
治疗黑色素瘤的新型二芳基衍生物04022a在不同种属肝微粒体中的稳定性及代谢产物分析
2023-08-14 01:39李梦君刘波王晓波李妍熊山牟艳玲解维林
药学研究 2023年7期
李梦君,刘波,王晓波,李妍,熊山,牟艳玲,解维林
(山东第一医科大学药学院<药物研究所>,先进药物递释系统全国重点实验室,国家卫健委生物技术药物重点实验室,山东省罕少见病重点实验室,山东 济南 250117)
黑色素瘤是一种源于黑色素细胞恶变而形成的肿瘤,恶性程度极高,易发生转移[1]。尽管黑色素瘤仅占皮肤癌的5%,但它却占皮肤癌相关死亡的75%[2]。早期首选的治疗方式为手术切除,选择做有足够安全范围的切除,可使患者具有良好的预后。但是对于手术无法切除,或者发生多处转移导致切除效果欠佳的晚期患者,则预后极差,因此迫切需要寻找更有效的治疗晚期黑色素瘤的药物。
鞘磷脂是一类在调节细胞功能中具有多种重要作用的脂类。鞘磷脂的代谢产物,包括神经酰胺(Cer)、鞘氨醇(Sph)和1-磷酸鞘氨醇(S1P),在细胞生长、分化、存活和凋亡中发挥着重要的作用。Sph通过鞘氨醇激酶1或2(SphK1或SphK2)磷酸化进一步产生S1P。S1P是一种重要的信号分子,它可以作为细胞内传递信号的第二信使。S1P已被证明可以调节内质网中游离钙离子的释放,这对细胞增殖有广泛影响[3]。也可以通过旁分泌或自分泌的方式与细胞膜上相应的G蛋白偶联受体结合作为第一信使参与生物效应,包括细胞增殖[4]、凋亡[5]、诱导血管生成[6]、促肿瘤炎症[7]以及激活侵袭和转移[8]等。SphK1和SphK2同工酶过表达有相反功能,SphK1促进细胞生长,抑制细胞凋亡,而SphK2与细胞生长抑制和增强细胞凋亡相关[9]。Sphk1表达水平在黑色素瘤[10-11]、结肠癌[12]、胃癌[13]、肺癌[14]等多种肿瘤组织中都明显高于正常组织。SphK1抑制、沉默或功能丧失突变可降低黑色素瘤中S1P的含量,促进抗肿瘤免疫[10],抑制血管生成[15],诱导黑色素瘤细胞凋亡和自噬,因此,SphK1是黑色素瘤治疗的理想靶标。
本课题组从SphK1的晶体结构出发,设计筛选出具有抑制活性的SphK1抑制剂SAMS10,并通过对先导化合物SAMS10进行结构修饰,合成了一系列的新型二芳基衍生物。前期研究表明新型二芳基衍生物04022a对黑色素瘤A375细胞系具有显著的抗癌活性,抑制A375细胞迁移,促进凋亡,并通过诱导A375细胞G2/M期阻滞来发挥抗增殖作用。通过对裸鼠构建异种移植瘤模型,表明它对肿瘤的生长具有剂量依赖性的抑制作用[11]。本研究采用肝微粒体温孵体系,考察04022a在不同种属肝微粒体中的代谢稳定性;采用超高效液相色谱-四极杆飞行时间质谱联用(UPLC-QTOF-MS/MS)方法,分离检测了04022a在肝微粒体中的代谢物,结合母药的结构性质和二级质谱图的碎片信息,鉴定了代谢产物的结构、可能的代谢途径和裂解途径,初步探讨该化合物的代谢规律,判断其成药性,为之后体内的药代动力学研究奠定基础。
1 材料
1.1 仪器Ultimate 3000型高效液相色谱仪(美国赛默飞世尔公司);Triple TOF 5600质谱仪(美国AB SCIEX公司),恒温混匀仪MTH-100(杭州米欧仪器有限公司);ME155DU型电子天平(梅特勒-托利多有限公司);IKAVortex 3涡旋混匀器(广州艾卡仪器设备有限公司);SB-5200DT超声波清洗机(宁波新芝生物科技股份有限公司);高速台式冷冻离心机(德国Eppendorf公司)。
1.2 药品与试剂04022a(纯度>97%)由山东省医学科学院药物研究所合成;睾丸酮购自阿拉丁生化科技有限公司;PBS缓冲液(0.01 mol·L-1,pH 7.2~7.4)购自北京索莱宝生物科技有限公司;大鼠肝微粒体(RLMS,目录号:M10011,20 mg·mL-1)、人肝微粒体(HLMS,目录号:M10001,20 mg·mL-1)和比格犬肝微粒体(DLMS,目录号:M10007,20 mg·mL-1)均购自武汉普莱特生物医药技术有限公司;NADPH再生系统(溶液A:26.1 mmol·L-1NADP+, 66 mmol·L-1葡萄糖6-磷酸,66 mmol·L-1MgCl2;溶液B:40 U·mL-1葡萄糖6-磷酸脱氢酶,5 mmol·L-1柠檬酸钠)购自北京汇智泰康医药技术有限公司;甲醇由美国赛默飞世尔公司所提供;甲酸由天津科密欧化学试剂有限公司所提供;甲醇、甲酸均为色谱纯。
2 方法
2.1 代谢稳定性研究的HPLC条件采用Inertsil ODS色谱柱(4.6 mm×250 mm,3.5 μm)进行样品分离,柱温25 ℃。流动相由含有0.1%甲酸的水(溶剂A)和甲醇(溶剂B)组成,采用梯度洗脱的方式,洗脱的梯度如下:0~3 min,5%B;3~25 min,5%B→50%B;25~27 min,50%B→70%B;27~30 min,70%B;30~30.01 min,70%B→5%B;30.01~35 min,5%B,分析的总时长为35 min。自动进样器的温度为25 ℃,流速为1 mL·min-1,进样量为10 μL,紫外检测器的检测波长为280 nm。
2.2 代谢产物研究的UPLC-QTOF-MS/MS条件超高效液相色谱配备有岛津的超高效液相色谱仪(日本京都)用于样品分析,采用Phenomenex Kinetex C18色谱柱(2.1 mm×100 mm,1.7 μm)分离母药与代谢产物,柱温40 ℃。流动相由溶剂A(含有0.1%甲酸的水)和溶剂B(甲醇)组成。分析物以0.3 mL·min-1的流速用以下梯度洗脱:0~2 min,5%B;2~18 min,5%B→35%B;18~58 min,35%B→50%B;58~88 min,50%B→70%B;88~114 min,70%B→95%B;114~117 min,95%B;117~117.1 min,95%B→5%B;117.1~120 min,5%B,整个色谱运行时间为120 min。自动进样器的温度设定为25 ℃,进样体积为10 μL。质谱检测由配备Duo-SprayTM离子源的Triple TOFTM5600系统在正电喷雾电离(ESI)模式下进行。质谱仪和UPLC系统由带有Analyst 1.8软件的工作站控制。优化的操作参数如下:离子源电喷雾电压(IS),5 500 V;雾化气压力(GS1),45 psi;加热气压力(GS2),45 psi;气帘气(CUR),35 psi;加热气温度(TEM),500 ℃;去簇电压(DP),80 eV;碰撞能量(CE),20 eV;碰撞能量的步阶(CES),5 eV。IDA方法包括一次TOF-MS测量扫描(m/z100-800)和10次产物离子IDA扫描(m/z50~800),累积时间分别为0.25 s和0.1 s。
2.3 溶液的配制精密称定13.58 mg的0 4022a,用1 mL的甲醇溶解,得到浓度为20 μmol·mL-1的母药溶液,用于进行实验组的肝微粒体孵育实验;另外称取1.02 mg的04022a,用1.5 mL的甲醇溶解制备成1 μmol·mL-1的储备液,并进一步用甲醇稀释成0.04、0.08、0.1、0.2、0.4、0.8 μmol·mL-1的系列对照品工作液,按“2.1”项下的色谱条件来检测样品,记录样品的峰面积。横坐标X是待测物04022a的质量浓度,纵坐标Y是对应的峰面积,以此来绘制标准曲线。
用1 mL的甲醇充分溶解精密称定的5 mg睾丸酮,制备成浓度为5 mg·mL-1的母药溶液,用于进行阳性对照组肝微粒体的孵育实验;另外称取1 mg的睾丸酮,溶于甲醇当中,配制成1 mg·mL-1的储备液,将储备溶液用甲醇逐级稀释来制备标准工作溶液,睾丸酮的浓度为:2、8、20、50、200、400、800 μg·mL-1。按“2.1”项下的色谱条件来检测样品,记录样品的峰面积。横坐标X是睾丸酮的浓度,纵坐标Y是对应的峰面积,以此来构建标准曲线。
2.4 代谢稳定性孵育体系预孵育体系的总体积为188 μL,包括181 μL 0.01 mol·L-1的PBS缓冲液,2 μL浓度为20 μmol·mL-1的04022a母药溶液以及5 μL不同种属的肝微粒体,为避免肝微粒体酶失活应在冰上加入。将其在37 ℃,850 r·min-1条件下预孵育5 min后,加入12 μL的NADPH再生系统启动反应(整个孵育体系中加入甲醇的最大体积为1%),继续温孵0、15、30、60、90、120 min后,立即加入等体积的冰甲醇使反应停止。将终止反应后的样品涡旋1 min,于4 ℃,14 000 r·min-1离心10 min,收集上清液,按“2.1”项下的色谱条件来检测样品,测定各时间点待测物04022a的含量,平行实验3次。同时设置阳性对照组(用睾丸酮代替待测物),阴性对照组(不加NADPH),空白对照组(不添加待测物,用PBS代替)。
2.5 代谢产物研究按“2.4”项下的孵育体系处理样品,将实验组和空白对照组中孵育120 min后的样品,按照“2.2”项下的条件进样分析。通过对比二者的总离子流图,寻找04022a在各种属肝微粒体中的代谢产物,再根据各色谱峰的准分子离子以及碎片离子信息,推测代谢产物可能的结构与裂解途径。
3 结果
3.1 代谢稳定性阳性对照组的孵育体系中用睾丸酮来代替待测物04022a,以此来验证该温孵体系的建立是否成功;阴性对照组的孵育体系中没有加NADPH,以此来考察04022a在酶不起作用的情况下能否发生代谢;空白对照组中不含待测物,将实验组和空白对照组的图谱进行比较,来判断是否发生了代谢,结果见图1。在孵育120 min时,阳性对照组中睾丸酮在大鼠、人、比格犬肝微粒体中的剩余药物百分比分别为6.59%±0.06%、7.73%±0.13%、82.75%±1.69%,可以看出睾丸酮在不同种属的肝微粒体中均发生了明显的代谢,且代谢速率较快,说明该孵育体系的构建是可靠的,可以用来进行待测物的体外代谢稳定性研究。

A.阴性对照组;B.实验组;C.阳性对照组
使用GraphPad Prism 8.0.1软件来绘制待测物04022a在不同种属肝微粒体中的孵育曲线,所得结果见图1。在孵育120 min时,实验组中04022a在大鼠、人、比格犬肝微粒体中的剩余药物百分比分别为88.30%±1.41%、82.15%±2.23%、98.63%±0.45%。阴性对照组中04022a在大鼠、人、比格犬肝微粒体中的剩余药物百分比分别为98.76%±1.54%、101.98%±2.98%、103.42%±0.69%。结果表明04022a在不同种属的肝微粒体中均发生了不同程度的代谢,且原型药的消除依赖CYPs。
3.2 酶动力学参数的计算纵坐标Y是各温孵时间点的平均剩余药物百分比的自然对数,横坐标X是孵育时间,以此作线性回归,根据斜率k值来计算酶动力学参数,结果见表1。根据体外半衰期可以看出该化合物代谢是否稳定。表1得出的结果显示,04022a在SD大鼠、人、比格犬肝微粒体中的代谢时间较长,代谢稳定性良好。

表1 04022a在大鼠、人和比格犬肝微粒体孵育体系中的酶动力学参数比较
3.3 代谢产物研究通过比较含药生物样品与空白生物样品的色谱图差异,初步判断04022a在大鼠和人肝微粒体中有5个相同的体外代谢产物,在比格犬肝微粒体中有4个代谢产物,代谢物的详细信息见表2,包括保留时间、可能的元素组成、特征碎片离子以及可能的代谢反应类型等。母药及其代谢物的提取离子色谱图如图2所示。
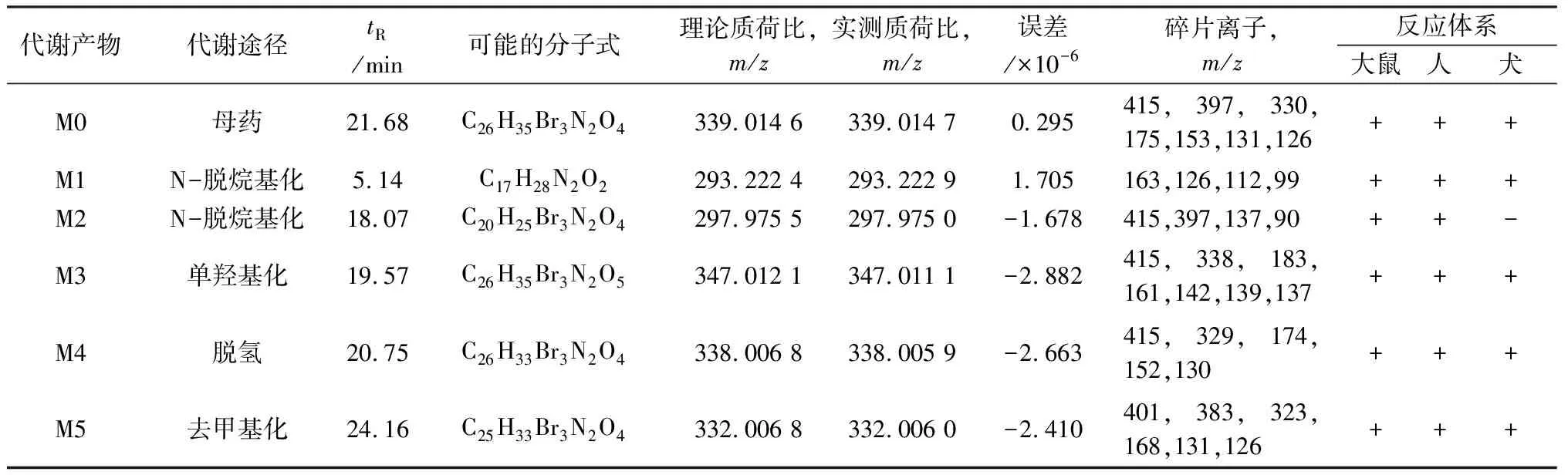
表2 UPLC-QTOF-MS/MS检测的不同种属肝微粒体中04022a代谢产物的质谱数据

图2 RLMS(A)、HLMS(B)和DLMS(C)中04022a代谢物的提取离子流图
3.3.1 04022a的质谱分析为了鉴定04022a的代谢物,了解母体药物(M0)的裂解过程具有重要意义。采用UHPLC-Q-TOF-MS在正ESI扫描模式下研究了M0的色谱和质谱行为。M0在21.68 min时被检测到,在m/z339处显示出分子离子[M+2H]2+,化学式为C26H35Br3N2O4。因为结构中含有3个Br,MS图中显示出Br的同位素峰,[M+2H]2+(339)∶[M+2H+2]2+(340)∶[M+2H+4]2+(341)∶[M+2H+6]2+(342)=1∶3∶3∶1,又因各同位素峰之间的分子量相差1 Da,因此均为[M+2H]2+峰。m/z415、131的碎片离子为N-C键发生断裂产生,m/z397比415少18 Da,是其脱去一分子H2O所形成的产物离子;m/z330为M0脱水形成的碎片离子,O-C键继而断裂形成m/z为175的[M+2H]2+碎片;C-C键断裂形成m/z为153的[M+2H]2+碎片;在m/z为126的[M+2H]2+碎片离子是由O-C键断裂产生的。M0的MS/MS谱图和碎裂途径如图3所示。

图3 04022a的MS2质谱图和可能的裂解途径
3.3.2 04022a的代谢产物分析在保留时间5.14 min观察到代谢产物M1的m/z293的质子化分子离子[M+H]+,因其MS图中并不存在Br的同位素峰,说明该化合物不含有Br元素,化学式为C17H28N2O2,推测其为母药发生N-脱烷基化所产生的I相代谢产物。N-C键发生断裂产生m/z为163的碎片;M1的O-C键发生断裂形成m/z为126的碎片离子,126分别失去CH3、C2H5形成m/z112、m/z99的产物离子。M1的MS/MS谱图和碎裂途径如图4所示。

图4 04022a代谢产物的MS2质谱图和可能的裂解途径
代谢产物M2在18.07 min的色谱图中出现,分子离子[M+2H]2+在m/z297,MS图中含有3个Br的同位素峰,[M+2H]2+(297)∶[M+2H+2]2+(298)∶[M+2H+4]2+(299)∶[M+2H+6]2+(300)=1∶3∶3∶1,说明结构中有Br的存在[16-17],且各同位素峰之间的分子量相差1 Da,因此均为[M+2H]2+峰。化学式为C20H25Br3N2O4。推测其为母药的哌啶环发生开环继而N脱烷基化形成的I相代谢产物。m/z415、90为M2的N-C键发生断裂所产生的两个碎片离子,m/z397与415相差18 Da,推测由失去一分子H2O所形成,m/z90失去CH2CH2NH2形成m/z为137的关键碎片离子。M2的MS/MS谱图和碎裂途径如图4所示。
代谢产物M3在19.57 min洗脱,在m/z347处显示分子离子[M+2H]2+,MS图中存在Br的同位素峰,[M+2H]2+(347)∶[M+2H+2]2+(348)∶[M+2H+4]2+(349)∶[M+2H+6]2+(350)=1∶3∶3∶1,说明结构中有3个Br的存在[16-17],且因各同位素峰之间的分子量相差1 Da,因此均为[M+2H]2+峰,化学式为C26H35Br3N2O5。M3的m/z347的分子离子[M+2H]2+以及m/z为 338、183、161、139的主要碎片离子[M+2H]2+与M0的m/z339的分子离子[M+2]2+以及m/z为330、175、153、131的主要碎片离子[M+2H]2+相比均多了8 Da,M3的m/z142的碎片离子比M0的m/z为126的碎片离子大16 Da,因此推测M3为母药发生单羟基化产生的I相代谢产物。m/z415为N-C键发生断裂产生;m/z137是由m/z139中的O-C键发生断裂所产生。由于m/z415、142、137这些特征碎片离子的存在,推测羟基化发生在哌啶环上,但具体的位置无法根据现有的质谱信息推断。M3的MS/MS谱图和碎裂途径如图4所示。
代谢产物M4在m/z338以分子离子[M+2H]2+的形式出现,保留时间为20.75 min,MS图中可以观察到Br的同位素峰,[M+2H]2+(338)∶[M+2H+2]2+(339)∶[M+2H+4]2+(340)∶[M+2H+6]2+(341)=1∶3∶3∶1,说明含有3个Br[16-17],且各同位素峰之间的分子量相差1 Da,因此均为[M+2H]2+峰,化学式为C26H33Br3N2O4。其[M+2H]2+峰m/z338比M0小1 Da,推测M4为母药脱氢产生的I相代谢产物。m/z415为N-C键发生断裂形成的产物离子;主要的碎片离子m/z329、174、152、130均比母药MS2图中的碎片离子330、175、153、131少1 Da,因此推测这些碎片离子与母药具有相同的裂解途径。因为m/z415、130这两个特征性产物离子的存在,推测脱氢发生在1-乙基-4-甲基哌啶环上。M4的MS/MS谱图和碎裂途径如图4所示。
在UPLC体系中,代谢产物M5的峰位于m/z332,洗脱时间为24.16 min,MS图中存在Br的同位素峰,[M+2H]2+(332)∶[M+2H+2]2+(333)∶[M+2H+4]2+(334)∶[M+2H+6]2+(335)=1∶3∶3∶1,说明含有3个Br原子[16-17],同时各同位素峰之间的分子量相差1 Da,因此均为[M+2H]2+峰,化学式为C25H33Br3N2O4。M5在m/z为332的分子离子[M+2H]2+以及其在m/z为323、168的主要碎片离子[M+2H]2+与M0在m/z为339的分子离子[M+2H]2+以及其在m/z为330、175的主要碎片离子[M+2H]2+相比均少了7 Da,因此推测M5为母药去甲基产生的I相代谢产物。m/z401、131为M5的N-C键发生断裂产生的关键碎片离子,m/z383比401的分子量小18 Da,是其脱去一分子H2O所形成的;M5中的O-C键断裂产生m/z为126碎片离子。因为m/z401、131和126这些诊断性碎片离子的存在,推测是母药N原子上的甲基丢失。M5的MS/MS谱图和碎裂途径如图4所示。
在肝微粒体中共鉴定出5种代谢产物。04022a在体外肝微粒体中主要通过脱烷基化、羟基化、脱氢等途径来进行代谢,结果如图5所示。

图5 04022a在RLMS、HLMS和DLMS中可能的代谢途径
4 讨论
肝脏是人体的主要代谢器官,对于外界侵入的物质可以通过代谢转化以降低所带入的毒性,从而有效达到保护机体的作用,当有毒外源性物质侵入机体内部时,必然会受到毒副作用影响或对机体产生不良反应,因此药物体外代谢模型主要基于肝脏。肝微粒体方法不仅制备简便,重现性好,而且酶混合物易于保存,培养条件易于优化。因此,在新药开发初期,常采用肝微粒体模型考察化合物代谢的速度和程度,对候选化合物的药代动力学特征进行初步的研究,以确定候选化合物在研发初期是否值得进一步开发[14]。本研究成功创建了一种简单、稳定、专属性较强且重复性较高的HPLC分析方法,测定肝微粒体中04022a的质量浓度。从测定结果可以看出,该化合物代谢稳定性良好,在HLMS和RLMS中的代谢行为较为相似。
本实验利用高效液相色谱串联飞行时间质谱技术,建立了简单、快速、灵敏度和分辨率高的分析方法,对04022a及其代谢产物进行分离鉴定。从样品的总离子流色谱图中可以得到的保留时间;从一级质谱图中可以获得母药及代谢产物的相对分子质量,根据常见的氧化、还原、水解和结合等代谢反应规律,再结合母药的结构分析,可以对代谢产物的结构和生物转化途径进行推测;通过二级质谱图的碎片信息,可以进一步推测代谢产物碰撞诱导产生碎片离子的裂解途径;通过计算理论质荷比和实测质荷比的误差,辅助验证推测的结果。研究表明,04022a在体外肝微粒体中主要通过脱烷基化、羟基化、脱氢等途径来进行代谢。该化合物在大鼠和人肝微粒体中产生了相同的代谢产物,在体外具有相同的代谢途径,之后选择大鼠来研究体内的生物转化情况。
猜你喜欢
食品工业科技(2021年17期)2021-09-14
高师理科学刊(2020年11期)2021-01-04
理化检验-化学分册(2020年3期)2020-04-24
天然产物研究与开发(2018年9期)2018-10-08
山东化工(2018年16期)2018-09-12
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
中成药(2016年4期)2016-05-17
华南农业大学学报(2015年5期)2015-12-04
特产研究(2015年1期)2015-04-12
