
植物镉毒害与硒对镉胁迫的缓解作用研究进展
2023-06-28 01:32翟夜雨黄五星袁岐山陈小龙陈洋杨金初邵惠芳
河南农业大学学报 2023年3期
翟夜雨, 黄五星, 袁岐山, 陈小龙, 陈洋, 杨金初, 邵惠芳
(1.河南农业大学烟草学院,河南 郑州 450002;2.河南中烟工业有限责任公司,河南 郑州 450016)
采矿、电镀、冶金、废物燃烧及使用含镉农药和化肥等往往导致土壤镉污染[1-2]。土壤中的镉具有较强毒性,半衰期长且极难分解转化。镉被植物根系吸收积累到阈值后,会抑制植物的生长发育和生殖进程,出现如根系发育缓慢或停滞、叶片卷曲变黄失绿、花粉可育性下降等症状,严重时导致植物死亡[3]。因此,寻找一种能够有效缓解镉对农作物毒害作用的方法已成为农业和环境科研工作者关注的焦点。近年来,越来越多的研究发现,低剂量的硒能够提高植物抗氧化能力和抗逆能力,缓解重金属等非生物胁迫。在此,本研究从镉胁迫对植物生长发育的影响(镉对植物光合作用的抑制、镉对植物养分吸收的抑制和镉诱发植物氧化损伤)以及硒对植物镉胁迫的缓解作用(硒降低植物细胞中镉含量、硒改善镉胁迫植物光合作用、硒调节镉胁迫植物矿质元素平衡和硒增强镉胁迫植物抗氧化能力)综述相关研究进展。
1 镉胁迫对植物生长的抑制
1.1 镉对植物光合作用的抑制
作为自养生物,植物通过光合作用吸收光能,把二氧化碳和水合成有机物以维持生长与代谢。光合作用主要在叶绿体内进行,涉及光吸收、电子传递、光合磷酸化、碳同化等步骤,可分为光反应和暗反应两个阶段。镉可以通过影响叶绿体结构,光合色素含量和光合作用过程抑制植物光合速率[4]。叶绿体结构对镉极为敏感,5 mg·L-1镉胁迫下玉米叶绿体会出现基质片层及基粒和类囊体片层排列紊乱[5]。100 μmol·L-1镉胁迫下小麦叶绿体基粒和类囊体片层结构出现紊乱,光捕获效率和光合电子传递效率显著降低[6-7]。
植物叶绿体中的光合色素主要包括叶绿素a、叶绿素b、胡萝卜素和叶黄素。在小白菜、黄瓜、茶树、大豆等[8-11]植物上研究发现,叶绿素含量随镉胁迫浓度的增加而大幅下降。并且叶绿素a比叶绿素b对镉更敏感,镉胁迫使叶绿素a/叶绿素b的比值下降。镉胁迫减少叶绿素含量的可能原因:一方面是镉干扰构成叶绿素关键元素铁的代谢,植物有效铁含量降低,叶绿素合成受到限制。例如李亮等[12]研究表明,镉胁迫下紫苏中铁含量降低29.3%。另一方面是镉取代叶绿素中Ca2+和Mg2+的结合位点,破坏正常叶绿素结构,降低叶绿素含量。相对于叶绿素,镉胁迫对胡萝卜素和叶黄素影响的研究十分有限。
从光合作用过程上看,镉胁迫对光吸收、电子传递和碳同化步骤都可以产生抑制。在光吸收步骤,镉可与捕光色素蛋白复合体上的亚基发生交互作用,抑制光吸收效率。在电子传递步骤,光系统Ⅱ比光系统I对镉更为敏感。罗丽娟等[13]研究表明,镉胁迫显著降低烟草叶片中PSⅡ电子传递量子效率,并显著改变单位PSⅡ反应中心数量。王竹承等[14]发现,镉胁迫降低马齿苋叶片PSⅡ电子传递效率,但并未改变PSⅡ反应中心最大活性。镉胁迫对碳同化的抑制,一方面是在暗反应阶段干扰卡尔文循环羧化和再生阶段酶的活性。例如,镉胁迫使酸枣[15]和裸燕麦[16]中核酮糖1, 5-二磷酸羧化酶和磷酸烯醇式丙酮酸羧化酶活性降低。另一方面是镉胁迫降低气孔导度影响碳同化效率。例如,镉胁迫下甜高粱、野苋菜和油菜气孔导度显著下降[17-19]。而气孔关闭的原因被认为与保卫细胞中钾、钙和脱落酸代谢被镉胁迫干扰有关。
1.2 镉对植物养分吸收的抑制
植物主要通过根系吸收矿质元素,并运输至各个器官中,维持体内生理代谢过程[20]。镉胁迫可抑制多种矿质元素的吸收、运输和分配[21-22]。例如,在大麦、番茄、苹果和紫花苜蓿等[23-26]植物中研究发现,镉胁迫显著抑制K、Mg、Fe、Ca、Zn和Mn的吸收,导致植物生长的停滞。此外,在氮素吸收过程中,镉通过抑制硝酸还原酶或亚硝酸还原酶的活性,降低根系对硝酸盐的吸收及其向地上部分的运输[27]。郭智等[28]研究表明,龙葵幼苗根系中硝态氮含量和硝酸还原酶活性随镉胁迫时间延长逐渐下降。
镉胁迫抑制矿质养分吸收的可能原因:第一,根系长期接触镉后根尖根尖区表皮和皮层细胞的分裂紊乱和异常增大,侧根数目减少并腐解,导致根系活力降低和形态的改变,主根呈棕褐色,并造成元素吸收效率降低。第二,根系对矿质元素的吸收受质膜选择特性的影响,镉可以通过改变细胞膜的通透性或影响元素在膜上的转运过程来干扰矿质养分的吸收[29]。研究表明,H+-ATP酶是质膜上的一类转运蛋白,主要分布在根系的表皮和皮层。H+-ATP在质膜上起着离子转运体的作用,这依赖于质膜H+-ATP所产生的电化学梯度。镉处理对玉米根细胞H+-ATP酶活性有抑制作用[30]。因此,镉导致H+-ATP酶活性降低并抑某些元素的吸收。此外,镉与植物必需二价阳离子结构相近,在吸收通道或金属转运体间存在竞争关系,会导致必要矿质养分的缺乏。TIAN等[31]通过转录组对水稻重金属P1b-ATP酶(HMAs)分析表明,镉与水稻组织中微量元素锌、铜对HMAs竞争,导致地上部分的矿质元素的缺乏。
1.3 镉诱发植物氧化损伤
活性氧(过氧化物、超氧化物、羟基自由基、单线态氧等)是光合和呼吸作用的副产物,在植物细胞内发挥信号转导或翻译后修饰的功能,如控制细胞程序性死亡、激活抗氧化防御系统等[32]。植物自身通过抗氧化防御系统,例如酶促系统(过氧化物酶、超氧化物歧化酶、抗坏血酸、过氧化物酶和过氧化氢酶等)和非酶抗氧化物(抗坏血酸、还原型谷胱甘肽、类胡萝卜素等)[33],清除过量的活性氧物质。
镉是一种非氧化还原金属,并不产生过氧根离子、过氧化氢、羟基自由基和丙二醛等,而是通过干扰植物活性氧清除系统,直接或间接导致植物体内活性氧的过量积累[34],并造成植物的氧化损伤。活性氧损伤主要包括植物细胞膜脂过氧化、蛋白质过氧化和脱氧核糖核酸及碱基损伤,并影响整个植物的发育[35]。例如,崔宏莉等[36]在菠菜镉胁迫中发现,随着镉处理浓度的增大,SOD和POD等活性都有先升高后降低的趋势,当胁迫程度超过菠菜正常抗逆能力时,细胞内多种抗氧化酶和膜结构均遭到破坏,造成植物生长停滞。韩航等[37]研究证实镉胁迫下类芦体内抗氧化酶系统受到抑制,且POD、CAT和SOD活性随镉含量的升高而呈明显下降趋势,导致类芦生理代谢紊乱,不能正常生长。同样,刘伟等[10]研究表明,随着镉处理浓度的增加,在黄瓜叶片中的过氧化物酶、过氧化氢酶活性显著下降,并导致细胞膜脂过氧化的产物丙二醛的含量显著增加。关于镉影响抗氧化酶活性的作用机制,有研究显示,镉在高浓度水平下,可能与金属元素 Ca、Fe、Zn等竞争结合抗氧化酶生物活性中心,致使其空间结构变化[38],最终导致抗氧化功能失调,与此过程相关的研究十分有限。图1表示镉胁迫对植物生长的影响。
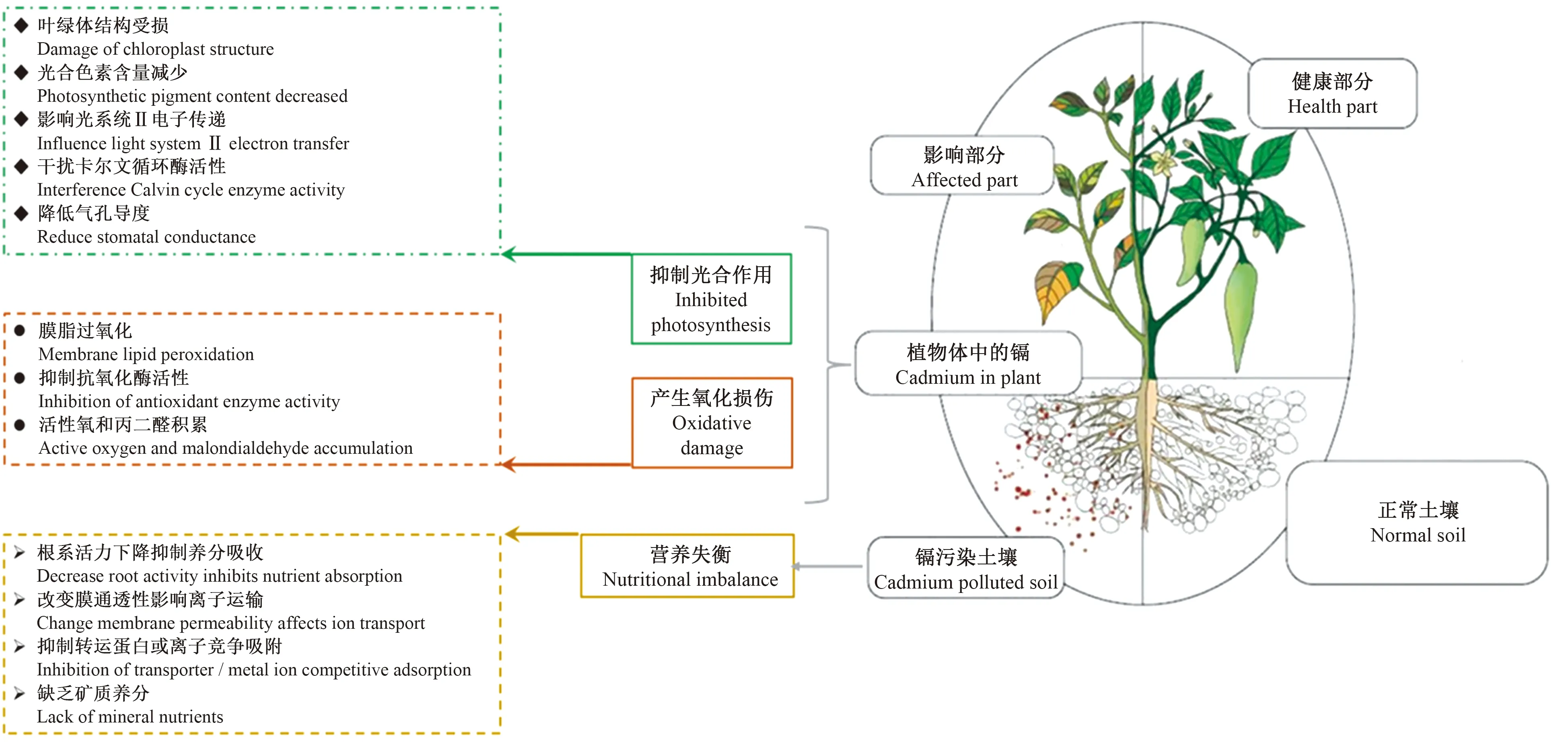
图1 镉胁迫对植物生长的影响
2 硒对植物镉胁迫的缓解作用
尽管硒对高等植物来说不是必需的[39],但大量研究证实,适宜浓度的硒对植物生长以及提高非生物胁迫的耐受性具有积极影响[40]。近年来,硒在减轻植物重金属(例如镉等)毒性方面的作用越来越受到关注。表1汇总了不同形态和浓度硒对植物镉胁迫的缓解作用[41-52]。周健等[53]研究表明,在小白菜中添加硒代甲硫氨酸、硒酸盐和亚硒酸盐均可有效提高抗氧化酶活性,并激活谷胱甘肽代谢途径,最终降低胁迫下小油菜的氧化损伤。徐境懋等[54]研究发现,亚硒酸盐处理可显著降低细胞中可溶物质组分和细胞器组分的镉含量,通过改变根系镉的亚细胞分布,减少镉向地上部的转运,并降低叶片膜脂过氧化程度。而张立等[55]研究表明,硒增强镉在细胞壁的固定和液泡中的螯合量,缓解镉对油菜生物量及光合色素含量的不利影响。图2总结了硒对植物镉胁迫缓解机制。下文将从硒降低植物细胞中硒含量、硒改善植物光合作用、硒调节植物矿质元素平衡和硒增强植物抗氧化能力4个方面阐述硒对植物镉胁迫的缓解作用。
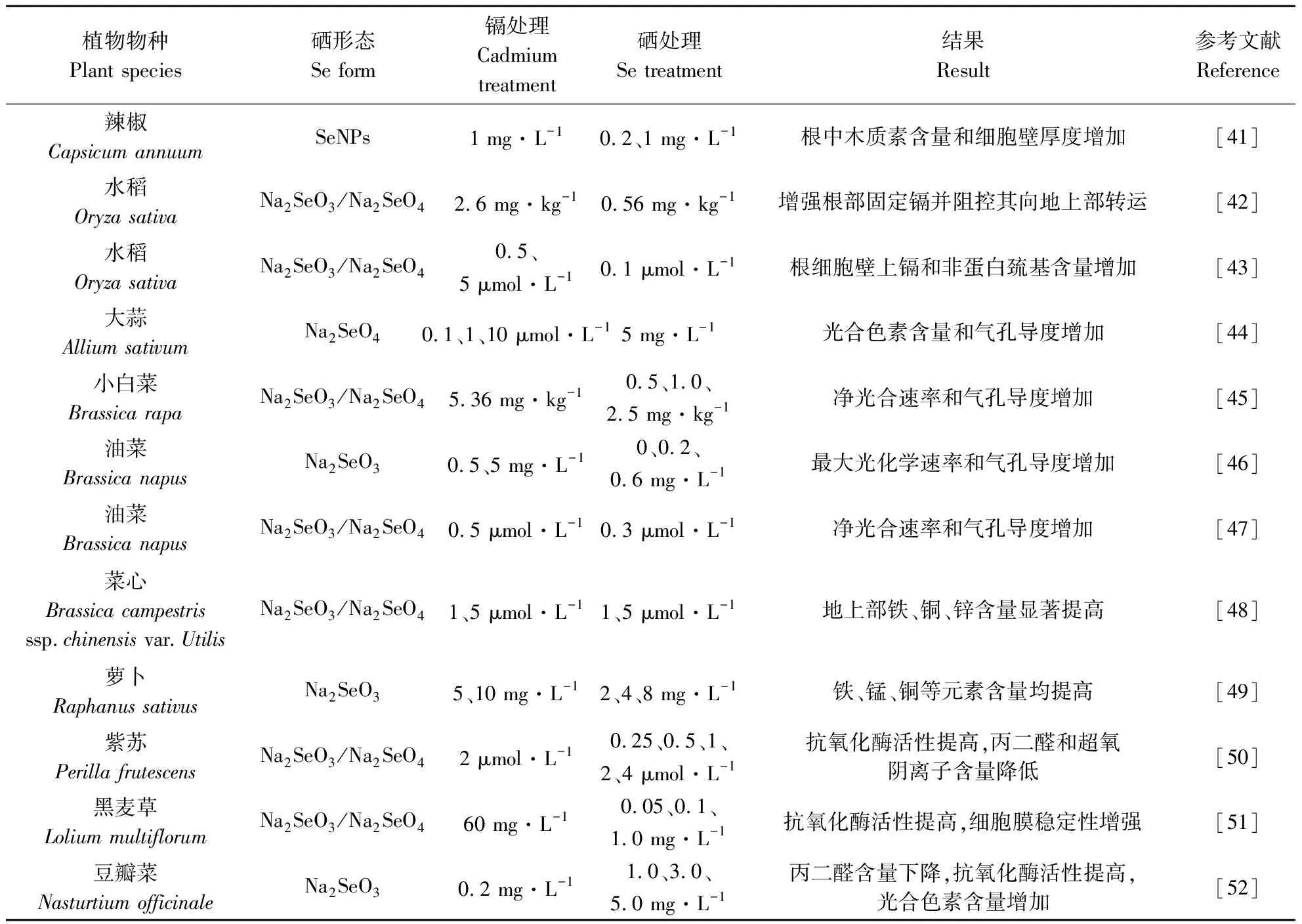
表1 不同浓度和形态硒对植物镉胁迫的缓解作用
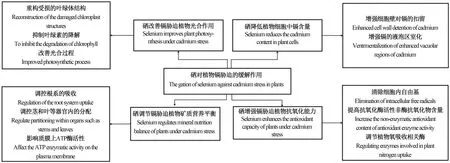
图2 硒对植物镉胁迫缓解作用
2.1 硒改善镉胁迫植物光合作用
土壤中的硒酸盐或亚硒酸盐通过植物根系或叶片进入植物内,并在叶绿体内转化为有机硒(硒代蛋氨酸和硒代半胱氨酸)。因此,植物硒代谢过程与叶绿体密不可分。硒能够帮助植物重构受损的叶绿体结构。FILEK等[56]发现镉胁迫引起油菜叶绿体内膜的降解,而施加2 mmol·L-1硒后,叶绿体超微结构重建,类囊体和基质片层结构重组,叶绿体大小、脂肪酸不饱和度和细胞膜流动性增大。硒能够调控和促进叶绿素的合成代谢,抑制镉胁迫下叶绿素的降解。张庆等[9]在茶树镉胁迫中发现,硒(1.5 mg·L-1)使得叶片中的叶绿素a和b分别增加52.1%和97.0% ,类胡萝卜素含量增加70.8%。同样,在高羊茅、香青兰和冬小麦等[57-59]植物上研究发现,低浓度硒能有效增加叶绿素a、b和类胡萝卜素含量,叶绿素a/叶绿素b的比值上升。硒增加光合色素含量的可能原因:一方面,增加作为非酶抗氧化物的类胡萝卜素等含量,能够显著增强植物的抗氧化能力,提高植物镉胁迫耐受性。另一方面,硒与叶绿素合成过程中关键元素铁密切相关,并能够使得植物对铁保持较高的吸收能力[60]。这种调控机制可能与硒调控叶绿体中铁硫蛋白有关。此外,硒通过改善光合过程,如调控光吸收、光合电子传递、碳同化等。YANG等[61]发现面喷施硒提高了镉胁迫下紫苏叶片的光合系统Ⅱ和电子传递速率,而根施硒提高了镉胁迫下的光合速率、气孔导度、蒸腾速率和气孔限制值。ZHANG等[62]对甘蓝型油菜的研究结果与之类似。卞威乐斯等[63]研究发现适量硒可以提高花生光合系统Ⅱ的最大光合效率和光反应中心的最大活性。硒调节光合作用过程有助于减少细胞内的活性氧积累,但此过程相关研究较少。
2.2 硒调节镉胁迫植物矿质营养的平衡
硒通过调控根系对矿质养分的吸收及在茎和叶等器官内的分配过程,维持植物内部矿质元素的动态平衡[20]。例如,刘帅等[59]研究了硒对镉胁迫下菜心内矿质营养的影响,结果显示随着镉浓度增加,微量元素吸收失衡,而低浓度硒增加了茎内Ca、Cu与叶部Fe、Cu和Zn含量,有效缓解镉的毒害作用,并恢复植株的正常生长。代邹等[64]研究在镉胁迫下,加硒后降低水稻的茎、叶和籽粒Ca、Zn含量,而增加根部的钙和铜含量。此外,仰路希等[65]证实在镉胁迫下,添加硒后的人参菜中Ca含量较单独镉处理增加28.76%,Zn含量增加28.52%。而SETAREH等[60]在萝卜中研究表明,除Zn外,硒均能提高Fe、Mn和Cu等微量元素的含量,而个别施硒使Zn含量降低。刘春梅等[66]对水稻的研究结果与之类似。因此可推断,在植物中硒可能与锌具有协同和拮抗的双重效应。目前,有关硒对镉胁迫下矿质营养代谢过程的十分有限,并因植物种类、植物发育阶段、所研究的器官、硒形态和培养方式(土培或水培)等因素的不同,存在显著的差异。
硒通过影响质膜上ATP酶活性,缓解镉胁迫下矿质营养的失衡。ATP酶广泛分布在植物细胞内,其主要功能是催化ATP的水解并释放能量,完成各种生理代谢过程。其中,跨膜ATP酶控制细胞代谢必需的物质进入,并输出可能阻碍细胞过程的毒素、废物和溶质[67]。K+-ATPase、Ca2+-ATPase、Mg2+-ATPase和H+-ATPase等是促进植物细胞内外离子和溶质转运的关键酶[68],也是阻断有害物质进入细胞内部的有效防御机制。因此,细胞内的跨膜ATP酶是镉胁迫的主要靶点之一。镉胁迫下导致根系细胞表面电位变化,造成H+-ATPase和Ca2+-ATPase活性显著下降,并抑制矿质养分的供应。硒能显著提高跨膜ATP酶的活性,促进矿质元素的运输,并维持胞内离子稳态。LIU等[69]在烟草研究中发现,2.5 mg·kg-1硒能显著促进H+-K+-ATPase、Na+-H+-ATPase和Ca2+-Mg2+-ATPase活性,缓解镉胁迫对细胞内环境的破坏。同样,林莉等[70]对水稻根系的研究结果与之类似。关于硒如何调控矿质元素在根部和地上部分的吸收、分配仍需更多研究,来揭示硒诱导植物内矿质离子代谢平衡的机制。
2.3 硒增强镉胁迫植物抗氧化能力
硒通过清除细胞内自由基、提高抗氧化酶活性和非酶抗氧化物含量,增强植物抗氧化防御系统的能力,降低植物膜脂过氧化和提升氧化胁迫的抗性[71]。例如,YU等[72]研究发现,10 μmol·L-1硒酸盐或亚硒酸盐均增强了抗氧化防御,降低了细胞内过氧化氢和丙二醛的积累,缓解镉胁迫对小白菜的毒害。同样,WU等[73]在油菜中发现,15 mg·L-1亚硒酸盐降低了镉胁迫所产生的超氧阴离子、过氧化氢和丙二醛等含量,抑制了镉的氧化损伤。但是,高浓度亚硒酸盐削弱了小白菜镉胁迫下的缓解效应,亚硒酸盐增加油菜地上部的镉积累。因此,可推断,硒的有益浓度范围在不同植物种类间存在显著差异,并且目前尚未确定。
在水稻、草莓和番茄等[74-76]植物上研究发现,硒直接调控并显著提升镉胁迫下抗氧化酶活性。如SOD、APX、CAT、POD 和GR等,减少细胞内活性氧数量,增强植物抗氧化防御能力,并降低植物镉积累水平。关于硒促进抗氧化酶活性的可能原因:硒调控相关抗氧化酶的辅酶因子,如Fe、Mn、Cu和Zn等元素的吸收和分布,激活相关抗氧化酶高效地清除活性氧[40],从而增强植物对镉胁迫的耐受性。因此,硒对某些必需元素吸收和分配的调控被认为是抗氧化酶、降低活性氧水平和提高植物逆境胁迫耐受性的关键机制。此外,硒可促进非酶抗氧化系统中还原型抗坏血酸的积累,并通过维持谷胱甘肽过氧化物酶途径中α-生育酚和谷胱甘肽的生物合成、减少游离态金属离子等多种反应,消除过量的过氧化氢。同时,硒可能作为信号分子介导GSH-ASA循环,提升细胞内GSH和AsA浓度水平应对镉的氧化胁迫[77]。除活性氧清除系统外,硒可能通过调节植物氮吸收相关酶,减轻镉诱导的氧化胁迫。SHAHID等[78]发现硒提升镉胁迫下氮代谢相关酶活性,如硝酸还原酶、亚硝酸还原酶、谷氨酰胺合成酶和谷氨酸合成酶等,改善硝酸盐同化途径,增强马铃薯抗氧化能力。硒对抗氧化酶和非酶抗氧化物的作用效应不同,且因植物种类、组织部位、处理方式和时间的不同而有明显差别。这可能是由于不同基因型植物或组织、器官对镉或硒的敏感性差异所致。
2.4 硒降低植物细胞中镉含量
硒通过增强细胞壁对镉的扣留,降低植物细胞中镉的含量。细胞壁是镉进入植物的第一道屏障,具有胞间层、初生壁和次生壁三层结构。其中,胞间层主要由果胶组成,初生壁主要由纤维素、半纤维素和果胶组成,而次生壁主要由纤维素和木质素组成。由于果胶、纤维素、半纤维素和木质素上含有大量羟基、羧基、巯基等带负电荷的官能团[79],因此细胞壁成为镉离子沉积的重要场所。例如在玉米和水稻中有超过80%的镉被吸附在细胞壁上[80-81]。近期研究表明,硒能增强植物根系细胞壁中果胶、纤维素、半纤维素和木质素的合成[82],将更多的镉滞留在细胞壁缓解镉胁迫。YANG等[83]发现,硒增加了镉胁迫下油菜中果胶含量和果胶甲酯酶活性,产生更多游离的羧基与镉结合。ZHAO等[84]发现,硒增加了镉胁迫下油菜果胶和半纤维素含量,显著降低了细胞中可溶组分镉的比例。ZHU等[85]对冬小麦的研究结果与之类似。CUI等[86]研究证实,硒通过上调木质素合成基因OsPAL、OsCoMT和Os4CL3表达,提高水稻细胞壁木质素含量,使67.4%~78.8%的镉固定在细胞壁上,减少镉细胞内的转运。。
硒通过增强镉的液泡区室化,降低植物细胞中镉的含量。金属硫蛋白、谷胱甘肽和植物络合素等配体含有大量的巯基位点可与镉离子结合形成可溶态的螯合物或络合物[87]。液泡膜上的跨膜转运蛋白通过主动运输的方式进一步将这些螯合物或络合物转运到液泡内储存起来[88],以减小镉胁迫对细胞正常代谢活动的影响。越来越多的研究发现,硒可以增强镉的液泡区室化储存。ISMAEL等[89]研究表明,硒诱导镉胁迫下油菜植物络合素合酶PCS1基因表达上调,产生更多的植物络合素与镉结合后储存在液泡中。LI等[52]对辣椒的研究中也有类似报道。此外,FENG等[90]和RANA等[91]认为,硒增加镉胁迫下植物络合素合成底物谷胱甘肽含量,形成谷胱甘肽-镉复合物和植物络合素-镉复合物(GSH-Cd和PCs-Cd),随后转运到液泡中储存。
3 展望
外源施硒已成为增强植物镉耐受性的重要策略,因其具有益作用。然而,目前存在以下几个问题。首先,硒的应用范围较窄,因土壤类型、处理时间、施用方法和实验条件等因素而异。因此,必须在特定植物和特定条件下测试硒的最佳范围。此外,需要进一步研究硒对植物进行生物强化的安全性,以避免硒的高浓度施用对植物产生毒害。其次,当前研究主要集中在温室水培或盆栽条件下对硒和镉作用的研究,而田间实践探究还相对不足。因此,需要在田间条件下深入研究硒缓解植物镉胁迫的诱导机制。第三,当前硒与重金属的研究主要围绕硒酸盐和亚硒酸盐两种形态。未来应该广泛研究其他硒形态,如有机硒和纳米硒,以探究它们对各类重金属在植物内的缓解机制。第四,硒与其他代谢产物、植物营养物质、激素和信号分子之间的交互作用需要进一步研究,结合转录组学、代谢组学等手段,以探索提高植物对金属毒害的耐受性和减少富集效应的最佳途径。在这些关键领域的深入研究将有助于进一步阐明硒在增强植物金属耐受性之外的分子机制。同时,这也将为保障食品安全和提高人类健康水平提供重要的科学依据。
猜你喜欢
阅读(科学探秘)(2020年8期)2020-11-06
国际放射医学核医学杂志(2020年2期)2020-05-30
中国果业信息(2019年1期)2019-01-05
中成药(2018年11期)2018-11-24
现代园艺(2017年21期)2018-01-03
生物学教学(2017年9期)2017-08-20
东北电力大学学报(2015年4期)2015-11-13
应用化工(2014年8期)2014-08-08
食品工业科技(2014年6期)2014-05-10
食品工业科技(2014年9期)2014-03-11
