
烤烟Y2001多腺毛改良株系的创制及抗性分析
2023-08-28 13:12罗锐王召军滕环瑜张博凯高嘉宁田涛闫筱筱崔红
烟草科技 2023年8期
罗锐,王召军,滕环瑜,张博凯,高嘉宁,田涛,闫筱筱,崔红*
1.河南农业大学烟草学院,郑州市郑东新区平安大道218号 450046
2.河南中烟工业有限责任公司,郑州市管城回族区经开第三大街9号 450000
烟草表面密布表皮毛,根据其有无分泌能力可以分为分泌型表皮毛和非分泌型表皮毛[1]。其中,分泌型表皮毛又称腺毛,根据其柄细胞数目的多少又分为长柄腺毛和短柄腺毛[2]。腺毛产生的次生代谢物质是烟草叶面化学成分的重要组成部分[3]。烟草腺毛分泌物主要由二萜类和糖酯等化合物构成[4]。其中,西柏烷类二萜占腺毛分泌物的大部分,主要以西柏三烯一醇和二醇的形式存在[5]。腺毛分泌物对烟叶香气品质至关重要,西柏烷类二萜可降解产生茄酮以及茄酮降解产物,是烟草重要的中性香气物质,具有改善烟气香气和吸味的作用[6-7]。蔗糖酯是卷烟燃吸时的重要香气前体物质,同时也是后期卷烟加工的保润剂[8]。此外,腺毛分泌物在烟株应对环境胁迫过程中也发挥着重要作用,例如分泌物对蚜虫等害虫有明显的趋避及毒杀作用[9-10],有效抑制了以昆虫为媒介的病害的传播[11]。同时,腺毛分泌物对真菌孢子囊生长也有抑制作用[12-13]。因此,提高腺毛密度及分泌物含量是烟草育种的重要目标之一。
近年来,植物表皮毛发生相关基因的研究在多种模式植物中越来越深入[14],许多重要调控基因被鉴定出来,例如在番茄中的研究发现了1个B型细胞周期蛋白SlCycB2[15],过表达SlCycB2能够显著降低番茄表皮毛密度,而SlCycB2 RNAi植株的表皮毛密度则显著增加[16]。对本氏烟草中的同源基因NbCycB2的研究也证明该基因是腺毛发生的负调控基因[17]。孟盈等[18]、潘阳等[19]对普通烟草中的B型细胞周期蛋白基因的同源基因NtCycB2研究也发现,NtCycB2基因敲除后,腺毛密度及腺毛分泌物显著提高,烤后烟叶中性致香物质的积累量也显著提高。此外,NtCycB2基因敲除后植株生长发育正常,对盐[20]、干旱[21]、蚜虫[22]等胁迫的抗性也明显提高。说明NtCycB2是进行腺毛密度及分泌物含量定向改良的理想基因。中烟100是豫中浓香型烟区的主栽品种,因产量高、易烘烤、耐肥性强等特点在生产上得到广泛应用。李艳华等[23]对其腺毛观察发现,与K326等优质烤烟品种相比,中烟100叶面非分泌型表皮毛比例偏高,导致分泌物含量较低,推测可能对烤后烟叶香气不利。为此,课题组前期选育出了分泌型腺毛比例较高的改良品种Y2001[24],其香气品质优于中烟100,且保留中烟100的优异农艺性状。Y2001的选育表明通过腺毛定向改良能够提高烟叶的香气品质,由此推测提高Y2001腺毛密度及分泌物含量有望进一步提高烟叶的香气品质。但高腺毛密度对Y2001的生物及非生物胁迫抗性是否也会产生影响,目前还鲜见研究报道。因此,采用EMS诱变创制了Y2001 NtCycB2基因突变体,并以该突变体为供体亲本,Y2001为轮回亲本,进行连续回交和自交,结合分子标记辅助选择、腺毛观察、腺毛分泌物测定,得到了多腺毛Y2001定向改良株系HY2001,并对其干旱、黑胫病和蚜虫胁迫抗性进行分析,旨在明确高腺毛密度烟草株系抗性的优劣,为豫中浓香型烟叶产区品种改良奠定基础。
1 材料与方法
1.1 试验材料
供试材料为Y2001,将Y2001裸种用清水洗净后采用0.8%(质量分数)甲基磺酸乙酯(EMS)处理,于人工气候室中进行培养。培养条件:光照14 h、28℃,黑暗10 h、24℃,光暗交替,相对湿度60%。
1.2 NtCycB2基因突变体的筛选
将EMS处理后的Y2001种子(M0)采用漂浮育苗培养至四叶一心期,共获得1 716株成活幼苗。采用CTAB法分别提取各单株的DNA,随后将提取的DNA按照10株1组进行等量混合,形成DNA混池,并根据NtCycB2基因序列设计特异性扩增引物NtCycB2(表1),对所有DNA混池及野生型Y2001基因组DNA进行PCR扩增,对PCR产物进行测序,筛选含有序列突变的混池,并对混池中的各个单株分别测序,最终确定突变单株。

表1 引物列表Tab.1 Primer list
1.3 NtCycB2基因突变检测分子标记的开发
根据突变体中NtCycB2基因序列的突变位点,开发了特异性检测标记MT(表1),该标记能够在含有突变位点的单株基因组DNA中扩增出条带。但该标记为显性标记,无法区分纯合单株和杂合单株,为此又针对野生型序列开发了特异性检测标记WT(表1),该标记能够在含有野生型序列的单株基因组DNA中扩增出条带。分别使用MT与WT引物对待测植株DNA进行扩增,仅有MT能扩增出条带,即为NtCycB2基因纯合突变单株。
1.4 多腺毛Y20.1 改良株系的创制
以筛选得到的NtCycB2基因突变体(M0代杂合单株)为供体亲本,Y2001为轮回亲本进行连续回交,每个世代均利用分子标记辅助选择NtCycB2基因杂合突变的单株进行下一轮回交,为加快回交进程,回交过程采用诱导早花技术进行室内加代[25]。进行4次回交后自交得到BC4F2,在BC4F2代中筛选NtCycB2基因纯合突变单株,进行腺毛表型观察及分泌物检测,并利用烟草全基因组SNP芯片进行遗传背景回复率的检测。烟草叶片用液氮速冻后,送至中国烟草总公司郑州烟草研究院国家烟草基因研究中心进行SNP芯片检测。候选单株自交收获种子,所得BC4F3代作为候选改良株系HY2001(图1)。
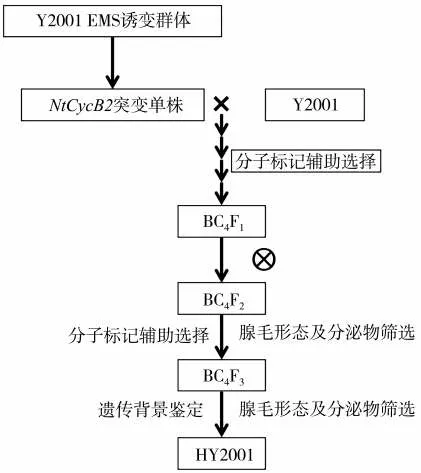
图1 HY2001的创制过程Fig.1 Creation process of HY2001
1.5 腺毛形态观察
在烟苗四叶一心期,选用同一叶位发育正常、大小一致的叶片,置于0.2%(质量体积分数)的罗丹明B水溶液中浸染30 min,染色结束后用蒸馏水漂洗3次,冲洗掉未结合的染料。用滤纸吸干叶表面水分后,置于超景深显微镜(VHX5000,日本基恩士公司)下进行腺毛观察,并在上表皮中部随机选择3个视野进行腺毛形态观察及密度统计。
1.6 腺毛分泌物分析
于烟苗十叶一心期,取同一叶位发育正常、大小一致的叶片进行分泌物的提取和检测,样品制备以及前处理参考韩锦峰等[26]的方法。从叶片上截取直径为5 cm的叶圆片,每50个叶圆片作为1个生物学重复,每个材料设置3次生物学重复。将叶圆片在二氯甲烷溶液中浸提,每个叶圆片浸提8次,每次2 s。在浸提液中加入1 mL内标(2.020 mg·mL-1的蔗糖八乙酸酯和2.542 mg·mL-1的正十七烷醇的混合溶液),混合均匀后进行过滤,过滤后利用旋转蒸发仪(上海申生科技有限公司)浓缩,经硅烷化处理后利用气相色谱质谱联用仪TRACE GC Ultra-DSQ IIMS(美国赛默飞世尔科技公司)进行检测,色谱检测参数参考王霄龙等[27]的方法。采用内标定量法(相对校正因子为1)进行定量分析。
1.7 基因表达水平测定
于烟苗四叶一心期,选取中烟100、Y2001与HY2001从上至下第2叶位叶片,按照植物总RNA提取试剂盒[DP423,天根生化科技(北京)有限公司]说明书提取总RNA,反转录为cDNA,以烟草L25核糖体蛋白基因为内参基因,采用荧光定量PCR方法,对二萜合成基因NtCYP71D16、NtCBTS、NtABS以及糖酯类合成相关基因NtASAT1、NtASAT2进行表达量分析,中烟100、Y2001与HY2001均设置3次生物学重复。引物序列如表1所示。
1.8 干旱胁迫抗性分析
将Y2001与HY2001裸种置于铺有湿润滤纸的培养皿上,于人工气候室中培养至两片子叶后,小心移栽至相同大小的培养钵中,并于烟苗生长至四叶一心期时停止浇水,模拟自然干旱胁迫并进行表型观察。干旱处理15 d后,分别取干旱处理和未进行处理的烟株从上至下第2叶位叶片,使用台盼蓝(Trypan Blue)和硝基四氮唑蓝(NBT)染色并进行死细胞数量和活性氧状况观察,具体操作参照关扬扬等[21]的方法。使用试剂盒(北京索莱宝公司)测定干旱处理及未处理的烟株从上至下第3叶位叶片的超氧化物歧化酶(SOD)、过氧化氢酶(CAT)及过氧化物酶(POD)的活性,具体操作按照试剂盒说明书进行。均设置3次生物学重复。
1.9 黑胫病抗性分析
疫霉菌(Phytophthora nicotiana)由河南省农业科学院烟草研究所提供,病菌首先使用燕麦培养基培养[28],待菌丝铺满整个平板后,方可使用。
选用六叶一心期的壮苗,Y2001与HY2001均设置6次生物学重复。采用针刺法[29]进行疫霉接种,使用无菌注射器针尖在茎部制造伤口,使用打孔器在培养好疫霉菌丝的培养基上打取直径5 mm的圆形菌饼,菌丝面朝伤口放置,用浸湿无菌水的脱脂棉覆盖菌饼,保持菌丝生长所需水分,再用保鲜膜覆盖脱脂棉防止水分过快挥发,在完成接种后,每天揭开保鲜膜喷水保持其湿度。在接种后第7天,根据国家标准GB/T 23222─2008[30]对Y2001和HY2001的发病程度进行调查,并取感病茎部发病及未发病交界处组织保存于-80℃,提取RNA后进行抗病相关基因表达量分析(Y2001和HY2001均设置3次生物学重复),引物序列见表1。
1.10 蚜虫抗性分析
离体叶片蚜虫选择试验:选取四叶一心期、大小一致的叶片,用脱脂棉包裹叶柄,Y2001与HY2001各选择3片叶作为3次重复,放置于铺好滤纸的培养皿中,所有叶片与培养皿中心距离相等,在培养皿中心放置50头蚜虫,使蚜虫进行自然选择,每2 h统计蚜虫数量,连续统计12 h。
蚜虫繁殖试验:选取四叶一心期的Y2001与HY2001烟苗放置于光照培养箱中,每株接种20头蚜虫,用防虫网与外界隔离,设置3次生物学重复,每3 d统计1次蚜虫数量,共统计5次。
1.11 数据处理
使用Primer Premier 5.0软件设计引物,使用Microsoft Excel软件计算平均数及标准差,使用SPSS 17.0软件进行ONE WAY ANOVA分析。采用Student's t-test进行两组数据间比较,采用LSD法进行多组数据间的差异显著性检验。
2 结果与分析
2.1 多腺毛Y20.1 改良株系的创制
对野生型Y2001中的NtCycB2基因进行测序,发现其序列与已经发表的K326中的序列完全一致[18],说明Y2001中NtCycB2基因没有发生突变。通过混池测序,成功筛选得到了1个NtCycB2基因杂合突变单株,该突变单株中NtCycB2基因的1条拷贝在37~85 bp处共发生了36个碱基缺失,以及5个碱基替换(图2A),针对序列突变设计了特异性检测标记MT(表1)。以该突变单株为供体亲本,Y2001为轮回亲本,连续回交4代,之后自交2代,结合分子标记辅助选择(图2B)、腺毛表型观察及分泌物检测,最终得到了NtCycB2基因纯合突变的多腺毛Y2001改良株系HY2001(图1)。

图2 HY2001幼苗植株形态及分子标记辅助筛选Fig.2 Plant morphology and molecular marker assisted selection of HY2001 seedlings
对室内植株形态观察发现(图2C、图2D),与中烟100相比Y2001和HY2001叶片狭长,叶面褶皱,叶边缘呈波浪形。
利用烟草全基因组SNP芯片对HY2001进行背景回复率检测,结果(图3)表明HY2001与Y2001的遗传背景相似度为97.86%。
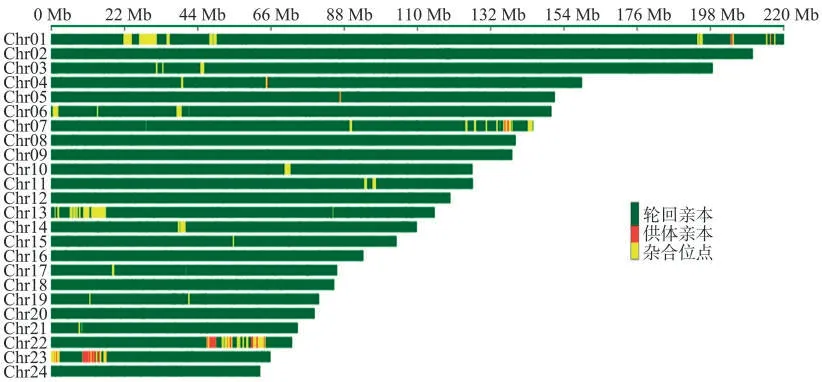
图3 HY2001遗传背景分析Fig.3 Genetic background analysis of HY2001
2.2 多腺毛Y20.1 改良株系腺毛密度分析
腺毛形态观察与腺毛密度统计结果见图4。从图4中可看出,中烟100、Y2001和HY2001叶片上同时存在长柄腺毛、短柄腺毛和非分泌型表皮毛。但中烟100非分泌型表皮毛所占有比例较高,而Y2001和HY2001主要由长柄腺毛构成,且HY2001不仅分泌型腺毛的比例提高,腺毛密度也明显增加,其中长柄腺毛的密度增加最明显,并且部分长柄腺毛存在分支现象,腺头饱满,发育完善,染色较中烟100与Y2001加深。从腺毛密度统计结果可以看出,与中烟100相比,HY2001长柄腺毛增加2.68倍,短柄腺毛提高1.71倍,非分泌型表皮毛降低34%,腺毛比例得到优化且腺毛密度显著提高。与Y2001相比,HY2001长柄分泌型腺毛密度增加97%,短柄分泌型腺毛提高76%,非分泌型表皮毛比例也有所增加。
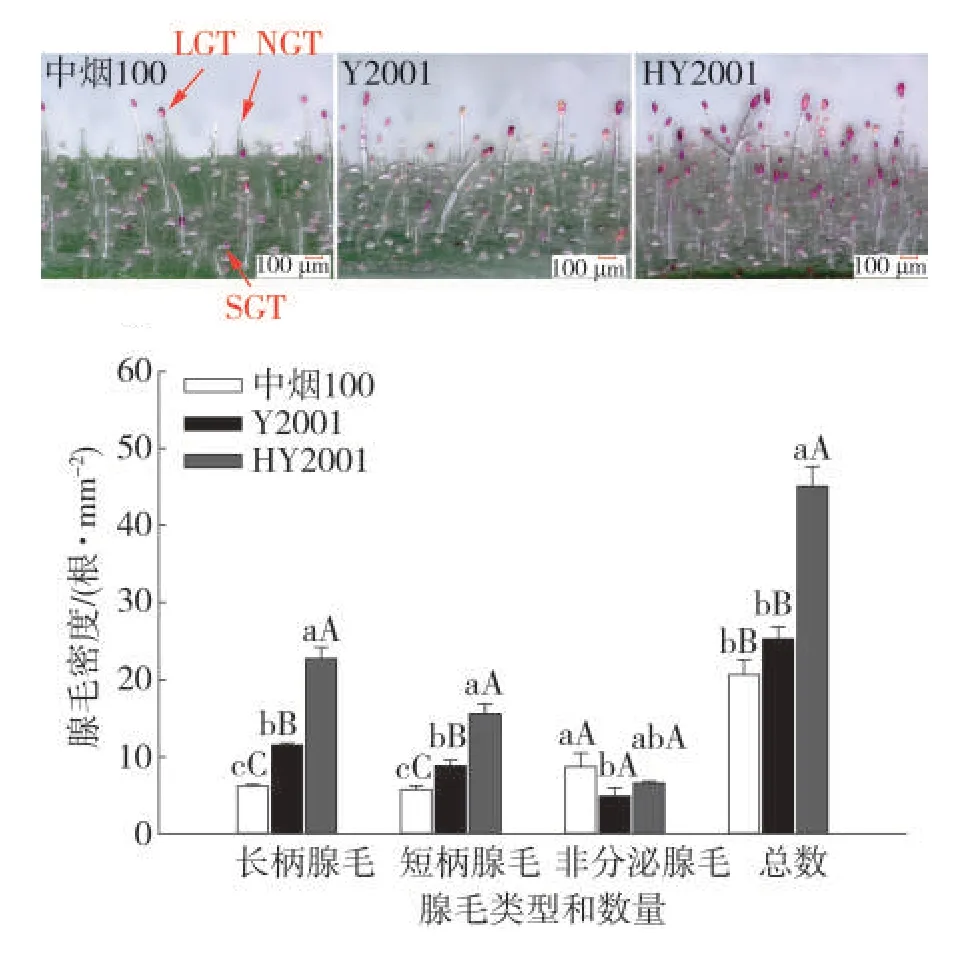
图4 中烟100、Y2001与HY2001的腺毛观察及密度统计Fig.4 Observation and density statistical analysis of trichomes of Zhongyan 100,Y2001 and HY2001
2.3 多腺毛Y20.1 改良株系腺毛分泌物含量分析
对中烟100、Y2001与HY2001叶片腺毛分泌物进行萃取并进行GC/MS检测,结果见图5。从图5中可以看出,中烟100、Y2001与HY2001的分泌物组分一致,均由西柏烷二萜和I、II型蔗糖酯构成(图5A)。从含量来看,Y2001相较于中烟100各组分含量均显著提高;而HY2001中各组分含量显著高于Y2001,其中西柏烷类二萜化合物含量分别为中烟100和Y2001的3.03倍和1.98倍,蔗糖酯含量分别为中烟100和Y2001的3.43倍和2.63倍,这与腺毛密度的显著提高有关。
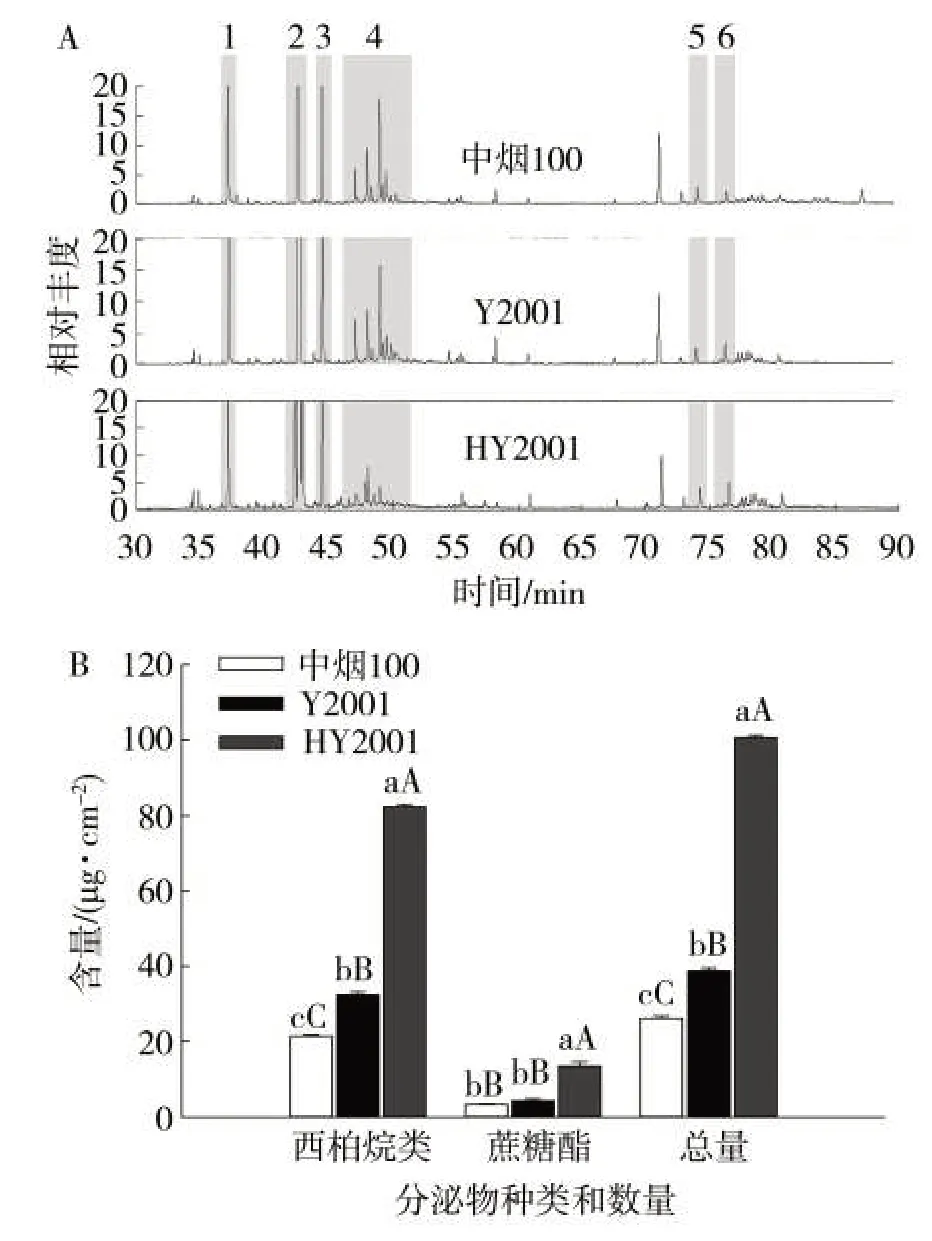
图5 中烟100、Y2001与HY2001腺毛分泌物含量分析Fig.5 Secretion contents of trichomes of Zhongyan 100,Y2001 and HY2001
2.4 腺毛分泌物合成相关基因表达分析
中烟100、Y2001及HY2001中的二萜合成基因NtCYP71D16、NtCBTS、NtABS及蔗糖酯合成基因NtASAT1、NtASAT2的表达量测定结果如图6所示。5个腺毛分泌物合成相关基因的表达均发生不同程度的上调。其中与二萜合成相关的NtCYP71D16、NtCBTS与NtABS表达水平相较于中烟100和Y2001均显著提高,这一结果与二萜类化合物增长相符合。同时糖酯合成相关基因NtASAT1、NtASAT2的表达量也有一定的上调,分别是中烟100与Y2001的3.18倍和1.31倍、1.90倍和1.63倍,这与叶面分泌物中蔗糖酯含量结果一致。
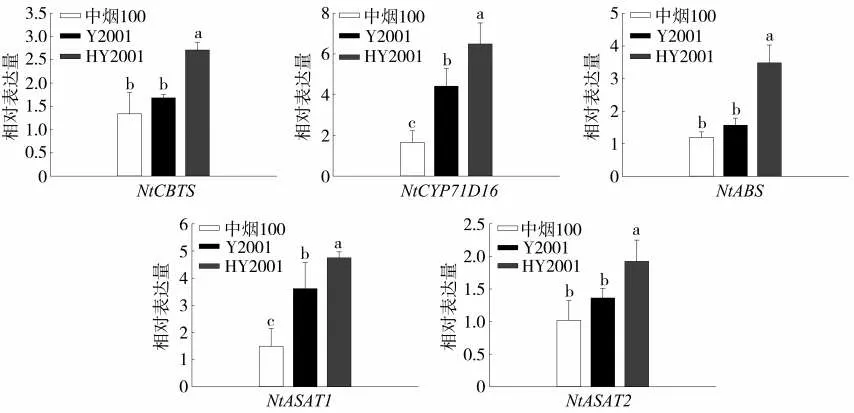
图6 中烟100、Y2001与HY2001腺毛分泌物合成基因表达水平分析Fig.6 Expression levels of trichome secretion biosynthesis related genes in Zhongyan 100,Y2001 and HY2001
2.5 干旱胁迫抗性分析
图7结果显示,干旱处理15 d时,Y2001中部以及下部叶片变黄萎蔫严重,仅上部叶片萎蔫较轻,生长状态较好;而HY2001只有靠近下部叶片萎蔫,轻微变黄,上部叶片状态良好(图7A)。对干旱过程中细胞氧化损伤程度进行了Trypan Blue染色及NBT染色观察死细胞及活性氧积累状况发现,两种染色条件均为HY2001染色最浅(图7B),说明HY2001受到的氧化损伤较轻。抗氧化酶活性分析结果(图7C)显示,干旱处理0 d的Y2001和HY2001的抗氧化酶活性相近,干旱处理15 d的叶片组织内的抗氧化酶活性均升高,但HY2001体内抗氧化酶活性明显高于Y2001,说明HY2001能更好地抵御干旱所带来的细胞氧化损伤。

图7 Y2001和HY2001干旱胁迫植株生长状况及氧化损伤程度比较Fig.7 Comparison of plant growth status and oxidative damage degree between Y2001 and HY2001 under drought stress
2.6 黑胫病抗性分析
将制备好的烟草疫霉菌菌饼分别接种于Y2001与HY2001烟苗茎基部。在接种后的第7天,Y2001植株呈现萎蔫状态,茎部呈枯木状,黑胫病斑环绕茎围,叶片呈凋萎状,靠近下部叶片也有病斑出现,且根部有腐烂的迹象,而HY2001植株整体状态较好,茎部病斑未达到茎围二分之一,且只有下部一片叶轻度凋萎,根部状态完好(图8A、图8C)。分别将Y2001和HY2001的茎基部患病部分剖开,发现Y2001茎部发生黑色病变,髓部干缩呈碟片状,失去运送水分及养分能力,而HY2001髓部组织发生轻度萎缩,但韧皮部完好(图8B),说明HY2001能够更好地抵抗病菌侵袭。

图8 Y2001和HY2001疫霉菌侵染后表型观察比较Fig.8 Comparison of phenotype between Y2001 and HY2001 infected by Phytophthora parasitica
对接菌前后抗病相关基因表达水平进行测定发现,在接菌前HY2001的抗病相关基因表达量略高于Y2001,但无显著差异。接菌后病程相关蛋白基因NtPR1b、细胞凋亡相关蛋白基因Nthsr203J、丝氨酸蛋白酶抑制基因NtNmIMSP、谷胱甘肽S-转移酶基因NtGST、细胞色素基因NtP450-1与NtP450-2表达水平在HY2001和Y2001中均有提高,但HY2001提高程度高于Y2001,分别是Y2001的1.56、1.28、1.33、2.06、1.75和1.22倍(图9),除NtP450-2外其他基因在HY2001和Y2001之间的差异均达到显著水平。说明HY2001对于黑胫病菌的抗性显著提高。

图9 Y2001与HY2001抗病相关基因表达量分析Fig.9 Expression levels of disease resistance related genes in Y2001 and HY2001
2.7 蚜虫抗性分析
取Y2001及HY2001离体叶片进行蚜虫选择试验结果如图10A和B所示。可以看出HY2001叶片上蚜虫数量始终显著低于Y2001,说明HY2001的蚜虫嗜好性显著低于Y2001。比较蚜虫在Y2001及HY2001烟株上的繁殖速率结果如图10C和图10D所示。随着时间的推进,Y2001和HY2001叶片上蚜虫数量不断增多,但HY2001上蚜虫数目始终少于Y2001,接蚜第15天时,Y2001蚜虫数量达到460头,与第1天相比,增长23.00倍,而HY2001蚜虫数量达到275头,与第1天相比增长13.75倍,增幅明显小于Y2001。且第15天时Y2001蚜虫的数量是HY2001的1.67倍,极显著高于HY2001。表明HY2001株系上蚜虫繁殖速率显著低于Y2001。
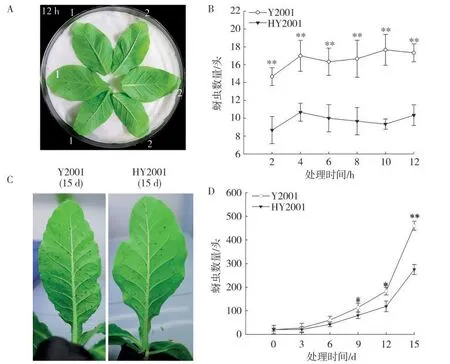
图10 Y2001和HY2001的蚜虫抗性比较Fig.10 Comparison of aphid resistance between Y2001 and HY2001
3 讨论
中烟100是豫中浓香型烟叶产区的主栽品种,Y2001是针对叶面非分泌型表皮毛比例偏高、腺毛分泌物含量低的问题选育而来的改良品种,证明了通过腺毛改良能有效提高烟叶的香气品质。本研究中通过EMS诱变创制了Y2001 NtCycB2基因突变体,并以Y2001为供体亲本,通过连续回交、自交及分子标记辅助选择,结合腺毛表型观察及分泌物检测,成功创制出多腺毛Y2001改良株系HY2001,该株系腺毛密度相比中烟100与Y2001显著提高,西柏烷二萜和蔗糖酯含量也高于中烟100和Y2001,实现了提高Y2001腺毛密度与分泌物含量的目的,且HY2001与Y2001的遗传背景相似度为97.86%,基本恢复了Y2001的遗传背景。由于材料创制过程是在实验室完成的,下一步将在河南浓香型烟叶产区进行多点小区试验及生产对比试验,以比较二者的植物学及农艺性状,从而更好评估HY2001的应用潜力。
有研究表明,腺毛能够在烟叶表面形成一层空气薄膜,减弱水分蒸腾[31],本研究中也发现,HY2001的抗旱性优于Y2001。此外腺毛密度及分泌物的增加也可作为屏障阻止昆虫在植物表面的移动,从而减少昆虫对植物体的侵害[32]。同时,腺毛分泌的次生代谢物质,可起到驱赶和毒杀的作用[33]。本研究中室内蚜虫抗性分析结果表明,HY2001对蚜虫的吸引力及蚜虫在HY2001叶片上的繁殖速率均低于Y2001,对蚜虫的抗性显著增强,由此推测以蚜虫为媒介的烟草病毒病发病率将会降低。室内接菌试验发现HY2001对疫霉菌抗性明显优于Y2001,其抗病相关基因表达量显著提升,这些基因在调节植物体内抗病物质的合成、提高防御酶活性、细胞过敏性死亡及抗病信号的传导等方面起重要作用[34-35],推测HY2001能更快地合成抗病物质并通过细胞凋亡来阻止疫霉菌丝对烟株的进一步侵染。由于本研究只在盆栽接种条件下调查了HY2001对烟草黑胫病的症状表现及抗病相关基因的表达,目前其抗性机理还不清楚,有待进一步深入研究。
4 结论
通过对Y2001 NtCycB2基因进行诱变,创制出多腺毛改良株系HY2001。HY2001株系的烟叶腺毛密度增加,分泌能力旺盛,二萜合成基因表达水平上调,叶面分泌物成分积累量显著提高。同时烟株抗氧化能力增强,抗病相关基因表达量水平提高,蚜虫的选择嗜好性及繁殖率降低,对干旱、黑胫病以及蚜虫的抗性显著提高。因此,HY2001株系在豫中浓香型烟叶产区具有较大的应用潜力。
猜你喜欢
亚热带植物科学(2022年1期)2022-05-17
中国烟草学报(2022年2期)2022-04-29
江淮法治(2022年2期)2022-03-17
河南农业科学(2021年8期)2021-10-05
四川农业科技(2019年5期)2019-07-01
中国烟草学报(2018年2期)2018-05-09
浙江柑橘(2016年1期)2016-03-11
湖南农业科学(2015年3期)2015-10-09
中国烟草学报(2012年4期)2012-04-09
中国烟草科学(2010年1期)2010-07-31
