
低氧运动预适应激活自噬调控Hippo/YAP/TAZ信号通路减轻急性低氧力竭运动大鼠骨骼肌损伤
2023-07-13 09:40孔海军谌晓安
山东体育学院学报 2023年3期
关键词:自噬
孔海军 谌晓安
摘 要:目的:探討低氧运动预适应激活自噬调控Hippo/YAP/TAZ信号通路对急性低氧力竭运动大鼠骨骼肌保护的机制。方法:50只6周龄SPF级雄性SD大鼠,随机分为空白对照组(Con组,n=10)、单纯急性低氧力竭运动组(HE组,n=10)、低氧预适应+急性低氧力竭运动组(HP+HE组,n=10)、运动预适应+急性低氧力竭运动组(EP+HE组,n=10)、低氧预适应+运动预适应+急性低氧力竭运动组(EP/HP+HE组,n=10)。预适应周期2 w,EP+HE组及EP/HP+HE组进行2次/d常压常氧跑台训练;HP+HE组及EP/HP+HE组进行10 h/d间歇性低氧暴露(FiO2 13.5%)。预适应后HE组、HP+HE组、EP+HE组及EP/HP+HE组大鼠进行低氧环境跑台力竭训练(FiO2 11.9%~12.0%)。低氧力竭运动后2 h,尾静脉取血检测血清骨骼肌损伤、炎症反应和氧化应激指标,苏木精-伊红染色观察腓肠肌病理特征,并检测腓肠肌自噬相关蛋白、Hippo/YAP/TAZ信号通路蛋白及mRNA。结果:(1)与HE组比较,预适应组大鼠低氧力竭运动后的腓肠肌结构损伤、炎症反应和氧化应激明显减轻。(2)HE组Atg5、Beclin1蛋白表达显著低于EP+HE组及EP/HP+HE组(P<0.05),EP/HP+HE组Atg5、Beclin1蛋白表达显著高于HP+HE组(P<0.05);HE组LC3-Ⅱ蛋白表达显著高于EP+HE组及EP/HP+HE组(P<0.05),EP+HE组及EP/HP+HE组显著高于HP+HE组(P<0.05)。(3)HE组MST1 mRNA显著高于EP+HE组及EP/HP+HE组(P<0.05),EP+HE组及EP/HP+HE组显著高于HP+HE组(P<0.05);HE组LATS1 mRNA显著低于预适应组(P<0.05);HE组YAP mRNA显著高于预适应组(P<0.05),EP/HP+HE组显著低于HP+HE组(P<0.05);HE组及HP+HE TAZ mRNA显著高于EP+HE组及EP/HP+HE组均显著降低(P<0.05)。(4)HE组MST1、LATS1及其磷酸化水平显著低于EP+HE组及EP/HP+HE组(P<0.05),EP/HP+HE组MST1显著高于HP+HE组(P<0.05);HE组YAP蛋白显著高于EP+HE组及EP/HP+HE组(P<0.05);HE组TAZ蛋白显著高于预适应组均显著降低(P<0.05),EP/HP+HE组均显著低于HP+HE组(P<0.05)。结论:低氧运动预适应可能通过激活骨骼肌自噬抑制Hippo/YAP/TAZ信号通路减轻急性低氧力竭运动大鼠的骨骼肌损伤、炎症反应和氧化应激反应。
关键词:低氧运动预适应;自噬;Hippo/YAP/TAZ信号通路;急性低氧力竭运动;骨骼肌损伤
中图分类号:G804.2 文献标识码:A文章编号:1006-2076(2023)03-0089-10
Hypoxic Exercise Preconditioning Activates Autophagy to Regulate Hippo/YAP/TAZ Signaling Pathway to Alleviate Skeletal Muscle Injury in Acute Hypoxic Exhaustive Exercise Rats
KONG Haijun1,2,CHEN Xiaoan1
1.College of P.E., Jishou University, Xiangxi 416000, Hunan, China; 2. College of P.E., Kashgar University, Kashgar 844000, Xinjiang, China
Abstract:Objective:To explore the mechanism of hypoxic exercise preconditioning activating autophagy to regulate Hippo/YAP/TAZ signaling pathway to protect skeletal muscle in rats undergoing acute hypoxic exhaustive exercise. Methods: 50 six-week-old SPF male SD rats were randomly divided into blank control group (Con group, n=10), simple acute hypoxic exhaustive exercise group (HE group, n=10), hypoxic preconditioning+acute hypoxic exhaustive exercise group (HP+HE group, n=10), exercise preconditioning+acute hypoxic exhaustive exercise group (EP+HE group, n=10), hypoxic preconditioning+exercise preconditioning+acute hypoxic exhaustive exercise group (EP/HP+HE group, n=10). The preconditioning period was 2 weeks, EP+HE group and the EP/HP+HE group were trained twice a day on the normal pressure and oxygen treadmill; HP+HE group and EP/HP+HE group were exposed to intermittent hypoxia with FiO2 13.5% for 10 hours/day. On the next day after preconditioning, rats in HE group, HP+HE group, EP+HE group and EP/HP+HE group were trained to run on the treadmill in a hypoxic environment (FiO2 11.9%~12.0%). Two hours after exercise, tail vein blood was taken to detect serum skeletal muscle injury, inflammatory reaction and oxidative stress indicators, gastrocnemius muscle tissue was taken for HE section staining and autophagy related protein, Hippo/YAP/TAZ signal pathway protein and mRNA were detected. Results: (1) Compared with HE group, the structural damage, inflammatory reaction and oxidative stress of gastrocnemius muscle of rats in preconditioning group were significantly reduced after hypoxic exhaustive exercise. (2) The expression of Atg5 and Beclin1 protein in HE group was significantly lower than that in EP+HE group and EP/HP+HE group(P<0.05), and the expression of Atg5 and Beclin1 protein in EP/HP+HE group was significantly higher than that in HP+HE group (P<0.05); The expression of LC3-Ⅱ protein in HE group was significantly higher than that in EP+HE group and EP/HP+HE group (P<0.05), and that in EP+HE group and EP/HP+HE group was significantly higher than that in HP+HE group (P<0.05). (3) MST1 mRNA in HE group was significantly higher than that in EP+HE group and EP/HP+HE group (P<0.05), and that in EP+HE group and EP/HP+HE group was significantly higher than that in HP+HE group (P<0.05); LATS1 mRNA in HE group was significantly lower than that in preconditioning group (P<0.05); YAP mRNA in HE group was significantly higher than that in preconditioning group (P<0.05), while that in EP/HP+HE group was significantly lower than that in HP+HE group (P<0.05); The mRNA of HE group and HP+HE TAZ was significantly higher than that of EP+HE group and EP/HP+HE group (P<0.05). (4) MST1, LATS1 and their phosphorylation levels in HE group were significantly lower than those in EP+HE group and EP/HP+HE group (P<0.05), and MST1 in EP/HP+HE group was significantly higher than those in HP+HE group (P<0.05); YAP protein in HE group was significantly higher than that in EP+HE group and EP/HP+HE group (P<0.05); TAZ protein in HE group was significantly higher than that in preconditioning group (P<0.05), while that in EP/HP+HE group was significantly lower than that in HP+HE group (P<0.05). Conclusion:Hypoxic exercise preconditioning may inhibit Hippo/YAP/TAZ signal pathway by activating skeletal muscle autophagy, thereby reducing skeletal muscle injury, inflammatory response and oxidative stress response in rats undergoing hypoxic exhaustive exercise.
Key words:hypoxic exercise preconditioning; autophagy; Hippo/YAP/TAZ signal path; acute hypoxic exhaustive exercise; skeletal muscle injury
急性高强度运动可能形成机械、代谢及氧化等多方面、复合性压力,短时间过度载荷可能诱导机体炎症反应、氧化应激,同时存在骨骼肌和内脏器官组织损伤风险[1]。人体及动物实验证据表明[2],急性高强度运动可诱导血清、骨骼肌、心肌等组织炎症及骨骼肌损伤标记物水平激增。此外,低氧条件下的急性运动运动通过激活氧化应激会加剧骨骼肌承受的复合压力[3]。有研究认为,模拟3 000 m海拔低氧环境75%~80% HRmax强度力竭跑导致运动员血清骨骼肌损伤标志物激增,且参与对抗压力源的蛋白表达水平急剧增加[4]。近年来的研究证实,运动可激活机体自噬途径,从而提高组织细胞自调控能力及能量稳态[5]。自噬途径激活可能参与急、慢性运动中骨骼肌功能整合过程,中等强度运动可导致机体自噬反应激活、骨骼肌自噬小体激增[6]。然而,迄今为止,高强度力竭训练骨骼肌自噬反应特征及其机制并未得到充分验证。有研究认为,低氧条件下急性运动可能导致骨骼肌自噬抑制,这可能与骨骼肌组织对低氧的“迟滞反应”或运动性氧化应激有关[7]。Hippo/YAP/TAZ信号通路在人类肿瘤发展、骨骼肌发育修复及肌卫星细胞功能调控过程中发挥重要作用,此外有证据显示该通路与骨骼肌组织自噬存在交互作用。Hippo通路影响骨骼肌自噬通量,广泛参与骨骼肌内组织稳态、氧化损伤及修复过程,且靶向Hippo/YAP/TAZ自噬轴参与调控YAP/TAZ转录活性[8]。研究证实,自噬机制扮演了YAP/TAZ的下游靶蛋白调节器,外源性自噬抑制剂干预或内源性敲除自噬相关基因(ATG5/7/10)可抑制YAP及TAZ介导的细胞增殖能力[9]。
大量研究显示,中短期中、高强度运动可能诱发间歇性相对/绝对心肌缺血缺氧,从而增强心肌对缺血缺氧的耐受能力,定量适应运动诱导的组织保护作用称为运动预处理(Exercise preconditioning,EP),EP可有效减轻高强度急性运动可能导致的心肌缺血缺氧损伤[10]。重复性暴露于低氧可使机体组织和细胞获得对缺氧的高度耐受性,该现象称为低氧预适应(Hypoxic preconditioning,HP)[11]。EP和HP对心肌、肺组织、脑组织的保护作用及机制已基本明了,但现阶段缺乏对急性高强度运动中骨骼肌等高活性组织保护作用的研究。同时,EP和HP的作用效应存在趋同性,但其具体作用机制大相径庭,现阶段尚无证据阐明二者作用差异的分子学机制。自噬可能介导EP和HP的激活过程,但其机制仍存在较大争议。多项研究证实,HP复氧阶段通过下调Beclin1表达诱导自噬抑制现象,相反,外源性抑制自噬可能减弱EP诱导的组织细胞保护作用,表明自噬部分参与了EP和HP的组织细胞保护机制[12]。另有證据显示,Hippo及其下行通路可能与低氧暴露骨骼肌损伤修复及组织保护密切相关[13]。基于上述结论,课题组假设Hippo/YAP/TAZ信号通路作为隐匿机制,通过将EP、HP诱导的间歇性骨骼肌缺血缺氧与自噬调控联系起来,进而实现低氧力竭运动中骨骼肌的保护作用。因此,本研究拟针对低氧运动预适应大鼠低氧力竭运动后腓肠肌损伤、自噬水平及Hippo/YAP/TAZ信号通路表达进行分析,探究低氧运动预适应激活自噬调控Hippo/YAP/TAZ信号通路减轻急性低氧力竭运动大鼠骨骼肌损伤的基本机制,为运动员高原训练、移居高原人群体育锻炼及官兵高原驻训提供理论依据及参考。
1 材料与方法
1.1 动物和主要仪器、试剂
50只6周龄SPF级雄性SD大鼠,体重170±10 g,购自新疆医科大学医学实验动物中心,实验动物许可证号:SYXK(新)2016-0002。
主要仪器设备:常压低氧发生设备(Hypoxic Tent System),大鼠实验跑台(BHW-PT/5s),全自动组织切片机(KH-Q330),显微图像分析系统(NGI-2000U),PCR扩增仪(Biometra),酶标仪(Bio-Rad),高速冷冻离心机(Eppendorf),EP凝胶成像系统(Alpha),电泳系统(Bio-Red),紫外分光光度计(Eppendorf)。
主要试剂:一氧化氮、丙二醛、超氧化物歧化酶、乳酸脱氢酶、肌酸激酶、白介素-6、白介素1β、肿瘤坏死因子-α试剂盒均购自南京建成生物工程研究所,苏木素伊红(HE)染色试剂盒、Trizol总RNA抽提试剂、cDNA合成试剂盒、双链cDNA合成试剂盒、dNTP Mixture、Taq PCR MasterMix均购自上海碧云天生物公司,cDNA引物Oligo购自上海生工公司,BCA蛋白浓度测定试剂盒购自美国Proteintech公司,自噬相关5同源物(Atg5)、自噬关键分子酵母Atg6同系物(Beclin1)、微管相关蛋白1轻链3-Ⅱ(LC3-Ⅱ)、哺乳动物不育系20样激酶1(MST1)、大肿瘤抑制因子1(LATS1)、Yes相关蛋白-1(YAP)及转录共激活因子PDZ结合基(TAZ)单克隆抗体抗体购自美国Proteintech公司,Anti-β-actin内参抗体、Mouse源二抗、Rabbit源二抗、6.6×8.5 cm PVDF膜均购自上海碧云天生物公司。
1.2 动物分组及干预方案
动物于实验室条件适应性喂养3 d后,随机分为5组,每组10只,分别为:空白对照组(Blank control group,Con组)、单纯急性低氧力竭运动组(Hypoxic exercise group,HE组)、低氧预适应+急性低氧力竭运动组(Hypoxic preconditioning+Hypoxic exercise group,HP+HE组)、运动预适应+急性低氧力竭运动组(Exercise preconditioning +Hypoxic exercise group,EP+HE组)、低氧预适应+运动预适应+急性低氧力竭运动组(Hypoxic preconditioning+Exercise preconditioning+Hypoxic exercise group,EP/HP+HE组)。
预适应干预前HE组、EP+HE组及EP/HP+HE组大鼠进行跑台运动适应性训练,3 d,1次/d,15 min/次,10~12 m/min,坡度0°,运动环境为常压常氧;HE组、HP+HE组及EP/HP+HE组大鼠进行低氧环境适应性暴露,3 d,1次/d,60 min,FiO2 14.5%(海拔约3000 m),常压环境。
低氧运动预适应周期为2 w。Con组及HE组大鼠保持正常饮食及活动,不参与预适应干预方案;EP+HE组及EP/HP+HE组大鼠进行跑台训练,2次/d,早晚各1次,30 min/次,15~20 m/min,匀速渐增坡度0°~5°,運动环境为常压常氧;HP+HE组及EP/HP+HE组大鼠晚间22:00~次日晨8:00于低氧帐篷休息,FiO2 13.5%(约3 500 m海拔),常压环境。低氧运动预适应结束次日,HE组、HP+HE组、EP+HE组及EP/HP+HE组大鼠进行低氧环境跑台力竭训练,FiO2 11.9%~12.0%(约4 500 m海拔),10~20 m/min递增跑速至力竭,运动过程中采用声、频闪光及电流刺激。力竭判断标准为:①大鼠呼吸短促,无力维持既定跑速;②后肢随转动皮带后拖达30 s,经声、频闪光及电流刺激均无反应。
1.3 动物处置、取样
低氧力竭训练2 h后,麻醉、空腹状态尾静脉取血;迅速固定,取大鼠后肢腓肠肌,剔除筋膜及脂肪组织,生理盐水反复漂洗,吸水纸擦干水分,经液氮急速低温后转移至-80℃超低温冰箱保存备用。
1.4 检测方法
1.4.1 血清和腓肠肌组织匀浆制备
大鼠全血室温自凝45 min,4℃、2 000 rpm离心10 min取上清即为血清;取500 mg腓肠肌组织,剪碎,加入PBS匀浆介质,2 000 r/min匀浆2 min,匀浆液转移至离心管,4℃、6 000 r/min,离心10 min,取上清-80℃保存。
1.4.2 生化检测
取预先制备血清,采用试剂盒检测血清一氧化氮(Nitric Oxide,NO)、丙二醛(Malondialdehyde,MDA)、超氧化物歧化酶(Superoxide dismutase,SOD)、乳酸脱氢酶(Lactate dehydrogenase,LDH)及肌酸激酶(Creatine Kinase,CK)水平。
1.4.3 酶联免疫吸附实验
取预先制备血清,按照试剂盒说明书稀释标准品至10 μg/mL、加样并设空白孔和待测样品孔,37℃孵育20 min,甩干液体,PBST重复洗涤3次,待测样品孔加入酶标试剂液,37℃孵育20 min,PBST重复洗涤3次,TMB底物显色,加入终止液,450 nm处测OD值,浓度转换,以此检测白介素-6(Interleukin-6,IL-6)、白介素1β(Interleukin-1β,IL-1β)、肿瘤坏死因子-α(Tumor necrosis factor-α,TNF-α)。
1.4.4 腓肠肌组织病理染色
取大鼠腓肠肌组织,PBS冲洗,4%多聚甲醛固定24 h;梯度酒精(75%、85%、95%×2、100%×2)各脱水1 h,二甲苯透明,重复2次,各30 min;50℃~52℃浸蜡2次,各1 h;组织包埋,凝固后转移至4℃冰箱保存。沿样品横截面切片,厚度8 μm,40℃水浴展片,60℃烤片过夜。二甲苯脱蜡3次,各10 min;梯度酒精(100%×2、95%×2、85%、75%)及蒸馏水各复水5 min。Harris苏木素液浸染5 min,漂洗;1%盐酸酒精脱色5 s,饱和磷酸氢二钠溶液反蓝10 min,70%、80%酒精各固定10 min;伊红液染色30 s,烤干、封片;镜下观察,切片中心视野采集图像。
1.4.5 RT-PCR
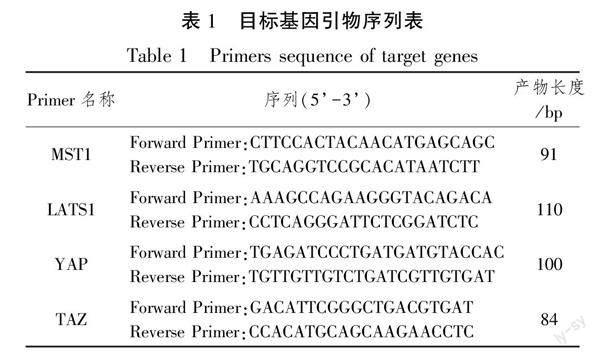
取200 μL腓肠肌匀浆液,采用Trizol法提取组织液总RNA,电泳检测完整性,分光光度仪检测RNA纯度及浓度。使用逆转录试剂盒逆转录成cDNA,采用实时荧光定量PCR(qRT-PCR)法检测mRNA相对表达水平。PCR反应体系20 μL:cDNA模板1 μL、ddH2O 7 μL、正向引物1 μL、反向引物1 μL、2×Taq PCR master mix10 μL,条件为95℃初始变性55 s;然后在95℃变性10 s,57℃退火30 s,40个循环。每组实验重复3次。根据熔解曲线剔除异常数据,导入相对定量公式(2-ΔΔCt)逐个计算样品基因的相对表达量,目标基因及其引物序列见表1。
1.4.6 蛋白印迹检测
取200 μL腓肠肌匀浆液,采用RIPA法提取组织总蛋白,4℃,15 000 g离心20 min;取上清液使用BCA蛋白浓度试剂盒测定蛋白浓度,加入5×SDS上样缓冲液,95℃变性15 min。配置SDS-PAGE凝胶,浓缩胶恒压80 V电泳30 min,分离胶恒压110 V电泳至溴酚蓝到达分离胶底部,20 μg上样,用电转法将蛋白质转移到PVDF膜上。5%脱脂牛奶室温封闭1 h,加入一抗(1[DK]∶1 000稀释),4℃摇床孵育过夜;TBST洗膜10 min×3次,加二抗(1[DK]∶3 000稀释),37℃孵育40 min;TBST洗膜10 min×5次。ECL法显影,凝胶成像系统成像后使用Quantity One3.0软件分析,以β-actin作为内参,计算目的蛋白相对表达量。
1.5 统计学分析
采用SPSS 25.0统计学软件进行统计分析,数据以平均数±标准差(MEAN±SD)表示,两样本平均数间比较采用t检验分析,多样本平均数间比较采用单因素方差分析(one way ANOVA)和两因素方差分析(one way ANOVA),组间比较采用LSD检验,P<0.05表示有显著性差异。
2 结 果
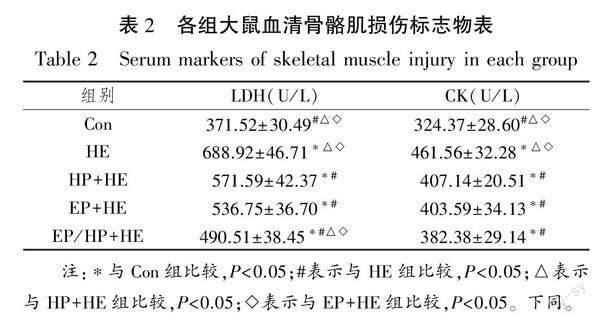
2.1 各组大鼠血清骨骼肌损伤标志物变化
本研究对各组大鼠血清骨骼肌损伤指标进行检测,结果表明(见表2),与Con组比较,低氧力竭运动各组LDH和CK水平均显著上升(P<0.05)。可见,HE组及预适应组大鼠均出现不同程度骨骼肌损伤。与HE组比较,预适应组血清LDH及CK水平均显著下降(P<0.05);预适应组间比较可知,EP/HP+HE组血清LDH显著低于HP+HE组及EP+HE组(P<0.05);EP+HE组血清LDH及CK低于HP+HE组,但比较并无统计学意义(P>0.05)。以上结果说明,急性低氧力竭运动导致大鼠骨骼肌損伤标志物水平激增,2 w低氧和/或运动预适应降低了低氧力竭后运动大鼠骨骼肌损伤。
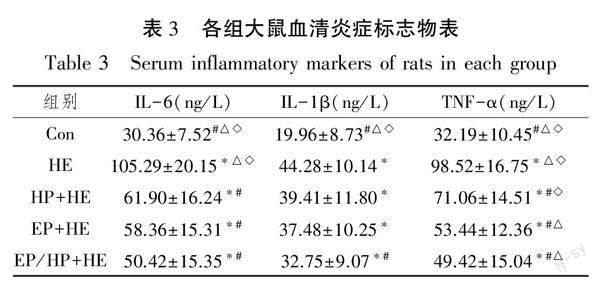
2.2 各组大鼠血清炎症因子变化
本研究检测了各组大鼠血清炎症标志物水平,结果表明(见表3),与Con组比较,低氧力竭运动各组血清IL-6、IL-1β及TNF-α均显著上升(P<0.05)。就IL-6而言,与HE组比较,预适应组均显著降低(P<0.05),预适应组内比较无统计学意义(P>0.05);就IL-1β而言,与HE组比较,EP/HP+HE组显著降低(P<0.05),HP+HE组及EP+HE组呈下降趋势,但无统计学意义(P>0.05);就TNF-α而言,与HE组比较,预适应组均显著降低(P<0.05),EP+HE组及EP/HP+HE组显著低于HP+HE组(P<0.05)。上述结果表明,急性低氧力竭运动导致大鼠血清炎症标志物水平激增,2 w低氧和/或运动预适应降低了低氧力竭运动后大鼠炎症水平。
2.3 各组大鼠血清氧化应激标志物变化
本研究检测了各组大鼠血清氧化应激标志物水平,结果表明(见表4),与Con组比较,低氧力竭运动各组血清NO、MDA均显著上升(P<0.05),血清SOD显著下降(P<0.05)。就NO而言,与HE组比较,预适应组均显著降低(P<0.05),EP/HP+HE显著低于HP+HE组及EP+HE组(P<0.05),可见低氧+运动预适应对血清NO的影响显著效果高于单一预适应;就MDA而言,与HE组比较,预适应组均显著降低(P<0.05),EP/HP+HE组显著低于HP+HE组及EP+HE组(P<0.05),EP+HE组显著低于HP+HE组(P<0.05);就SOD而言,与HE组比较,预适应组均显著上升(P<0.05),EP/HP+HE组显著高于HP+HE组(P<0.05)。上述结果表明,急性低氧力竭运动导致大鼠血清氧化应激标志物水平明显波动,2 w低氧和/或运动预适应降低了低氧力竭运动后大鼠氧化应激水平,低氧+运动预适应对低氧力竭运动后大鼠血清氧化应激标志物的影响可能好于单一预适应。
2.4 各组大鼠腓肠肌组织微细结构病理变化
Con组大鼠腓肠肌肌纤维结构正常,呈多边形,排列有序,细胞核分布于肌细胞边缘,颜色、边界及形态均正常;HE组肌纤维肿胀、碎解,炎症组织浸润明显,组织间隙散布不均,细胞排列混乱。预适应组较HE组肌纤维肿胀及炎症组织浸润减轻,细胞形态及边界组织趋于正常。此外,预适应各组镜下比较可知,EP+HE组、EP/HP+HE组肌纤维肿胀及炎症组织浸润症状减轻,肌细胞及组织边界清晰(见图1)。
2.5 各组大鼠腓肠肌自噬相关蛋白表达的变化
本研究对各组大鼠骨骼肌自噬相关蛋白Atg5、Beclin1及LC3-Ⅱ进行检测,结果发现(见图2),与Con组比较,低氧力竭运动组大鼠腓肠肌Atg5及Beclin1蛋白呈下降趋势,LC3-Ⅱ蛋白呈上升趋势。就Atg5蛋白表达而言,与Con组比较,低氧力竭运动各组均显著下降(P<0.05);与HE组比较,EP+HE组及EP/HP+HE组显著上升(P<0.05);EP/HP+HE组显著高于HP+HE组(P<0.05)。就Beclin1蛋白表达而言,与Con组比较,HE组及HP+HE组显著降低(P<0.05);与HE组比较,EP+HE组及EP/HP+HE组显著上升(P<0.05);EP/HP+HE组显著高于HP+HE组(P<0.05)。就LC3-Ⅱ蛋白表达而言,与Con组比较,HE组及HP+HE组显著上升(P<0.05);与HE组比较,EP+HE组及EP/HP+HE组显著降低(P<0.05);EP+HE组及EP/HP+HE组显著高于HP+HE组(P<0.05)。上述结果表明,急性低氧力竭运动导致大鼠腓肠肌组织自噬水平被抑制,2 w低氧和/或运动预适应提高了低氧力竭运动后大鼠腓肠肌自噬水平,低氧+运动预适应对低氧力竭运动后大鼠腓肠肌自噬的影响可能好于单一预适应。
2.6 各组大鼠腓肠肌Hippo/YAP/TAZ信号通路相关mRNA表达的变化
本研究检测了各组大鼠腓肠肌MST1、LAST1、YAP及TAZ mRNA水平,结果发现(见图3),与Con组比较,低氧力竭运动组MST1、LATS1 mRNA显著降低(P<0.05),YAP及TAZ mRNA显著上升(P<0.05)。就MST1 mRNA表达而言,与HE组比较,EP+HE组及EP/HP+HE组显著上升(P<0.05);EP+HE组及EP/HP+HE组显著高于HP+HE组(P<0.05)。就LATS1蛋白表达而言,与HE组比较,预适应组均显著上升(P<0.05)。就YAP蛋白表达而言,与HE组比较预适应组均显著降低(P<0.05);EP/HP+HE组显著低于HP+HE组(P<0.05)。就TAZ蛋白表达而言,与HE组及HP+HE比较,EP+HE组及EP/HP+HE组均显著降低(P<0.05)。上述结果表明,急性低氧力竭运动抑制大鼠腓肠肌组织MST1、LAST1 mRNA表达,同时上调YAP及TAZ mRNA表达;2 w低氧和/或运动预适应提高了低氧力竭运动后大鼠腓肠肌组织MST1、LAST1 水平并下调YAP、TAZ mRNA表达。
2.7 各组大鼠腓肠肌Hippo/YAP/TAZ信号通路相关蛋白表达变化
本研究对各组大鼠骨骼肌Hippo/YAP/TAZ信号通路蛋白MST1、LATS1、YAP及TAZ进行检测,结果发现(见图4),与Con组比较,低氧力竭运动组MST1、LATS1蛋白及其磷酸化呈下降趋势,YAP及TAZ蛋白呈上升趋势。就MST1蛋白表达而言,与Con组比较,HE组、HP+PE组及EP+HP组均显著下降(P<0.05);与HE组比较,EP+HE组及EP/HP+HE组显著上升(P<0.05);EP+HE组及EP/HP+HE组显著高于HP+HE组(P<0.05)。就p-MST1蛋白表达而言,与Con组比较,低氧力竭运动组显著降低(P<0.05);与HE组比较,EP+HE组及EP/HP+HE组显著上升(P<0.05);EP+HE组及EP/HP+HE组显著高于HP+HE组(P<0.05)。就LATS1蛋白表达而言,与Con组比较,HE组及HP+HE组显著降低(P<0.05);与HE组比较,预适应组均显著上升(P<0.05);EP+HE组及[CM(23]EP/HP+HE组显著高于HP+HE组(P<0.05)。就p-LATS1蛋白表达而言,与Con组比较,HE组及HP+HE组显著降低(P<0.05);与HE组比较,预适应组均显著上升(P<0.05);EP/HP+HE组显著高于HP+HE组(P<0.05)。就YAP蛋白表达而言,与Con组比较,低氧力竭运动组均显著上升(P<0.05);与HE组比较,EP+HE组及EP/HP+HE组均显著降低(P<0.05)。就TAZ蛋白表达而言,与Con组比较,HE组、HP+HE组及EP+HE组均显著上升(P<0.05);与HE组比较,预适应组均显著降低(P<0.05),EP/HP+HE组均显著低于HP+HE组(P<0.05)。上述结果表明,急性低氧力竭运动抑制大鼠腓肠肌组织MST1、LAST1蛋白及磷酸化水平,同时导致 Hippo/YAP/TAZ通路下游蛋白YAP、TAZ表达上升;2 w低氧和/或运动预适应提高了低氧力竭运动后大鼠腓肠肌组织MST1、LAST1蛋白及磷酸化水平并下调YAP、TAZ蛋白表达。
3 讨 论
3.1 EP+HP对骨骼肌损伤、炎症反应和氧化应激的影响
研究证实,急性剧烈运动导致机体广泛炎症反应、氧化应激和骨骼肌损伤,低氧环境可能进一步强化机体的应激反应[14]。IL-6是免疫级联中重要的基础免疫因子,大强度急性运动后,肌肉细胞产生的IL-1β可能通过诱导蛋白质降解参与肌肉适应调控,IL-6刺激中性粒细胞脱颗粒及皮质醇生成。TNF-α可由嗜中性粒细胞、活化的淋巴细胞、巨噬细胞、NK细胞、LAK细胞、星形胶质细胞、平滑肌细胞和部分转化细胞产生,TNF-α以分泌的可溶形式和膜锚定形式存在,短时间高强度运动可刺激IL-6、TNF-α及IL-1β大量分泌[15]。王成美等认为[16],单次大强度运动可导致机体血清TNF-α、IL-6激增。Dufaux等[17]亦证实,2~5 h跑测验和5 km赛跑后均可见血清TNF-α、IL-1β升高。血清CK、LDH是骨骼肌损伤的早期标志物,广泛存在于体内组织器官及血清,对运动应激反应敏感度较高,在高强度运动后参与诱导骨骼肌损伤。此外,为进一步探究低氧高强度运动对骨骼肌损伤、炎症反应和氧化应激的影响,本研究设置了氧浓度11.9%~12.0%(约4 500 m海拔)环境的力竭跑台训练,结果发现,低氧力竭运动后大鼠腓肠肌结构损伤、炎症反应和氧化应激反应与单纯力竭训练相似,且低氧环境加剧了大鼠腓肠肌结构损伤、炎症反应和氧化应激现象。此外本研究亦发现,2 w EP或HP明显减轻了大鼠低氧力竭运动后的腓肠肌结构损伤、炎症反应和氧化应激现象,且EP+HP对骨骼肌损伤的预防作用可能优于单纯HP或EP。以往的研究认为,在组织细胞保护效应上,EP与HP都可诱导机体产生内源性保护效应,且EP诱导机体产生的内源性保护效应与HP相似,HP和EP干预可能分别改善机体低氧耐受及骨骼肌性能[18]。但上述机制并非互相独立,机体对低氧条件的短期适应可能形成通气能力上升、骨骼肌微血管增生、骨骼肌代谢底物增加、低氧缓冲能力改善等优势,运动预适应也可能获得类似适应现象。因此,EP+HP可能形成骨骼肌缺氧、机械负荷的累积效应,在早期的低氧运动训练中,EP+HP功能性适应的保护效应增强,机体产生结构性适应并改善骨骼肌耐低氧、耐负荷能力,从而减轻急性低氧力竭运动后骨骼肌损伤、炎症反应和氧化应激症状。对于EP和HP的保护目标而言,现有的研究集中于心肌、肺、脑、肾、肠粘膜等器官组织,极少有研究涉及骨骼肌保护效应,本研究表明,单纯或复合形式的2 w EP和HP可以作为避免急性低氧高强度训练骨骼肌损伤及炎症、氧化应激反应的措施,但EP和HP配合干预的周期、强度、频率、形式等因素仍不明确,有赖于进一步实验证实。
3.2 EP+HP对骨骼肌自噬蛋白的影响
低氧是诱导细胞氧化应激反应的强刺激剂,同时也是激活骨骼肌自噬的重要外源性因素。骨骼肌自噬可清除受损线粒体,同时抑制线粒体产生ROS,维持细胞内稳态,有研究证实自噬可能参与EP或HP组织器官的适应过程[19]。Atg5、Beclin-1和LC3-Ⅱ是机体自噬与凋亡过程的重要信号分子,Atg5参与介导自噬体的形成,而Beclin-1通过组成PtdIns3KC3复合体诱导细胞自噬,Atg5可能介导了LC3-Ⅱ和Beclin-1激活所诱发的细胞凋亡过程[20]。因此,本研究以骨骼肌自噬为切入点,探讨EP+HP大鼠腓肠肌自噬的变化规律。自噬是一种溶酶体依赖性降解過程,在细胞能量平衡和细胞器更新中发挥重要作用。研究证实,自噬可能通过禁食、缺血/再灌注(I/R)、低氧暴露或运动等方式激活[21]。LIU等发现,晚期EP可通过激活自噬减轻力竭性运动诱导的心肌损伤[22],HUANG等亦证实EP上调Beclin1表达并诱导自噬形成早期心肌保护效应[23]。有研究显示,长期低氧运动导致自噬效应增强进而发挥组织保护效应。本研究结果表明,低氧力竭运动组导致运动后大鼠腓肠肌Atg5、Beclin1表达降低并上调了LC3-Ⅱ蛋白表达。研究认为,高强度运动后12 h内骨骼肌自噬体可能存在抑制期,可能与骨骼肌微管丢失有关[24],该结果基本佐证了本研究结论。前期研究显示,急性高强度运动诱导的氧化应激和ATP耗竭可能导致骨骼肌组织Beclin1表达上调,Beclin1-Bcl-2复合体裂解是诱导自噬的关键机制[25]。低氧条件下力竭训练骨骼肌总负载量过高,可能抑制Beclin1-Bcl-2复合体裂解并降低Beclin1表达。此外,本研究结果显示,与HE组比较,预适应组大鼠低氧力竭运动后腓肠肌Atg5、Beclin1表达上升,LC3-Ⅱ蛋白表达下降,因此,EP、HP或其累积结果导致自噬活动增强。据报道,Beclin1及其激活物Atg5的上调与AMPK、PI3K、ULK1和mTOR的上游介导有关,这些标志物先前已被证明参与EP和HP诱导的组织保护作用[26]。EP、HP诱导的自噬效应促进Beclin1-Bcl-2复合物裂解,Beclin1蛋白表达上升,I/R或单纯低氧暴露也表现出类似结果[27]。本研究中大鼠腓肠肌自噬蛋白Atg5、LC3-Ⅱ和Baclin1的变化情况表明,EP、HP可代偿性上调自噬通量水平,进而发挥其降解氧化损伤蛋白、ROS和衰老细胞器等亚细胞组件的功效,进而减轻低氧力竭运动后骨骼肌损伤、炎症和氧化应激反应。
3.3 EP+HP對骨骼肌Hippo/YAP/TAZ信号通路的影响
现有研究证据表明,Hippo/YAP/TAZ通路是一条重要的细胞内信号通路,与广泛的生理事件有关,包括细胞增殖、迁移及自噬的调节[28]。在哺乳动物中,Hippo信号通路的主要功能是抑制转录调节因子YAP和TAZ的活性,其核心组成元件为MST1和LATS1[29]。运动可以通过改变靶向基因的表达激活Hippo/YAP/TAZ通路,同时该通路的激活程度受运动负荷量的影响,如,有研究显示,EP可导致小鼠血管内皮生长因子、JG12和Bcl-2蛋白表达显著增加,Bax、Caspase 3、IL-6和衰老细胞的表达降低,导致MST1及其下游信号分子LAST1上调,进而下调YAP及TAZ表达[30]。另有研究证实,高强度运动训练尤其是高强度急性运动可能导致Hippo/YAP/TAZ通路抑制现象[31]。研究表明,YAP和TAZ活性通过上游调节器MST1、LATS1负反馈转录机制控制[32]。动物实验证实,急性力竭运动和短期低氧暴露(12.5%,48 h)可诱导骨骼肌YAP、TAZ表达上调,这表明骨骼肌在高强度运动负荷作用下出现降解效应[33]。此外,Duchenne肌营养不良mdx模型小鼠腓肠肌总YAP、TAZ及其磷酸化水平也呈上升趋势,因此运动、低氧或异常骨骼肌肥大均导致骨骼肌MST1、LAST1蛋白及其下游标志物YAP、TAZ表达异常[34]。该结果佐证了本研究中低氧力竭训练大鼠腓肠肌Hippo/YAP/TAZ通路标志物变化特征。上述结论与本研究结果基本一致,急性低氧力竭运动抑制大鼠腓肠肌组织MST1、LAST1蛋白及磷酸化水平,同时导致下游蛋白YAP、TAZ表达上升。
本研究发现,EP、HP提高了低氧力竭运动后大鼠腓肠肌组织MST1、LAST1蛋白及磷酸化水平并下调YAP、TAZ蛋白表达。现阶段研究证实,Hippo信号通路可能参与介导AKT/AMPK/mTOR级联反应,Hippo激酶MST1可结合并抑制Akt1表达活性,Akt也可反向激活MST1磷酸化,表明Akt1和MST1存在双向调节机制[35]。此外,Hippo通路效应器YAP通过Pten靶向抑制磷酸酶Pten及mTOR表达[36]。总之,上述研究表明Hippo和mTOR介导的骨骼肌功能调节信号通过多种机制紧密耦合。AMPK在机体内扮演细胞能量代谢“调节器”角色,在氧化应激或极端条件下对细胞能量代谢的适应性调节发挥重要作用,mTOR信号通路是蛋白合成的主要通路,有学者猜测mTOR可能是调控骨骼肌内组织代谢的核心靶点[37]。EP+HP累积效应可能导致肌内AMP/ATP比值及LKB1活性紊乱,AMPK-TSC2信号级联反应激活,同时诱导mTOR活性上升,进而下调肌内YAP及TAZ表达。但目前的研究证据尚无法充分解释Hippo/YAP/TAZ信号通路是否在规律性运动中发挥骨骼肌功能调控作用,但EP及HP可能通过AKT/AMPK/mTOR-Hippo/YAP/TAZ逆向调节发挥低氧力竭运动中骨骼肌整合作用。此外,多项研究证实,自噬激活现象抑制了YAP及TAZ表达[38],本研究也发现了类似的现象。mTORC1信号是自噬途径主要的上游抑制通路,而在TSC1缺失的细胞中,mTORC1通路则维持组成型激活状态[39]。LIANG等研究表明,在敲除TSC1后,细胞自噬及Hippo通路下游物均显著抑制[40]。有文献报道,YAP及TAZ蛋白可以与自噬途径的受体蛋白Beclin1交互影响,进而被Beclin1呈递至自噬体内经由溶酶体降解[41]。因此,EP+HP可能通过激活骨骼肌自噬下调低氧力竭运动大鼠YAP、TAZ表达,并可能参与AKT/AMPK/mTOR级联反应调控骨骼肌损伤、炎症反应和氧化应激反应。
4 结 论
低氧和/或运动预适应可有效减轻急性低氧力竭运动导致的氧化应激、炎症反应及骨骼肌损伤;低氧和/或运动预适应可通过激活自噬反应调控骨骼肌Hippo/YAP/TAZ信号通路并发挥急性低氧力竭运动中的骨骼肌保护机制。
参考文献:
[1]李晓智,高亮.急性运动对机体免疫系统影响的Meta分析[J].西安体育学院学报,2020,37(3):353-363.
[2] 周文婷,胡扬,徐飞.急性低氧暴露与低氧运动习服的血液学评价指标[J].中国应用生理学杂志,2018,34(6):509-512.
[3] 徐玉明,曹建民,陈玉平,等.低氧调控大鼠运动性肌损伤的特征MicroRNA表达[J].中国运动医学杂志,2016,35(10):913-920.
[4] LUCA R,RYAN H,ALEXANDER H,et al.High-altitude acclimatization improves recovery from muscle fatigue[J].Med Sci Sports Exerc,2020,52(1):161-169.
[5] MOOREN C,KRUGER K.Exercise, autophagy, and apoptosis[J].Prog Mol Biol Transl,2015,135(7):407-422.
[6] 李梦影,李灵杰,马春伟,等.运动通过脂噬作用调节脂代谢及其分子机制[J].生理学报,2022,74(2):309-319.
[7] MARCUS M,WILLIAM A,OSCAR H,et al.Acute normobaric hypoxia blunts contraction-mediated mTORC1- and JNK-signaling in human skeletal muscle[J].Acta Physiol,2022,234(2):1-19.
[8] UL D,LAURENCE B,ANDREW P,et al.Osimertinib-resistant NSCLC cells activate ERBB2 and YAP/TAZ and are killed by neratinib[J].Biochem Pharmacol,2021,190(9):642-651.
[9] JI L,LI X,ZHOU Z,et al.LINC01413/hnRNP-K/ZEB1 axis accelerates cell proliferation and EMT in colorectal cancer via inducing YAP1/TAZ1 translocation[J].Mol Ther Nucleic Acids,2020,19(3):546-561.
[10] 万栋峰,潘珊珊,原阳.心肌线粒体自噬相关蛋白BNIP3在运动预适应晚期保护效应中的变化[J].体育科学,2017,37(3):35-43.
[11] 李海英,苗卫国,马静芬,等.线粒体自噬在运动预适应抗急性低压低氧大鼠脑海马损伤中的作用[J].中国运动医学杂志,2014,33(6):524-529.
[12] 孙其龙,潘珊珊,黄悦,等.运动预适应通过间歇性缺血缺氧诱导细胞自噬参与早期心肌保护的研究[J].体育科学,2018,38(7):25-32,49.
[13] ZHAO K,SHEN C Y,LU Y S,et al.Muscle yap is a regulator of neuromuscular junction formation and regeneration[J].J Neurosci,2017,37(18):4859.
[14] 高照,杨杏萍,张援,等.运动后氧化应激水平及合理补充抗氧化剂的科学评价方法[J].中国组织工程研究,2017,21(28):4577-4584.
[15] JAAK J,SILLE V,PRIIT P.Circulating inflammatory cytokine responses to endurance exercise in female rowers[J].Int J Sports Med,2018,39(14):1041-1048.
[16] 王成美,曾红兵.高强度运动诱发肌肉损伤对血糖调节、肌酸激酶、IL-6、TNF-α水平的影响[J].基因组学与应用生物学,2018,37(3):970-975.
[17] DUFAUX B,ORDER U,LIESEN H.Effect of a short maximal physical exercise on coagulation,fibrinolysis,and complement system[J].Int J Sports Med,1991,12(1):38-42.
[18] ZHANG H,LIU M Y,KIM H,et al.Preconditioning improves muscle regeneration after ischemia-reperfusion injury[J].J Orthop Res,2021,39(9):1889-1897.
[19] LU N,LI X X,TAN R L,et al.HIF-1α/Beclin1 mediated autophagy is involved in neuroprotection induced by hypoxic preconditioning[J].J Mol Neurosci,2018,66(2):238-250.
[20] CHEN J,ZHANG L,ZHOU H,et al.Inhibition of autophagy promotes cisplatin-induced apoptotic cell death through Atg5 and Beclin 1 in A549 human lung cancer cells[J].Mol Med Rep,2018,17(5):6859-6865.
[21] 方夢蝶,刘波,刘伟.自噬的分子细胞机制研究进展[J].中国细胞生物学学报,2012,34(4):382-390.
[22] LIU H T,PAN S S.Late exercise preconditioning promotes autophagy against exhaustive exercise induced myocardial injury through the activation of the AMPK-mTOR-ULK1 pathway[J].Biomed Res Int,2019,7(11):1-10.
[23] HUANGY,LIU H T,YUAN Y,et al.Exercise preconditioning increases Beclin1 and induces autophagy to promote early myocardial protection via intermittent myocardial ischemia hypoxia[J].Int Heart J,2021,62(2):407-415.
[24] 王祯,雷槟恺,于亮,等.急性离心运动后恢复期大鼠骨骼肌微管形态及自噬体降解特征研究[J].中国康复医学杂志,2022,37(2):148-154.
[25] HE C, BASSIK C, MORESI V,et al.Exercise-induced BCL2-regulated autophagy is required for muscle glucose homeostasis[J].Nature,2012,481(7382):511-515.
[26] 阮雯静,万福生.Beclin1-Vps34在自噬发生发展中的作用[J].中国细胞生物学学报,2016,38(11):1420-1426.
[27] SHENG R,QIN Z H.The divergent roles of autophagy in ischemia and preconditioning[J].Acta Pharmacol Sin,2015,36(4):411-420.
[28] YUAN P,HU Q Y,HE X M,et al.Laminar flow inhibits the Hippo/YAP pathway via autophagy and SIRT1-mediated deacetylation against atherosclerosis[J].Cell Death Dis,2020,11(2):1-11.
[29] 陈林云,欧阳家和,段琼,等.YAP/TAZ的调控机制研究进展[J].生命科学,2020,32(12):1381-1390.
[30] AMIN A,KATHRIN M.Deathly triangle for pancreatic β-cells: Hippo pathway-MTORC1-autophagy[J].Autophagy,2021,17(12):4494-4496.
[31] BRENDAN G,LEE H,Annie T,et al.The Hippo signal transduction network for exercise physiologists[J].J Appl Physiol,2016,120(10):1105-1117.
[32] MA S H,MENG Z P,CHENR,et al.The Hippo pathway: biology and pathophysiology[J].Annu Rev Biochem,2019,88(6):577-604.
[33] MARIANA P,MAURIZIO R,JUNG P,et al.Contact inhibition controls cell survival and proliferation via YAP/TAZautophagy axis[J].Nat Commun,2018,9(1):2961-2968.
[34] GIAN V,FRANCESCA P,ROSARIA O,et al.Hippo signaling pathway is altered in Duchenne muscular dystrophy[J]. PLoS One,2018,13(10):42-49.
[35] NEGAR N,BENTOLHODA H,MARZIEH J,et al.The Hippo signaling pathway in leukemia:function, interaction, and carcinogenesis[J].Cancer Cell Int,2021,21(1):705-712.
[36] GAN W J,DAI X M,DAI X P,et al.LATS suppresses mTORC1 activity to directly coordinate Hippo and mTORC1 pathways in growth control[J].Nat Cell Biol,2020,22(2):246-256.
[37] FARNAZ F,MARK P,SEMONE M.Creatine supplementation and skeletal muscle metabolism for building muscle mass- review of the potential mechanisms of action[J].Curr Protein Pept Sci,2017,18(12):1273-1287.
[38] 詹永通,吳桂豪,范旭红,等.Hippo-YAP信号通路调控细胞衰老的分子机制[J].中国病理生理杂志,2022,38(5):920-931.
[39] 许银丰,汪倩,钱楚莹,等.mTORC1信号通路调控细胞自噬的研究进展[J].中国科学(生命科学),2022,52(2):266-272.
[40] LIANG N,ZHANG C,PATRICIA D,et al.Regulation of YAP by mTOR and autophagy reveals a therapeutic target of tuberous sclerosis complex[J].J Exp Med,2014,211(11):2249-2263.
[41] TANG F Y,GERHARD C.LATS1-Beclin1 mediates a non-canonical connection between the Hippo pathway and autophagy[J].Mol Cell Oncol,2020,7(4):51-58.
收稿日期:2022-11-04
基金项目:国家自然科学基金项目(编号:31660736);新疆维吾尔自治区高校科技计划项目(编号:XJEDU2021SY040)。
作者简介:孔海军(1992- ),男,山东济宁人,博士研究生,讲师,研究方向为运动生物化学。
通讯作者:谌晓安(1974- ),女,湖南张家界人,博士,教授,博士研究生导师,研究方向为运动人体科学。
猜你喜欢
中国医药导报(2017年2期)2017-03-18
中国中药杂志(2017年1期)2017-03-06
医学信息(2016年34期)2017-02-22
中国中药杂志(2016年21期)2017-02-16
中国中药杂志(2016年21期)2017-02-16
糖尿病新世界(2016年16期)2016-12-09
中西医结合心血管病电子杂志(2016年6期)2016-11-14
科技视界(2016年11期)2016-05-23
山东体育学院学报(2015年2期)2015-05-27
