
种植密度对菊芋块茎产量、植株光能截获及养分转运特性的影响
2023-06-28 02:04:18张铎丰晨晨樊燕鸽苗玉红王宜伦李岚涛
河南农业大学学报 2023年3期
张铎, 丰晨晨, 樊燕鸽, 苗玉红, 王宜伦, 李岚涛
(1.河南农业大学资源与环境学院,河南 郑州 450046; 2.河南农业大学林学院,河南 郑州 450046; 3.河南省科学院化学研究所有限公司,河南 郑州 450014)
菊芋(Helianthustuberosus),又名鬼子姜,属多年生草本植物,原产于北美,后经欧洲传入中国,由于其具有耐寒、耐盐碱、耐贫瘠等优良特性,目前在中国各地均有种植[1]。菊芋块茎中富含菊粉(菊糖),菊糖是一种天然果聚糖混合物,其所含β(1→2)糖苷键不易被人体消化吸收,可有效降低人体吸收能量,起到膳食纤维作用;菊粉可在乳品、饮料、焙烤制品、保健食品中广泛使用,已被世界上四十多个国家列为食品添加剂[2-4]。菊芋叶片中所特有的绿原酸被称为“第七大类营养素”,广泛应用于保健行业,具有改善胰岛素敏感性,调节糖、脂代谢等功能[5]。菊芋茎秆粗壮,叶片肥厚,生长旺盛阶段可直接刈割作家畜青饲料,茎秆收获后也可加工成颗粒饲料储藏使用[6]。菊芋因其发达的枝干和根系可有效防风固沙,有“地上一把伞,地下一张网”之称[7]。种植密度是影响作物产量的重要因素之一,对作物群体结构、生理生态、光能截获、养分吸收和产量等均有显著影响。大量研究表明,种植密度是影响作物产量的决定性因素,适宜的密度可充分利用光能,从而增加作物单株产量,提高群体产量[8]。门果桃等[9]通过三因素随机区组试验研究了密度、基肥和追肥等因子对菊芋产量的影响,发现密度对菊芋产量影响最大。张宇等[10]采用科尔沁菊芋开展的密度效应田间试验表明,在行距60 cm、株距30 cm的条件下,菊芋产量高达4.5×104kg·hm-2。刘冰等[11]研究指出,在行株距为30~40 cm时,菊芋产量最高且光合能力较强。此外,合理的冠层结构可显著提高作物光合有效辐射率,而作物生长后期基部叶片郁积遮阴,减少了冠层光合有效辐射截获量,不利于光合产物的积累[12-13]。杨晓璐等[14]指出,不同的株距和行距对马铃薯光分布、光合特性和叶绿素荧光特性均有明显影响。柏延文等[15]认为,高密度条件下玉米中间层可截获更多的光能,且随种植密度增加,玉米吐丝后干物质积累量、干物质转运量及其对籽粒贡献率均呈先升高后降低趋势。凌一波等[16]研究表明,花前干物质转运量和转移率随密度增大而增加,花后干物质积累量和转移量则随密度增加而降低。菊芋作为重要的能源和经济作物,合理密植对其产量形成、品质构建和植株生长发育均具有重要影响。然而,目前基于田间试验系统探究菊芋种植密度对其块茎产量、光能截获及养分吸收转运的报道则相对匮乏。基于此,本研究利用2 a菊芋密度效应田间试验,探究菊芋密度与产量间关系,明确种植密度对菊芋光能截获及养分吸收转运规律影响,确定适宜种植密度,为当地菊芋优化栽培种植和高产稳产提供科学依据。
1 材料与方法
1.1 供试材料
2019—2020年分别于河南省新乡市河南农业大学现代科教园区(35.11°N,113.95°E)和南阳市方城县汉山村(33.28°N,112.87°E)布置菊芋密度效应田间试验。供试土壤新乡为沙质潮土,南阳为壤质黄褐土,0~20 cm耕层土壤理化性状如表1所示。供试菊芋品种均为“南芋1号”。
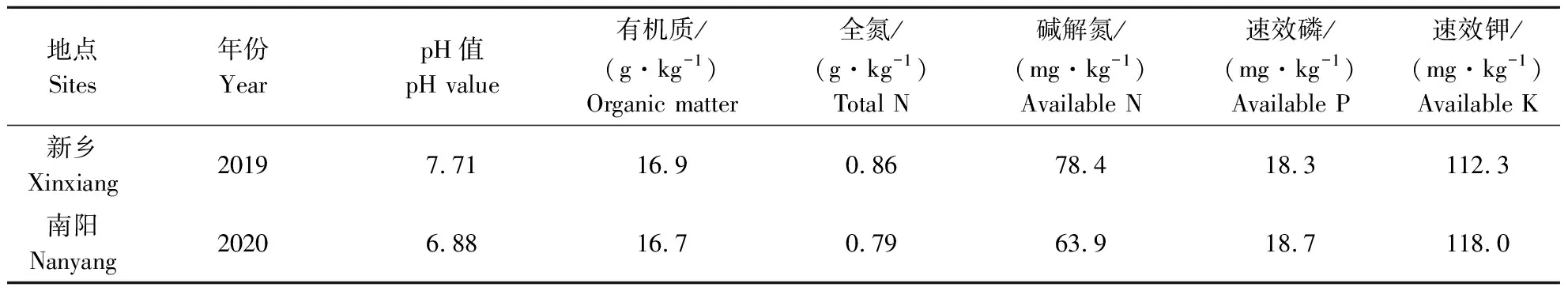
表1 试验田土壤基础养分状况
1.2 研究方法
2 a田间密度试验均设置5个种植密度,分别为D1(1.80×104株·hm-2)、D2(2.25×104株·hm-2)、D3(2.70×104株·hm-2)、D4(3.15×104株·hm-2)和D5(3.60×104株·hm-2)。新乡点于2019-03-18日播种,2019-11-04收获;南阳点于2020-03-21播种,2020-11-12收获。所有处理小区面积均为20 m2(长×宽=5.0 m×4.0 m),3次重复,随机区组排列。氮肥、磷肥和钾肥用量分别为N 180 kg·hm-2、P2O5120 kg·hm-2和K2O 135 kg·hm-2。氮、磷、钾肥品种分别为ESN树脂包膜尿素(N质量分数44%),过磷酸钙(P2O5质量分数12%)和硫酸钾(K2O质量分数54%),所有肥料均作基肥一次性施入。其他栽培管理措施均同当地农户保持一致。
1.3 项目测定与方法
1.3.1 菊芋块茎产量的测定 于菊芋成熟期,选取各小区测产区(10 m2)菊芋全部单刨,单收,称取鲜质量,根据已采植株样及占用面积计算单位面积的块茎产量/(kg·hm-2)。
1.3.2 菊芋生理生化指标的测定 叶片SPAD值的测定分别于菊芋营养生长中期(MVP)、营养生长末期(LVP)和开花期(FP),各小区选取10株长势一致的植株,采用日本产Minolta SPAD-502 型叶绿素计,沿主茎自上而下测其植株上、中和下层完全展开叶SPAD值(根据植株株高,将植株均分为3层),将同一小区内测试结果求取平均值。
冠层PAR的测定 采用AccuPAR冠层分析仪(美国Decagon公司生产)进行测定光合有效辐射(photosynthetically active radiation,PAR)。每个小区选取中间2个行间,与作物行向垂直,测定3个不同位点,分2层测定:冠层顶部和冠层底部入射的PAR、反射的PAR,取平均值作为该观测点的PAR,单位为μmol·m-2·s-1,计算冠层吸收性光合有效辐射量(APAR)和吸收系数(FPAR)[17]。
植株养分的测定 从10株菊芋中选取4株,烘干、称质量,计算地上部植株生物量/(kg·hm-2);参照《土壤农化分析》,分别采用凯氏定氮法、钼锑抗比色法、火焰光度法测定植株地上部氮、磷、钾含量(%),并计算植株氮、磷素和钾素积累量/(kg·hm-2)[18]。
1.4 数据处理与分析
参照王丹丹[19]的方法计算菊芋养分相关指标,采用Microsoft Excel 2019进行基础数据输入与前期处理;SPSS 20.0软件进行试验点与密度处理间方差分析和显著性检验(LSD法),显著性水平设定为P<0.05;Origin 2018软件作图。
2 结果与分析
2.1 种植密度对菊芋块茎产量的影响
种植密度显著影响成熟期菊芋块茎产量(图1)。随种植密度增加,菊芋块茎产量均呈先升高后趋于稳定趋势变化。2019年两者间关系可用“线性+平台”方程拟合,2020年则可用“一元二次方程”拟合,决定系数(R2)分别为0.903和0.991,效果较为理想。根据上述密度效应函数,计算可得2019和2020年试验区菊芋适宜种植密度分别为3.15×104和3.24×104株·hm-2,对应产量分别为3.89×104和4.58×104kg·hm-2。交互作用方差分析表明,试验地点(S)和处理(T)对作物产量影响均达到极显著水平,试验地点和处理交互作用(S×T)对作物产量无显著影响。
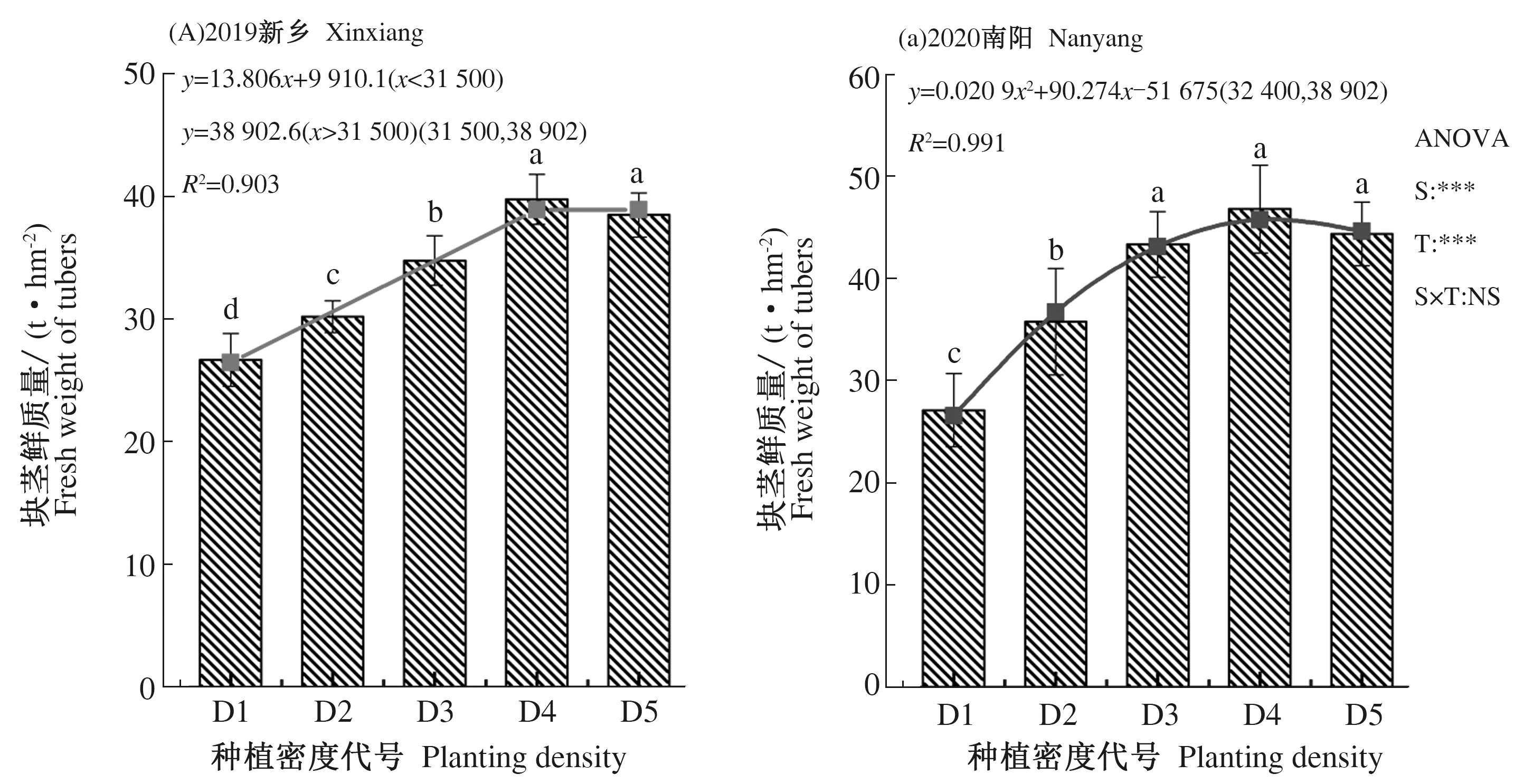
图中不同小写字母表示不同处理间在P<0.05水平差异显著,NS、**和***分别表示试验地点、处理及其交互作用不显著、在0.05、0.01和0.001水平上显著。下同。
2.2 种植密度对菊芋不同叶层SPAD值的影响
为深入探究种植密度对菊芋叶片叶绿素含量的影响,采用叶绿素计SPAD仪测试菊芋植株不同叶层叶片SPAD值(图2)。叶层间,2 a不同生育期菊芋叶片SPAD值均随叶层下移呈先升高后降低趋势变化,即中层>上层>下层。密度间,上部叶随种植密度增加,菊芋叶片SPAD值无显著差异;中层和下层差异性则显著升高,且叶层越深,密度间差异性越高。综合各部位和生育时期分析,试验地点(S)和处理(T)对叶片SPAD值影响分别达95%和99%显著性水平,交互作用(S×T)对叶片SPAD值则无明显影响。
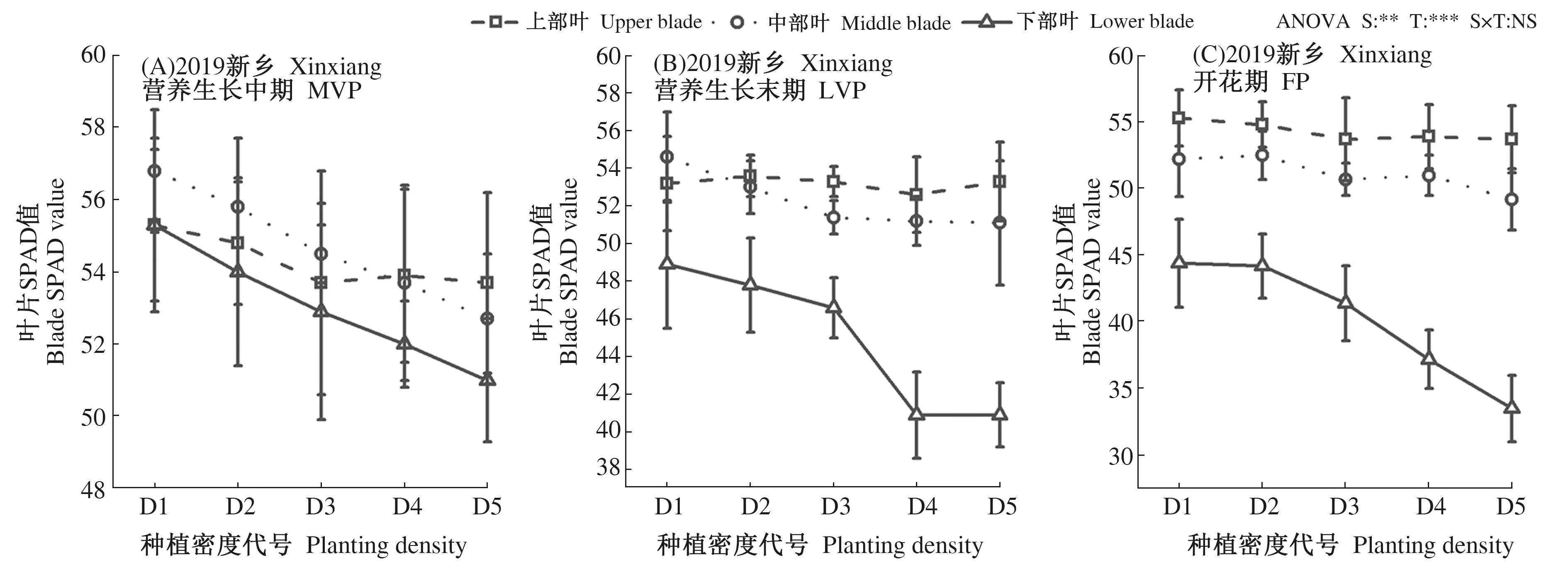
图2 种植密度对菊芋叶片SPAD值的影响
2.3 种植密度对菊芋光合有效辐射的影响
菊芋植株冠层各生育时期APAR和FPAR受种植密度影响显著(表2)。随着种植密度增加,2019年MVP和LVP其APAR呈先上升后下降趋势,2019年FP和2020年所有生育时期则均呈逐渐上升趋势。FPAR除2019年MVP外,其他所有生育期均呈逐渐上升趋势。各生育时期综合分析,随着种植密度增加,2 a APAR和FPAR均呈逐渐上升趋势,在种植密度为D5时达到最大。其中,2019年APAR和FPAR最高(D5)较最低(D1)分别增长了41.3%和42.1%,2020年则分别为69.3%和82.9%。由表2可知,试验地点及处理对菊芋冠层APAR和FPAR均有显著影响,交互作用对菊芋APAR无显著影响,对FPAR则影响显著。
2.4 种植密度对菊芋养分积累和转运的影响
2.4.1 种植密度对菊芋氮素积累和转运影响 由表3知,随种植密度增大,各生育时期菊芋氮素积累量均呈先升高后趋于稳定趋势变化。2019年新乡点,与D1相比,综合生育时期效应,种植密度分别为D2、D3、D4和D5时,氮素积累量分别平均提高23.8%、37.1%、61.7%和54.7%;2020年南阳点增幅则分别平均为8.8%、25.2%、54.5%和45.6%。此外,适宜提高种植密度可有效增加菊芋花前转运量和花后转运量,花前转运率则显著下降。
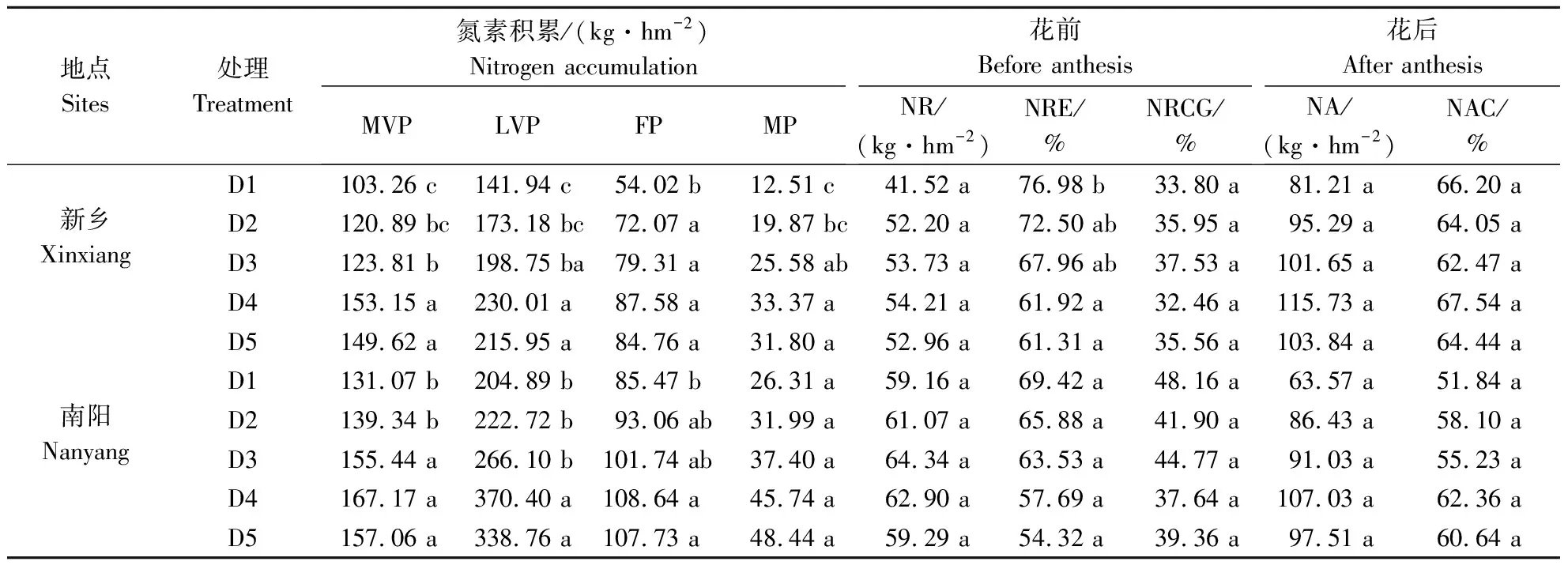
表3 种植密度对菊芋氮素积累和转运率的影响
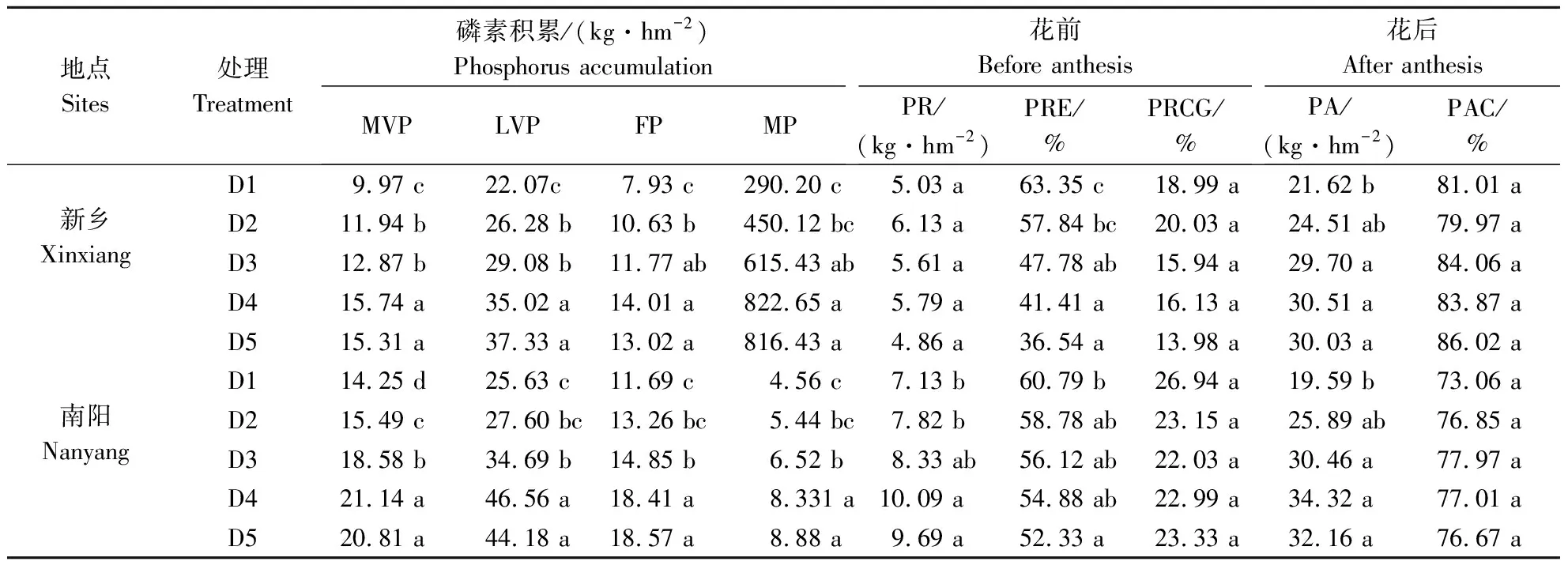
表4 种植密度对菊芋磷素积累和转运率的影响
2.4.2 种植密度对菊芋磷素积累和转运影响 适宜的种植密度可显著提高菊芋各生育期磷素积累量、花前磷素转运率和花后磷素积累量(表5)。2019年和2020年各时期磷素积累量均随种植密度增加呈抛物线趋势变化,至D4时达最高。综合2 a各生育时期数据分析,D4较D1、D2、D3和D5时分别提高了69.1%、45.4%、24.5%和0.7%。2019年,花前养分转运量和花后养分转运量最高(D4)较最低(D1)分别提高了15.1%和41.1%,2020年则分别提高了41.5%和75.2%。
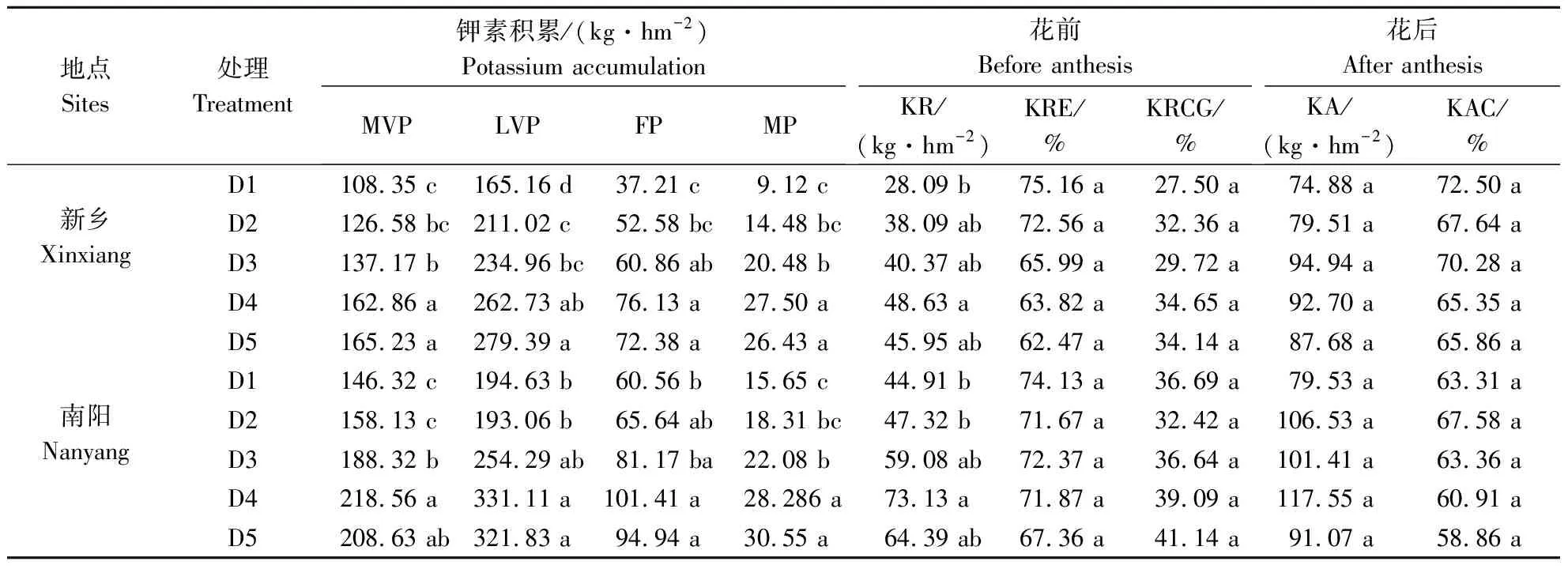
表5 种植密度对菊芋钾素积累和转运的影响
2.4.3 种植密度对菊芋钾素积累和转运影响 不同种植密度可显著影响菊芋各生育时期钾素积累量和花前钾素转运量(表5)。随种植密度增加,除2019年在MVP和LVP时钾素积累量在种植密度为D5时最高,其他时期均在种植密度为D4时最高。综合各生育时期效应分析,2019年当种植密度为D2、D3、D4和D5时较D1钾素积累量分别提高26.5%、41.8%、65.5%和69.9%,2020年则分别增加4.3%、30.8%、62.9%和57.2%。此外,种植密度可显著影响花前钾素转运量,随种植密度增加,花前钾素转运量呈现出先升高后降低的趋势。
3 结论与讨论
研究表明,种植密度可显著影响作物养分吸收和光温资源捕获效率,密度压力增大会导致产量出现显著差异,合理栽培密度可有效提高作物对光能与空间的利用率,促进作物单株生长发育,在一定程度上提高作物产量[20]。种植密度作为制约群体与个体生长的因素之一,通过影响植物的营养生长和生殖生长,进而对农作物产量造成影响[21]。李奋灿[22]研究表明,菊芋种植密度为2.45×104株·hm-2时会获得最大的地上生物量和块茎产量。本试验结果显示,2019年菊芋产量与种植密度关系符合“线性+平台”趋势变化,2020年则符合“一元二次方程”趋势变化,虽变化趋势不同,但2 a试验结果均表明,当菊芋种植密度为D4时可获得最大块茎产量,当种植密度过小或过大时,则不利于产量提升。光能利用特性是表征植物固定太阳能效率的重要指标[23]。植物群体结构是影响光能冠层分布的的重要原因,作物高产高效与合理群体结构,良好的冠层光辐射分布及光能高效利用密不可分[24-25],合理密植是作物利用光能资源,发挥群体优势的基础和保障,其对作物光合利用特性具有显著的调控作用[26-27]。叶片SPAD值与植物光合作用和养分吸收效率密切相关。SPAD值较高时叶片可充分利用光能进行有效的光合作用。相反,SPAD值含量低的叶片光合功能差,干物质积累量少,叶片也存在过早衰老的现象[28]。李岚涛等[29]指出,夏玉米不同叶层叶片SPAD值于植株间分布呈明显的“钟型”变化。本研究表明,随种植密度增加,菊芋上、中、下部叶片SPAD值均呈下降趋势,可能是由于菊芋地上部较大,随着种植密度增加从而出现了光照资源竞争。植株光合有效辐射(PAR)是太阳辐射中对植物光合作用有效的光谱成分,波长范围380~710 nm[30],吸收性光合有效辐射(APAR)则是植物冠层吸收的参与光合生物量累积的PAR部分;FPAR是植被冠层吸收利用光能的重要参数,是定量描述植被光合作用、生产潜力的重要指标,同时也是生态系统的功能模型、作物生长模型、净初级生产力模型、气候模型等的重要参量[31-32]。本研究中,随种植密度增加,菊芋APAR和FPAR均呈上升趋势,主要原因是随种植密度增加,菊芋叶片之间互相遮挡,光能截获能力增加。栽培密度可以显著影响菊芋不同生育时期地上部植株对养分的吸收积累效应。本试验中,2 a菊芋对氮、磷、钾营养元素的吸收、转运和积累随生长发育进程推进呈先增加后减少趋势。处理间,在种植密度为3.15×104株·hm-2条件下,菊芋地上部氮、磷、钾养分积累量达到最高,种植密度对菊芋生育时期内磷素营养转运影响较大,对氮、钾元素转运有一定的影响,但未达到显著水平。
本研究结果表明,种植密度可显著影响菊芋块茎产量、生育时期光能截获及养分吸收积累与转运特性。随着种植密度的增加,菊芋叶片中下层叶片SPAD值逐渐减小,吸收性光合有效辐射量(APAR)和吸收系数(FPAR)则逐渐增加,各生育期养分积累量、开花期养分转运量,花后养分转运量和养分转运率总体均呈逐渐升高趋势。通过“线性+平台”和“一元二次方程”拟合,本试验条件下,菊芋适宜种植密度为3.15×104~3.30×104株·hm-2,在此密度范围内,菊芋产量达到最高,群体间光能截获及养分吸收利用效能最佳。
猜你喜欢
中国农业信息(2022年1期)2022-05-25 13:31:46
农业机械学报(2021年11期)2021-12-07 05:36:44
今日农业(2021年15期)2021-11-26 03:30:27
大气科学(2021年1期)2021-04-16 07:34:18
人大建设(2020年1期)2020-11-17 05:49:22
今日农业(2019年16期)2019-09-10 08:01:36
现代园艺(2017年19期)2018-01-19 02:50:01
农业环境科学学报(2017年2期)2017-03-20 14:57:37
湖南农业(2016年11期)2016-03-07 15:55:35
云南中医学院学报(2015年2期)2015-07-31 18:11:58
