
中国兔源十二指肠贾第虫分子流行病学研究进展
2023-06-28 02:04:16李俊强张苗苗张峻晨付寅秦子扬李晓迎张龙现
河南农业大学学报 2023年3期
李俊强, 张苗苗, 张峻晨, 付寅, 秦子扬, 李晓迎, 张龙现
(1.河南农业大学动物医学院,河南 郑州 450046;2.河南省人兽共患病国际联合实验室,河南 郑州 450046)
十二指肠贾第虫(Giardiaduodenalis)(亦称蓝氏贾第鞭毛虫或肠贾第虫)是引起贾第虫病的主要病原。据估算,全球范围内每年约2.8亿人感染十二指肠贾第虫,其引起的贾第虫病在发展中国家发病率较高[1]。贾第虫病典型的临床症状为腹泻,5岁及以下儿童严重时可能出现死亡[2]。贾第虫主要通过粪口途径进行传播,被感染个体排出的包囊可通过饮水或食物传播,是潜在的传染来源,目前已经报道多起水源性及食源性贾第虫病的暴发[3]。然而,迄今为止尚无有效防控贾第虫病的药物与疫苗[4]。家兔是一种重要的经济动物,同时也是多种人兽共患病原的重要传播来源[5],ROBINSON等[6]和NAGUIB等[7]曾分别报道家兔是导致人隐孢子虫病暴发和感染的主要来源。同时,家兔也可作为宠物进行家庭饲养,其携带的人兽共患病原具有通过食物或饮水进行传播的潜在威胁。国内外报道十二指肠贾第虫在家兔中的感染率为1.9%~72.3%[8-15],基因型鉴定分类为A、B和E这3种集聚体,均具有潜在的人兽共患性[8,14-15],表明家兔在十二指肠贾第虫的人兽共患传播过程中起重要作用。中国是世界上最大的兔肉生产国[16]。本综述对目前中国兔源十二指肠贾第虫的分子流行病学研究进行了总结和分析,以更好了解贾第虫病在中国家兔中的流行病学特点,评估其人兽共患传播风险,以期为中国兔源贾第虫病的防控提供理论指导。
1 十二指肠贾第虫分类
贾第虫(Giardiaspp.)是一种广泛存在于人类和多种动物的肠道寄生性原虫,根据其形态结构和感染宿主范围目前被分为9个种类[17],分别为十二指肠贾第虫(Giardiaduodenalis)、微小贾第虫(Giardiamicroti)、鼠贾第虫(Giardiamuris)、敏捷贾第虫(Giardiaagilis)、苍鹭贾第虫(Giardiaardeae)、鹦鹉贾第虫(Giardiapsittaci)、瓦氏贾第虫(Giardiavarani)、袋狸贾第虫(Giardiaperamelis)和仓鼠贾第虫(Giardiacricetidarum)。新的贾第虫种还在不断被发现与鉴定,其中袋狸贾第虫和仓鼠贾第虫分别于2016年[18]和2018年[19]被命名。
十二指肠贾第虫宿主范围最为广泛,可感染包括人在内的多种哺乳动物,具有重要的人兽共患性[20]。十二指肠贾第虫在全球范围内流行,对人类、宠物及家畜健康造成严重危害,引起较为重大的经济损失[21]。十二指肠贾第虫是一个复杂的综合体[22],常用于十二指肠贾第虫基因型分型的位点包括小亚基核糖体(small subunit ribosomal RNA,SSUrRNA)、磷酸丙糖异构酶(triosephosphate isomerase,tpi)、谷氨酸脱氢酶(glutamate dehydrogenase,gdh)和β-贾第素(β-giardin,bg)。通过比较基因片段的特异性和序列的差异性,十二指肠贾第虫被分为8个集聚体(A~H),每个集聚体形态学差异不显著,但具有一定的宿主特异性[20-21]。十二指肠贾第虫集聚体A和集聚体B感染人类和多种哺乳动物,具有显著人兽共患潜力[2];集聚体C和D主要感染犬科动物;集聚体E主要感染猪和反刍动物;集聚体F多感染猫科动物;集聚体G仅感染啮齿类动物;集聚体H仅感染海洋动物[20]。十二指肠贾第虫不同集聚体之间感染宿主范围存在显著差异,因此将不同的集聚体分别命名为不同的贾第虫种[20-21]。
根据宿主范围,十二指肠贾第虫集聚体A和B具有广泛的宿主范围,可在人类与动物间进行传播,严重危害公共卫生安全[2]。到目前为止,除了集聚体A和B之外,集聚体C、D、E和F也均有感染人的零星报道[23-29],其中集聚体E感染人的报道稍多[23,25-27,29]。基于十二指肠贾第虫多位点基因分型(multi locus genotyping,MLG)技术,集聚体A又可以分为3个亚型,分别为AⅠ、AⅡ、AⅢ,各亚型之间具有寄生宿主偏好性[21,30]。亚集聚体AⅠ主要感染家畜;AⅡ主要感染人;AⅢ主要感染野生哺乳动物[21]。集聚体B之前被分为2个亚型(BⅢ和BⅣ)[21-22],然而在系统发育进化分析中并没有获得足够支持,且有其他亚型存在[9,31-32]。
2 兔源十二指肠贾第虫概述
针对兔源十二指肠贾第虫的分子流行病学研究主要是利用巢式PCR扩增和序列分析技术[20],可以鉴别贾第虫不同物种及同物种的不同基因型或亚型,对于贾第虫基因型(集聚体)分析常用的靶点基因包括SSUrRNA、bg、gdh和tpi等[20,33]。其中,SSUrRNA基因最为保守,主要用于检测其感染情况;bg、tpi和gdh基因主要用于集聚体或亚型鉴定[13]。PCR检测具有特异性强和敏感性高的特点,但不同集聚体混合感染同一宿主或集聚体遗传差异较小时,难以依靠单一分子位点对十二指肠贾第虫分离株进行准确分型鉴定与区分混合感染情况,因此需要进行多位点基因分型技术进行基因型的鉴定。
目前,在世界范围内,关于兔源十二指肠贾第虫的流行病学调查研究较少。除中国外,只有尼日利亚、厄瓜多尔、澳大利亚和欧洲的部分国家报道了兔源十二指肠贾第虫的感染情况。其中,尼日利亚的兔源十二指肠贾第虫的感染率最高,可达72.3%[8];其次是厄瓜多尔为20.0%[34];欧洲多个国家的平均感染率为7.6%[35];俄罗斯和澳大利亚的感染率分别为1.8%[36]和1.0%[37]。这表明兔源十二指肠贾第虫在世界范围内严重危害动物健康和威胁公共卫生安全。
3 兔源十二指肠贾第虫流行病学分析
3.1 兔源十二指肠贾第虫分子流行情况
截至2022年6月,中国共有9个省、自治区、直辖市报道了兔源十二指肠贾第虫的分子感染流行情况(表1),感染率为1.5%~18.7%,总体平均感染率为9.5%(412/4 324)。不同地区兔源十二指肠贾第虫的平均感染情况存在差异,其中,吉林地区感染率最高,为18.4%(81/438);黑龙江、山东、甘肃、河南、上海、陕西、新疆地区感染率分别为16.1%(95/590)、11.2%(69/616)、8.6%(51/592)、8.4%(80/955)、6.5%(9/139)、3.5%(19/537)和1.9%(6/321);辽宁地区感染率最低,为1.5%(2/136)。不同地区兔源十二指肠贾第虫感染率不同,可能是采样的动物品种、饲养条件和免疫状态等条件不同所致,但是总体感染率较高,应该引起足够的重视。
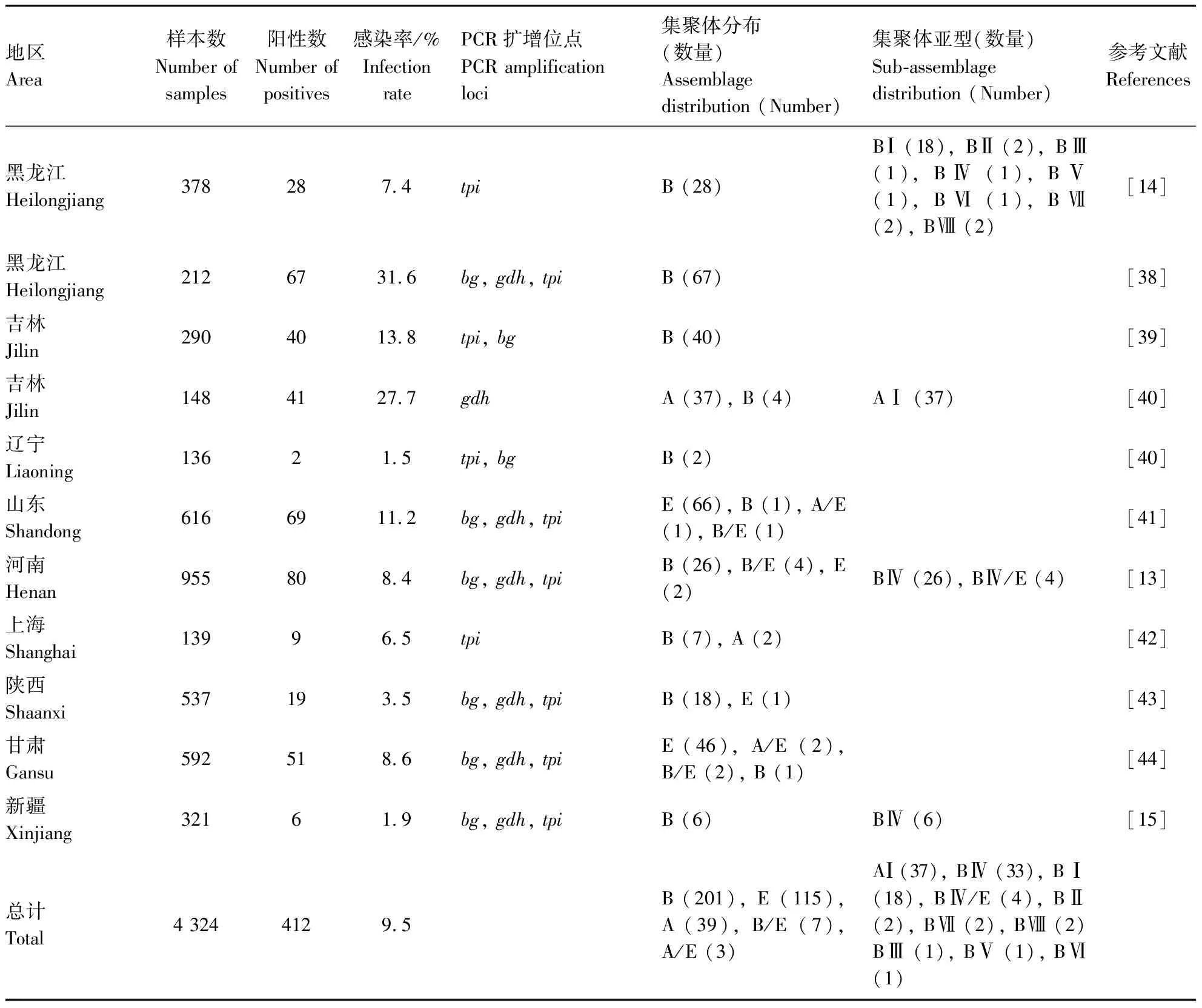
表1 中国兔源十二指肠贾第虫流行和集聚体分布情况
3.2 兔源十二指肠贾第虫感染风险因素分析
家兔是一种重要的经济动物。据统计不同的年龄、品种、经济用途和饲养地点的家兔中十二指肠贾第虫的感染率各异。对于家兔不同的年龄组,虽然十二指肠贾第虫感染率差异不显著(χ2=0.42,P>0.05),然而年龄≥6月龄家兔十二指肠贾第虫的感染率(7.8%,132/1 695)要略高于年龄<6月龄的感染率(7.2%,149/2 077)[13,15,39,41-45]。
家兔品种繁多,包括獭兔、伊拉兔、长毛兔、蒙古兔、新西兰白兔等。不同品种间兔源十二指肠贾第虫的感染率差异显著(χ2=127.23,P<0.001),以蒙古兔的感染率较高,为23.1%(51/221)[41];伊普吕兔的感染率次之,感染率为13.1%(13/99)[44];新西兰白兔、伊拉兔和獭兔的感染率分别为7.2%(15/207)[41]、6.2%(52/836)[43-44]和5.1%(43/839)[13,15,43-45];长毛兔的感染率较低,为1.1%(4/377)[41,45];伊乐高兔和比利时兔未检测到十二指肠贾第虫的感染[44]。
家兔根据不同经济用途可以分为肉用兔、皮用兔、毛用兔、宠物兔、试验兔和皮肉兼用兔。以不同经济用途进行分类统计十二指肠贾第虫感染时发现,不同用途之间兔源十二指肠贾第虫感染差异显著(χ2=109.07,P<0.001)。其中,皮肉兼用兔的感染率最高,为23.1%(51/221)[41];肉用兔和试验兔的感染率次之,分别为7.9%(125/1 588)[41.43-44]和7.2%(37/517)[14,42];皮用兔感染率为5.1%(43/839)[13,39,43-44];毛用兔的感染率最低,为1.1%(4/377)[13,42,45]。
家兔的饲养地点一般分为室内饲养和室外饲养。当比较不同饲养地点对家兔感染十二指肠贾第虫的影响时,共统计1 042份家兔粪便样品,户外饲养的家兔十二指肠贾第虫的感染率为19.2%(76/396),室内的感染率为5.4%(35/646)[39,41],不同饲养地点家兔感染十二指肠贾第虫差异显著(χ2=47.51,P<0.01)。以上不同类型(年龄、品种、用途、饲养地点)所导致的家兔十二指肠贾第虫感染率的差异,可能是不同的宿主抵抗能力、饲养环境、营养状态和生态因素等共同因素导致[39]。
3.3 兔源十二指肠贾第虫基因型分布
目前,在中国家兔中共检测到3种十二指肠贾第虫集聚体,分别为集聚体A、B和E[8,13,37,41]。山东、甘肃和河南地区的家兔中还检测到了十二指肠贾第虫集聚体B/E和集聚体A/E的混合感染。十二指肠贾第虫集聚体A、B和E均具有潜在的人兽共患性,集聚体B在中国为优势集聚体,占总检测数量的55.1%(201/365);其次为集聚体E,所占比例31.6%(115/365);集聚体A的感染相对较低,所占比例10.7%(39/365);混合感染所占比例2.7%(10/365)。
不同地区兔源十二指肠贾第虫的集聚体分布存在显著差异,在现有的研究报道中,均检测到集聚体B的感染,而且大部分地区报道是以集聚体B为优势亚型(表1)。然而,山东[41]和甘肃[44]地区以集聚体E为优势基因型,吉林地区以集聚体A为优势基因型[40]。这种集聚体基因型分布的差异可能与家兔饲养环境、种群密度、营养状况和生态环境等因素有关。
对于兔源十二指肠贾第虫进行了集聚体亚型分析发现,集聚体A亚型AⅠ在吉林地区的家兔中被鉴定[40];十二指肠贾第虫集聚体B亚型BⅣ广泛分布于河南[13]和新疆[15]。在黑龙江地区,兔源十二指肠贾第虫集聚体B亚型具有多样性,可以分为不同的亚型[14,38]。这种不同亚型的鉴定结果,主要是集聚体B相对于其他集聚体具有较强的遗传多样性所致。
3.4 兔源十二指肠贾第虫系统发育分析
为探讨兔源十二指肠贾第虫集与其他宿主源(人、牛、羊、猪、犬、猫、鼠等)分离株的系统发育进化关系,并更直观地阐明,本综述利用已有数据库数据制作系统发育进化树。通过Clustal X 2.0软件将GenBank数据库中获得的十二指肠贾第虫不同集聚体数据[46],基于邻接法(Neighbor-Joining)使用MEGA X软件,选择Kimura 2-parameter模型,使用bootstrap检验,进行1 000次重复计算,分别构建出基于tpi(图1)、gdh(图2)、bg(图3)基因序列的系统发育进化树。基于十二指肠贾第虫不同基因的系统发育进化树分析表明,兔源十二指肠贾第虫集聚体A、B和E分离株与其他宿主源的集聚体分离株分别聚集在同一进化分枝,并表现良好的自举值。这与KUMAR等[47]研究结果一致。
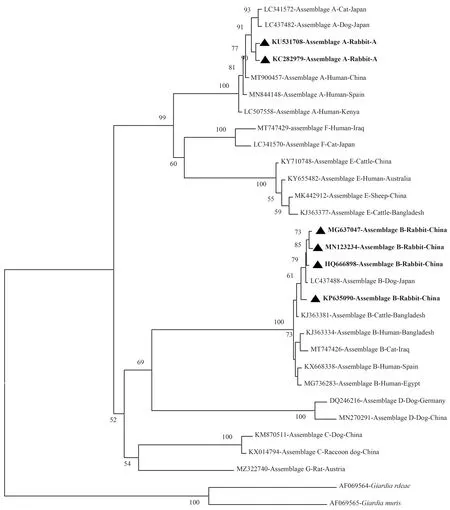
三角形代表兔源分离株。下同。
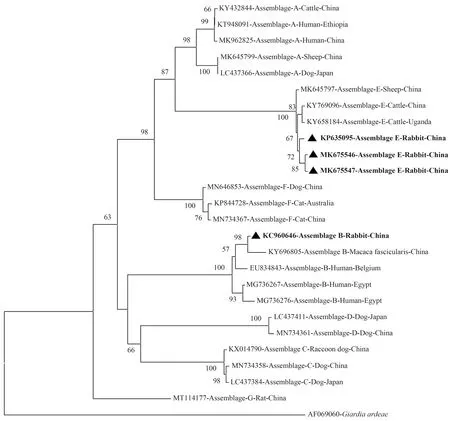
图2 兔源十二指肠贾第虫不同集聚体gdh基因位点系统发育进化树
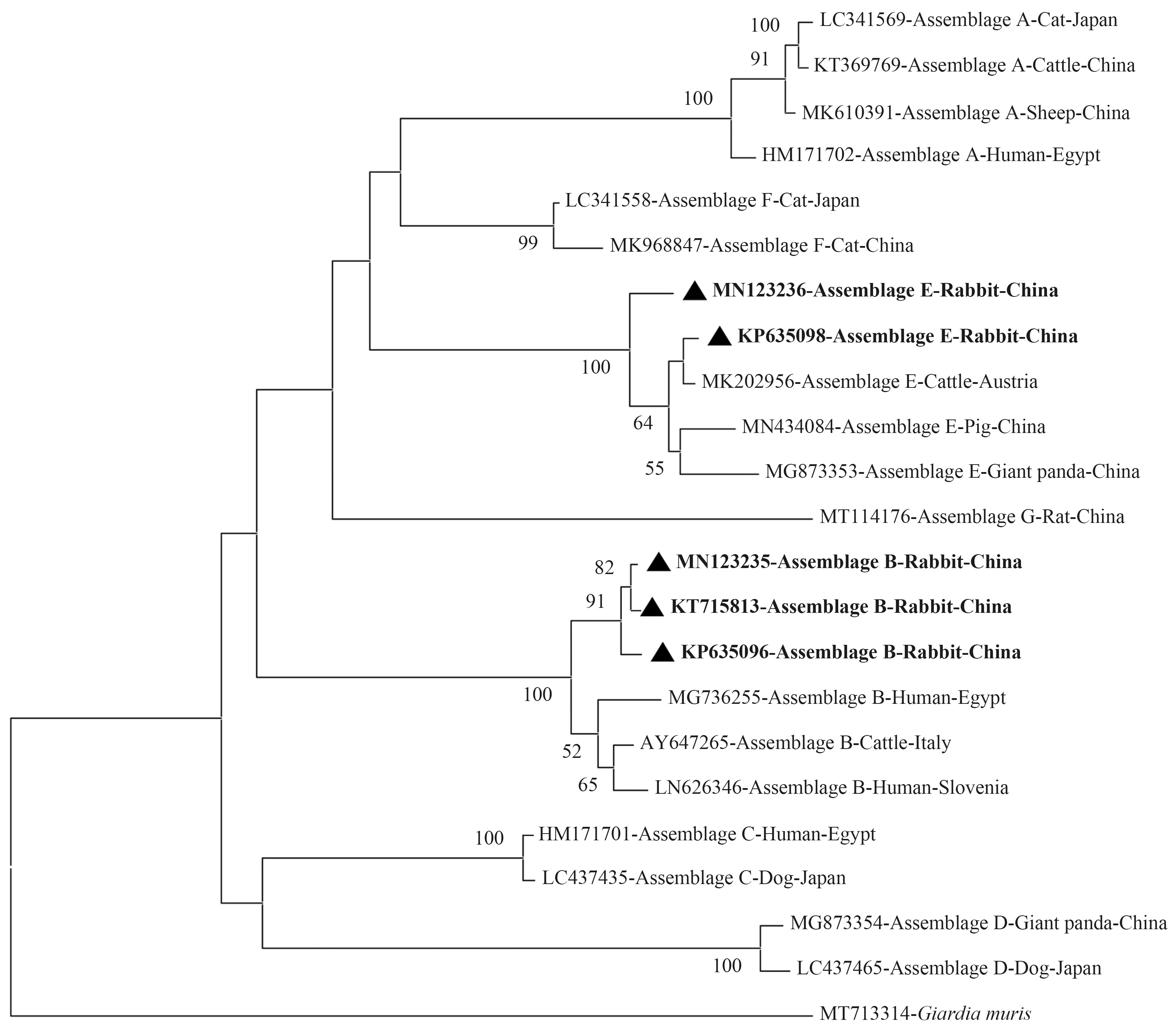
图3 兔源十二指肠贾第虫不同集聚体bg基因位点系统发育进化树
3.5 兔源十二指肠贾第虫基因组特征分析
第二代和第三代全基因组测序技术的提高和完善为十二指肠贾第虫基因组的全基因组测序提供了技术与支持。目前,已经获得人源和部分动物源十二指肠贾第虫集聚体A、B、C、D和E分离株的全基因组序列[48-50],基因组大小为10.7~12.8 Mb,特别是对于集聚体A分离株的测序和组装基因组序列质量还在持续不断优化[51]。然而,对于兔源十二指肠贾第虫的基因组的研究还未见报道。因此,关于兔源十二指肠贾第虫基因组特征的研究,特别是家兔优势集聚体B的基因组特征,以及与其他动物源集聚体之间的比较基因组学将是今后的研究方向之一。
4 存在问题与展望
对十二指肠贾第虫的研究对于防控工作具有重大的公共卫生意义[52]。中国已经有9个省、自治区、直辖市报道了兔源十二指肠贾第虫的分子流行情况,其总体平均感染率为9.5%。在家兔中共检测到3种十二指肠贾第虫集聚体,分别为集聚体A、B和E,均具有潜在的人兽共患性,集聚体B在中国家兔中为优势基因型。家兔是一种重要的经济动物,中国是世界上最大的兔肉生产国,同时家兔也是人兽共患病原的重要传播来源,其携带的人兽共患病原体具有通过食物或饮水进行共患传播的可能性。因此,关于兔源十二指肠贾第虫的感染和流行情况的调查研究应该引起足够的重视。兔源十二指肠贾第虫的感染是以集聚体B为主,表现出明显的宿主适应性特征。此外,十二指肠贾第虫集聚体B也是人类、非人灵长类动物、啮齿动物和马属动物等宿主感染的优势基因型[1],同样对这些宿主表现出宿主适应性特征。因此,这些宿主适应性的分子机制值得深入研究,也是今后关于十二指肠贾第虫的重要研究方向之一。同时,对于十二指肠贾第虫不同集聚体基因组特征,以及比较基因组学将有助于进一步解析贾第虫在分类学、遗传进化方面等的研究,并最终对贾第虫病传播动力学和防控措施的制定起到积极作用。
另一方面,十二指肠贾第虫宿主范围广泛,可感染多种类型的野生动物、家养动物和啮齿动物等。家养反刍动物是十二指肠贾第虫重要的保虫宿主,啮齿动物在十二指肠贾第虫的跨物种传播过程中可能起到重要作用[53]。然而,这些均需要更多的调查数据进行进一步证实和验证。十二指肠贾第虫鉴定的8个集聚体(A~H)中,其宿主范围和遗传特征均存在显著的差异性。鉴于此,有多位学者提议将十二指肠贾第虫不同的集聚体及亚型分别命名为不同的贾第虫种类,以充分体现其宿主范围和遗传特征[54]。该提议还需要更多的响应和认可,但这表明对于十二指肠贾第虫分类地位和遗传进化的研究仍是今后重要的研究方向。随着全基因组测序技术的发展和提高,基于基因组学和比较基因组学研究十二指肠贾第虫不同集聚体之间的基因组序列特征,将推动贾第虫在分类和遗传进化等方面的研究进展,并最终对贾第虫病传播动力学和防控措施的制定起到积极作用。
猜你喜欢
动物医学进展(2024年3期)2024-03-18 07:09:54
今日农业(2021年17期)2021-11-26 23:38:44
兽医导刊(2020年8期)2020-12-28 15:09:07
科学(2020年3期)2020-11-26 08:18:20
中国人兽共患病学报(2019年11期)2019-01-10 20:04:42
作文·初中版(2017年4期)2017-04-18 11:20:14
天津农学院学报(2016年2期)2016-12-01 05:40:09
兽医导刊(2015年9期)2016-01-04 12:00:06
兽医导刊(2015年7期)2016-01-04 11:59:58
肝胆胰外科杂志(2015年1期)2015-02-27 11:11:34
