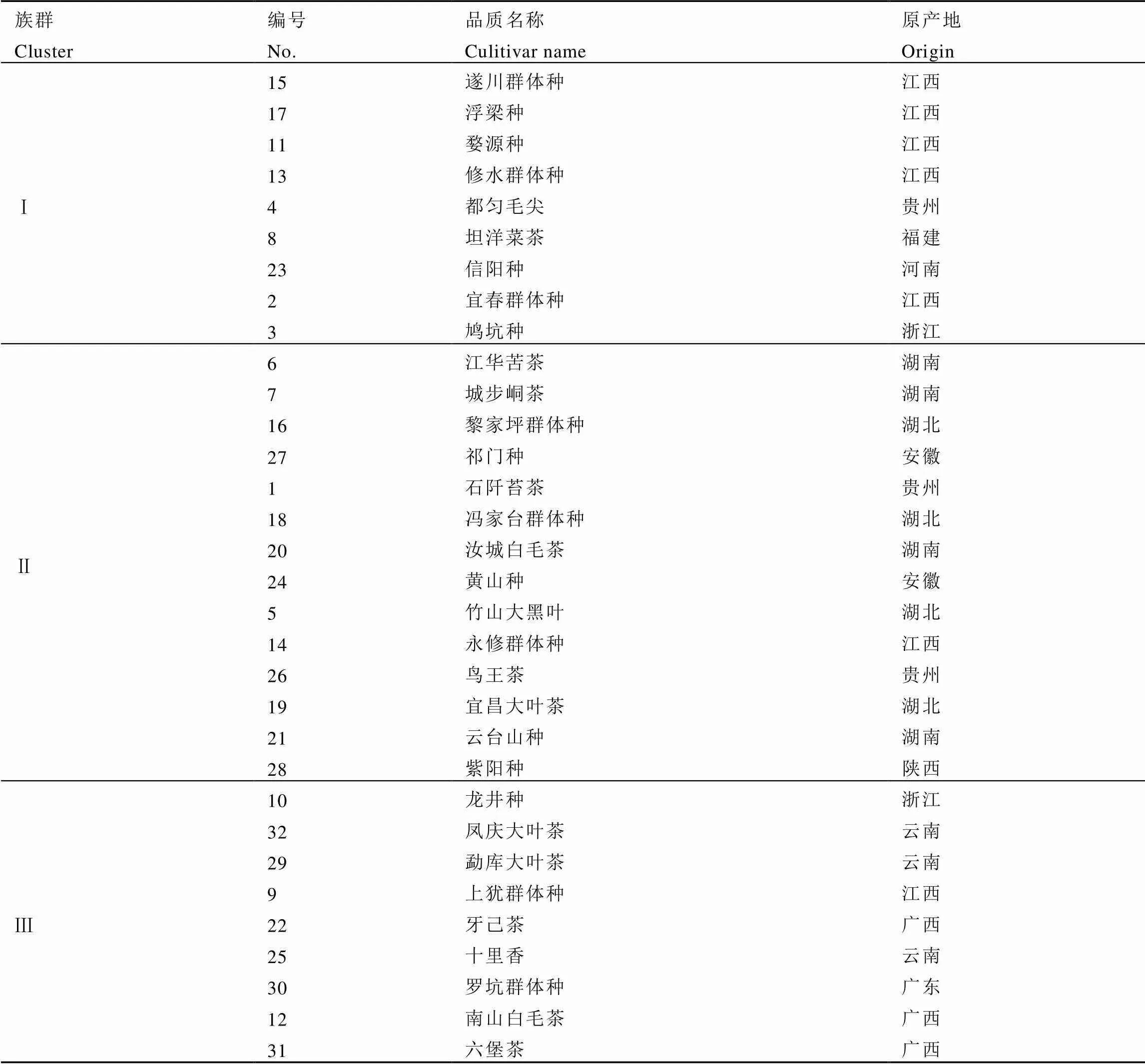
表7 基于主要生化成分含量的32份茶树群体种资源聚类族群
3.2 茶树种质主要生化成分的遗传多样性分析
两年供试样品生化成分测定结果表明,供试茶树种质资源的游离氨基酸、茶多酚、咖啡碱含量总体上符合正态分布,且表现出丰富的遗传多样性,供试样品咖啡碱含量的变异系数(17.95%和14.55%)最大,其次为茶多酚含量的变异系数(13.61%和8.11%),再次为游离氨基酸的变异系数(5.62%和7.52%),说明供试样品中资源类型丰富,可为选育生化成分含量特异(如低咖啡碱含量、低茶多酚含量等)的优良茶树单株提供参考。在立地条件和栽培管理措施基本一致的情况下,供试材料咖啡碱含量的差异较大,这与宁功伟等[27]、董方等[28]、丁帅涛等[29]的研究结果相似,推测原因可能是不同茶树品种间遗传背景差异所导致。咖啡碱合成通路中-甲基转移酶基因()[30]、咖啡碱合成酶基因()[31]等关键酶基因及[32]、[33]等转录因子的表达水平均会影响茶类植物中咖啡碱的含量。Jin等[34]研究表明,低咖啡碱茶树品种中存在两种控制咖啡碱生物合成的分子机制,即低转录水平的等位基因或其编码的仅具有可可碱合成酶活性的蛋白质。除此之外,植物体内的生物碱与其抗逆性有关,具有抵御病害、应对环境胁迫等生理作用,咖啡碱作为茶树体内最主要的生物碱,其含量的差异与不同品种间抗逆性的联系有待进一步研究。综合测定两年的生化含量,结果发现,生化成分含量特异的地方群体种资源有9份:宜春群体种(低茶多酚含量)、江华苦茶(高咖啡碱和茶多酚含量)、龙井种(低茶多酚含量)、遂川群体种(高咖啡碱含量)、黎家坪群体种(高茶多酚含量、高氨基酸含量)、十里香(高氨基酸含量)、鸟王茶(低咖啡碱含量)、罗坑群体种(高氨基酸含量)、凤庆大叶茶(低咖啡碱含量),其中凤庆大叶茶咖啡碱含量极低,且两年测定结果一致,可以作为筛选低咖啡碱含量种质或研究茶树品种苦味改良的育种材料[35]。目前针对特定人群的低咖啡碱产品开发取得了快速进展,但大多数脱咖啡碱工艺会影响产品的风味且增加了额外的加工成本,因此选育低咖啡碱茶树品种具有广阔发展前景[36]。蒋会兵等[37]通过对云南省内51份茶树地方品种进行生化成分测定,筛选出包括凤庆大叶茶在内的4份具有较低咖啡碱含量的茶树地方群体种资源,以及昆明十里香在内的适制绿茶的群体种,与本研究结果一致。对32份供试茶树群体种的生化成分含量测定结果进行聚类分析,供试茶树群体被划分为3类,分别包含不同类型的种质资源:类群Ⅰ的主要特征是咖啡碱含量较高;类群Ⅱ中亚群c主要表现为较低的咖啡碱和茶多酚含量;类群Ⅲ包含具有较高氨基酸含量、较低咖啡碱含量的种质资源。茶树群体种变异类型多但自然变异具有不确定性,对各群体种基于生化成分含量划分类群,了解群体种的共性,可为育种材料及相关代谢研究的材料选择、核心种质的筛选提供科学依据。
从各类型茶树种质资源地理分布范围来看,不同类型的茶树种质资源分布具有明显的地域差异:类群Ⅲ除龙井种外主要分布在广东、广西、云南等茶区,这些茶区位于中国西南及南方;而其他省份种质资源除上犹群体种外皆属于类群Ⅰ、类群Ⅱ。其中来自贵州省、江西省的供试茶树群体种分布于多个类群,说明这些地区包含的茶树群体种质资源丰富。姚明哲[38]、陈勋等[39]基于SSR分子标记技术对茶树种质进行遗传多样性进行了研究,结果亦表明,江西省、贵州省的茶树种质在中国各省份茶树种质资源中表现出中度偏高的遗传多样性水平。另外,随着无性系品种的推广、引进,大多栽植地方群体种的茶园被改植换代,群体种质遭到破坏,若不及时加以保护,会导致物种基因多样性降低、大量种质类型丢失[40]。中国西南地区作为茶树起源的中心地带,江西省作为茶树传播、演化的中间地带,含有丰富的茶树群体种质资源[41],应重点开展这些地区茶树地方群体种表型性状、生化性状、遗传背景、群体结构研究工作,推进茶树群体种质的发掘与保护,加快核心种质库的建立,为茶树遗传育种提供更多宝贵的种质材料[42]。
3.3 基于表型与生化成分两种聚类结果的比较
利用聚类分析对茶树品种资源进行分类能够快捷、准确地得出资源间的相似性并展现出品种的综合性状,被灵活运用于育种工作的亲本选配环节。本研究基于表型性状与主要生化成分分别对32份供试材料进行聚类分析,不同分析标准聚类结果不完全一致,基于表型性状的分析将供试材料分为4类,而基于主要生化成分的分析将供试材料分为3类。基于表型的聚类结果中聚在类群Ⅰ的石阡苔茶、竹山大黑叶、上犹群体种、龙井种、南山白毛茶、罗坑群体种、六堡茶在基于生化成分的聚类结果中分离至类群Ⅱ与类群Ⅲ;基于表型的聚类结果中聚在类群Ⅱ的信阳种、牙己茶、十里香在基于生化成分的聚类结果中分离至类群Ⅰ与类群Ⅲ;基于表型的聚类结果中聚在类群Ⅲ的遂川群体种、浮粱种在基于生化成分的聚类结果中分离至类群Ⅰ。两种聚类结果也有相同的地方,例如基于表型性状的聚类分析中聚为类群Ⅳ的勐库大叶茶与凤庆大叶茶在基于生化成分的分析结果中依旧聚在同一类中,且距离相近;黎家坪群体种、冯家台群体种、汝城白毛茶、永修群体种、宜昌大叶茶始终在同一类群中;祁门种、黄山种、鸟王茶、云台山种、紫阳种总是聚在同一类群。
综合分析两种聚类结果表明,表型性状与生化成分间具有一定相关性,说明供试材料遗传多样性丰富、遗传背景复杂。表型多样性是遗传多样性与环境多样性共同作用的结果[43],有研究表明,茶树表型性状易受环境条件影响,尤其是数量性状变异较大,表现出丰富的遗传多样性[44]。生化成分含量水平的变异在长期的自然选择与人为驯化下发生了明显的变异[45]。两种聚类结果的差异表明,依据单一表型或生化成分的聚类分析只能作为参考,在实际应用中还需结合更深层且灵敏度更高的分子标记方法,综合考虑表型、生化成分与DNA分子间的遗传多样性,才能高效精准地指导茶树育种工作。
[1] 郭燕, 张树航, 李颖, 等. 燕山板栗种质资源叶片表型性状多样性研究[J]. 园艺学报, 2022, 49(8): 1673-1688. Guo Y, Zhang S H, Li Y, et al. Diversity analysis of leaves phenotypic traits of Yanshan Chestnut [J]. Acta Horticulturae Sinica, 2022, 49(8): 1673-1688.
[2] 姬广磊. 黄鮟鱇()形态特征及其遗传学研究[D]. 青岛: 中国海洋大学, 2007. Ji G L. Study on the morphological characters and genetics of[D]. Qingdao: Ocean University of China, 2007.
[3] 陈琪予, 马建强, 陈杰丹, 等. 利用图像特征分析茶树成熟叶表型的遗传多样性[J]. 茶叶科学, 2022, 42(5): 649-660. Chen Q Y, Ma J Q, Chen J D, et al. Genetic diversity of mature leaves of tea germplasms based on image features [J]. Journal of Tea Science, 2022, 42(5): 649-660.
[4] Naegele R P, Mitchell J, Hausbeck M K. Genetic diversity, population structure, and heritability of fruit traits in[J]. PLos One, 2016, 11(7): e0156969. doi: 10.1371/journal.pone.0156969
[5] 吴国林. 基于ISSR和SSR标记的鄱阳湖流域野生菰种质资源遗传多样性研究[D]. 南昌: 江西农业大学, 2014. Wu G L. Studies on genetic diversity of germplasm resources of(Griseb.) Trucz. based on ISSR and SSR markers [D]. Nanchang: Jiangxi Agricultural University, 2014.
[6] 刘彤. 柳州九万山野生茶树种质资源研究[D]. 长沙: 湖南农业大学, 2016. Liu T. The research of Liuzhou Jiuwan Mountain wild tea germplasm resources [D]. Changsha: Hunan Agricultural University, 2016.
[7] 莫岚, 黄亚辉, 赵文芳, 等. 金秀野生茶树群体的生化特征[J]. 福建农林大学学报(自然科学版), 2018, 47(4): 409-414. Mo L, Huang Y H, Zhao W F, et al. Analysis of biochemical components of wild tea colonies in Jinxiu [J]. Journal of Fujian Agriculture and Forestry University (Natural Science Edition), 2018, 47(4): 409-414.
[8] 晏嫦妤, 李家贤, 黄华林, 等. 广东茶树群体品种品质成分差异初步研究[J]. 广东农业科学, 2009, 59(12): 39-41. Yan C Y, Li J X, Huang H L, et al. Study on differences in quality composition of Guangdong tea germplasms [J]. Guangdong Agricultural Sciences, 2009, 59(12): 39-41.
[9] 黄亚辉, 罗荣康, 晏嫦妤, 等. 连南茶树种质资源叶片表型性状及生化成分的研究[J]. 广东茶业, 2017, 45(6): 11-13. Huang Y H, Luo R K, Yan C Y, et al. Study on leaf phenotypic traits and biochemical components of tea germplasm resources in Liannan [J]. Guangdong Tea Industry, 2017, 45(6): 11-13.
[10] 王小萍, 唐晓波, 王迎春, 等. 52份茶树资源生化组分的表型多样性分析[J]. 茶叶科学, 2012, 32(2):129-134. Wang X P, Tang X B, Wang Y C, et al. Phenotypic diversity analysis of biochemical composition in 52 tea germplasms [J]. Journal of Tea Science, 2012, 32(2): 129-134.
[11] 甘春萍, 邓慧群, 陈佳, 等. 广西地方群体品种主要生化成分分析研究[J]. 农业与技术, 2012, 32(4): 95-99. Gan C P, Deng H Q, Chen J, et al. Analysis of main biochemical components of Guangxi local population varieties [J]. Agriculture and Technology, 2012, 32(4): 95-99.
[12] 万明轩. 菠菜表型三维点云数据分析与形态品质指标遗传多样性研究[D]. 上海: 上海师范大学, 2020. Wan M X. Study on analysis of three-dimensional point cloud data of spinach phenotype and genetic diversity of morphological quality index [D]. Shanghai: Shanghai Normal University, 2020.
[13] 张悦, 索玉静, 孙鹏, 等. 柿种质资源果实形态多样性分析[J]. 园艺学报, 2022, 49(7): 1473-1490. Zhang Y, Suo Y J, Sun P, et al. Analysis on fruit morphological diversity of persimmon germplasm resources [J]. Acta Horticulturae Sinica, 2022, 49(7): 1473-1490.
[14] 陈亮, 杨亚军. 茶树种质资源描述规范和数据标准[M]. 北京: 中国农业出版社, 2005. Chen L, Yang Y J. Description specification and data standard of tea germplasm resources [M]. Beijing: China Agriculture Press, 2005.
[15] 郭荣华, 刘志斋, 蔡一林, 等. 两个历史上重要的玉米地方品种群体的表型多样性评价[J]. 玉米科学, 2009, 17(2): 24-28. Guo R H, Liu Z Z, Cai Y L, et al. Evaluation of phenotypic diversity for two maize landrace population shaving historical importance in China [J]. Journal of Maize Sciences, 2009, 17(2): 24-28.
[16] 丁洲, 李烨昕, 袁艺, 等. 安徽茶区优良群体种的表型性状和遗传多样性分析[J]. 茶叶科学, 2018, 38(2):155-161. Ding Z, Li Y X, Yuan Y, et al. Phenotypic traits and genetic diversity of elite tea population in Anhui (China) [J]. Journal of Tea Science, 2018, 38(2): 155-161.
[17] 黄海涛, 王风雷, 王贤波, 等. 鸠坑茶树资源芽叶表型性状的多样性分析[J]. 浙江农业科学, 2013, 64(3): 251-254. Huang H T, Wang F L, Wang X B, et al. Diversity analysis of bud and leaf phenotypic traits of Jiukeng tea resources [J]. Journal of Zhejiang Agricultural Sciences, 2013, 64(3): 251-254.
[18] 刘青. 贵州三都县古茶树遗传多样性研究[D]. 贵阳: 贵州大学, 2020. Liu Q. Genetic diversity of ancient tea trees in Sandu County, Guizhou Province [D]. Guiyang: Guizhou University, 2020.
[19] 周炎花. 基于叶片形态和EST-SSR茶树遗传多样性与遗传演化研究[D]. 福州: 福建农林大学, 2010. Zhou Y H. Study of genetic diversity and genetic evolution of tea germplasms based on leaf morphology and EST-SSR markers [D]. Fuzhou: Fujian Agriculture and Forestry University, 2010.
[20] 胡波, 李国元, 谢志兵, 等. 主成分分析法在冬枣果实贮藏特性分析中的应用[J]. 食品科技, 2014, 39(2): 44-47. Hu B, Li G Y, Xie Z B, et al. Application of principle component analysis on winter jujube storage characteristic [J]. Food Science and Technology, 2014, 39(2): 44-47.
[21] 杨玲, 张彩霞, 康国栋, 等. ‘华红’苹果果肉的流变特性及其主成分分析[J]. 中国农业科学, 2015, 48(12): 2417-2427. Yang L, Zhang C X, Kang G D, et al. Rheologic properties of ‘Huahong’ apple pulp and their principal component analysis [J]. Scientia Agricultura Sinica, 2015, 48(12): 2417-2427.
[22] 郎彬彬, 黄春辉, 朱博, 等. 基于果实相关性状的江西野生毛花猕猴桃初级核心种质的构建方法研究[J]. 果树学报, 2016, 33(7): 794-803. Lang B B, Huang C H, Zhu B, et al. Study on the method of constructing a primary core collection of Jiangxi wildbased on fruit traits [J]. Journal of Fruit Science, 2016, 33(7): 794-803.
[23] 李伟, 郜海燕, 陈杭君, 等. 基于主成分分析的不同品种杨梅果实综合品质评价[J]. 中国食品学报, 2017, 17(6): 161-171. Li W, Gao H Y, Chen H J, et al. Evaluation of comprehensive quality of different varieties of bayberry based on principal components analysis [J]. Journal of Chinese Institute of Food Science and Technology, 2017, 17(6): 161-171.
[24] 李勋兰, 洪林, 王武, 等. 晚熟杂柑新品种果实品质综合评价[J]. 果树学报, 2018, 35(2): 195-203. Li X L, Hong L, Wang W, et al. Comprehensive evaluation of fruit quality of new late-maturing mandarin cultivars [J]. Journal of Fruit Science, 2018, 35(2): 195-203.
[25] 吕正鑫, 贺艳群, 贾东峰, 等. 猕猴桃种质资源表型性状遗传多样性分析[J]. 园艺学报, 2022, 49(7): 1571-1581. Lü Z X, He Y Q, Jia D F, et al. Genetic diversity analysis of phenotypic traits for kiwifruit germplasm resource [J]. Acta Horticulturae Sinica, 2022, 49(7): 1571-1581.
[26] 林子翔. 观赏辣椒种质资源园艺性状的多元统计分析与种质创新研究[D]. 杭州: 浙江农林大学, 2020. Lin Z X. Multivariate statistical analysis and germplasm innovation of ornamental pepper germplasm resources [D]. Hangzhou: Zhejiang A&F University, 2020.
[27] 宁功伟, 杨盛美, 段志芬, 等. 云南野生茶树厚轴茶种质资源化学成分多样性分析[J/OL]. 分子植物育种, 2023: 1-20[2023-05-06]. https://kns.cnki.net/kcms/detail//46. 1068. S.20230118.1701.005.html. Ning G W, Yang S M, Duan Z F, et al. Diversity analysis of chemical components of wild tea germplasm resources () in Yunnan [J/OL]. Molecular Plant Breeding, 2023: 1-20[2023-05-06]. https://kns.cnki. net/ kcms/detail//46.1068.S.20230118.1701.005.html.
[28] 董方, 李小飞, 沈思言, 等. 江西茶树资源的遗传多样性分析及优异种质筛选[J]. 江西农业大学学报, 2022, 44(6): 1466-1477. Dong F, Li X F, Shen S Y, et al. Genetic diversity analysis and screening of excellent germplasm of tea plant resources in Jiangxi [J]. Acta Agriculturae Universitatis Jiangxiensis, 2022, 44(6): 1466-1477.
[29] 丁帅涛, 程晓梅, 张亚, 等. 基于表型性状和生化成分的陕西茶树种质资源遗传多样性研究[J]. 西北农业学报, 2019, 28(4): 607-619. Ding S T, Cheng X M, Zhang Y, et al. Genetic diversity in phenotypic traits and biochemical components of tea resources in Shanxi [J]. Acta Agriculturae Boreali-occidentalis Sinica, 2019, 28(4): 607-619.
[30] Zhang Y R, Fu J M, Zhou Q Y, et al. Metabolite profiling and transcriptome analysis revealed the conserved transcriptional regulation mechanism of caffeine biosynthesis in tea and coffee germplasm plants [J]. Journal of Agricultural and Food Chemistry, 2022, 70(10): 3239-3251.
[31] Li G D, Li Y, Yao X Z, et al. Establishment of a virus-induced gene-silencing (VIGS) system in tea plant and its use in the functional analysis of[J]. International Journal of Molecular Sciences, 2023, 24(1): 392. doi: 10.3390/ijms24010392.
[32] Li P H, Ye Z L, Fu J M, et al. CsMYB184 regulates caffeine biosynthesis in tea plants [J]. Plant Biotechnology Journal, 2022, 20(6): 1012-1014.
[33] Ma W H, Kang X, Liu P, et al. The NAC-like transcription factor CsNAC7 positively regulates the caffeine biosynthesis-related genein[J]. Horticulture Research, 2022, 9: uhab046. doi: 10.1093/hr/uhab046.
[34] Jin J Q, Yao M Z, Ma C L, et al. Natural allelic variations of TCS1 play a crucial role in caffeine biosynthesis of tea plant and its related species [J]. Plant Physiology and Biochemistry, 2016, 100(12): 18-26.
[35] 蔡志翔, 严娟, 宿子文, 等. 不同类型桃种质资源主要酚类物质含量评价[J]. 园艺学报, 2022, 49(5): 1008-1022. Cai Z X, Yan J, Su Z W, et al. Genetic diversity analysis of phenotypic traits for kiwifruit germplasm resources [J]. Acta Horticulturae Sinica, 2022, 49(5): 1008-1022.
[36] Zhu Q F, Liu L J, Lu X F, et al. The biosynthesis of EGCG, theanine and caffeine in response to temperature is mediated by hormone signal transduction factors in tea plant (L.) [J]. Frontiers in Plant Science, 2023, 14. doi: 10.3389/fpls.2023.1149182.
[37] 蒋会兵, 田易萍, 陈林波, 等. 云南茶树地方品种农艺性状与品质性状遗传多样性分析[J]. 植物遗传资源学报, 2013, 14(4): 634-640. Jiang H B, Tian Y P, Chen L B, et al. Diversity of tea landraces based on agronomic and qualitytraits in Yunnan Province [J]. Journal of Plant Genetic Resources, 2013, 14(4): 634-640.
[38] 姚明哲. 利用ISSR和EST-SSR标记研究中国茶树资源的遗传多样性和遗传结构[D]. 杭州: 浙江大学, 2009. Yao M Z. Studies on genetic diversity and structure of tea germplasm in China based on ISSR and EST-SSR markers [D]. Hangzhou: Zhejiang University, 2009.
[39] 陈勋, 龚自明. 基于EST-SSR标记的湖北茶树种质资源遗传多样性分析[J]. 分子植物育种, 2017, 15(5): 1831-1838. Chen X, Gong Z M. Analysis of genetic diversity with EST-SSR markers for tea germplasm in Hubei Province [J]. Molecular Plant Breeding, 2017, 15(5): 1831-1838.
[40] 张晓丽, 郭辉, 王海岗, 等. 中国普通野生稻与栽培稻种SSR多样性的比较分析[J]. 作物学报, 2008, 105(4): 591-597. Zhang X L, Guo H, Wang H G, et al. Comparative assessment of SSR allelic diversity in wild and cultivated rice in China [J]. Acta Agronomica Sinica, 2008, 105(4): 591-597.
[41] 刘振, 赵洋, 杨培迪, 等. 湖南省茶树种质资源现状及研究进展[J]. 茶叶通讯, 2011, 38(3): 7-10, 18. Liu Z, Zhao Y, Yang P D, et al. The status and research progress of Hunan tea germplasm [J]. Tea Communication, 2011, 38(3): 7-10, 18.
[42] 马建强, 姚明哲, 陈亮. 茶树种质资源研究进展[J]. 茶叶科学, 2015, 35(1):11-16. Ma J Q, Yao M Z, Chen L. Research progress on germplasms of tea plant () [J]. Journal of Tea Science, 2015, 35(1):11-16.
[43] 张见明. 26份武夷名丛茶树种质资源秋茶生化成分多样性分析[J]. 武夷学院学报, 2016, 35(9): 1-7. Zhang J M. Diversity analysis of biochemical components in autumn shoots of 26 Wuyi Mingcong tea plant germplasm resources [J]. Journal of Wuyi University, 2016, 35(9): 1-7.
[44] 乔婷婷. 茶树资源遗传多样性及其表型性状关联EST-SSR位点的初步鉴定[D]. 北京: 中国农业科学院, 2010. Qiao T T. Genetic diversity of tea ((L.)O. Kuntze) and association analysis on phenotypic traits with EST-SSR markers [D]. Beijing: Chinese Academy of Agricultural Sciences, 2010.
[45] 齐冬晴. 不同茶树品种(品系)鲜叶香气及其表型差异的研究[D]. 长沙: 湖南农业大学, 2015. Qi D Q. Studies on the tea leaves aroma components and phenotypic differences of different tea varieties [D]. Changsha: Hunan Agricultural University, 2015.
Diversity Analysis of Leaf Phenotype and Biochemical Components in Tea Local Population Resources
TANG Lu, LI Changle, GE Yue, WANG Pu, ZHAO Hua, WANG Mingle, WANG Yu, GUO Fei*, NI Dejiang
National Key Laboratory for Germplasm Innovation & Utilization of Horticultural Crops, College of Horticulture and Forestry Sciences, Huazhong Agricultural University, Wuhan 430070, China
In order to protect tea germplasm resources and promote tea germplasm innovation, the phenotypic character and biochemical component diversity of 32 tea germplasm resources from 12 provinces were studied. The results showed that the genetic diversity index of 16 phenotypic character ranged from 0.81 to 1.94, and the phenotypic genetic diversity of the test population was high. The cumulative contribution rate of the six phenotypic charactersincluding leaf area, leaf shape, leaf upper surface, young shoot pubescence, leaf tip shape and leaf cross section were 71.78%, which was the main factor causing the phenotypic difference of the tested tea population. Based on the determination results of biochemical components in two years, the variation coefficient of caffeine content (17.95% and 14.55%) was the largest, followed by the variation coefficient of tea polyphenol content (13.61% and 8.11%).The variation coefficient of free amino acid (5.62% and 7.52%) was the lowest. Cluster analysis based on the results of biochemical component content determination was conducted to divide the tested tea tree populations into three groups, each including different types of germplasm. From the geographical distribution of the tested tea populations, the content of biochemical components showed regional differences.
tea local population species, phenotypic character, biochemical components, diversity
S571.1
A
1000-369X(2023)04-473-16
2023-05-06
2023-07-17
农业生物种质资源挖掘与利用(2022YFD1200505)、临沧市建设国家可持续发展议程创新示范区科技专项(202204AC100001-A01)
唐璐,女,硕士研究生,主要从事茶树种质资源方面研究,Tanglu@webmail.hzau.edu.cn。*通信作者:guofei@mail.hzau.edu.cn
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-21
乡村地理(2018年2期)2018-09-19
西南农业学报(2018年5期)2018-06-05
现代园艺(2017年21期)2018-01-03
茶叶学报(2017年3期)2017-11-23
湖南农业(2016年3期)2016-06-05
家庭科学·新健康(2016年5期)2016-05-12
音乐天地(音乐创作版)(2016年11期)2016-02-05
中国康复理论与实践(2015年10期)2015-12-24
医学研究杂志(2015年5期)2015-06-10
