
不同致病力松材线虫和拟松材线虫种群繁殖对温度的响应
2023-05-27 14:12应晨希吴小芹温彤玥
环境昆虫学报 2023年2期
应晨希,吴小芹*,温彤玥,张 岩
(1. 南京林业大学南方现代林业协同创新中心,南京林业大学林学院,南京 210037;2. 江苏省有害生物入侵预防与控制重点实验室,南京 210007)
松材线虫病是我国最具危险性的森林病害。目前该病已在我国19个省(市、自治区)742个县级行政区发生(李硕等,2022),已对我国的松林生态系统造成了严重的破坏,并带来巨大的经济损失。作为我国最重要的林业外来入侵有害生物,松材线虫Bursaphelenchusxylophilus在我国入侵扩张过程中是如何适应环境的目前并不明确。植物病害的发生通常是寄主、病原和环境三者共同作用的结果(Aikawaetal., 2006)。松材线虫种群的增殖与建立成功与否是病害发生的关键,因此研究松材线虫种群的生态适应性显得尤为重要。研究发现,入侵生物对各种环境因子的适应幅度较广,对环境有较强的忍耐力,温度是其中最重要的影响病害发生的环境因子之一(Wangetal., 2005)。Mamiya曾报道松材线虫在低于10℃时不能发育,在28℃以上增殖会受到抑制,在33℃以上不能繁殖;并认为一个地区的年平均气温可作为该地区松材线虫病发生的重要指标,如日本发病最严重的南部地区,年平均温度在15~17℃以上;而日本北部,年平均温度在10~12℃以下,发病轻微(Mamiyaetal., 2004)。Gruffudd等报道欧洲地区夏季平均气温高于20℃时松材线虫病发生的风险就会提高(Gruffuddetal., 2019)。我国也有研究表明,在夏季高温干旱时,该病导致的松树死亡会显著增加,在20℃以下时,松树发病,但不枯死;在18℃以下时,苗木生长正常(Chengetal., 2009)。说明温度对松材线虫病的发生发展影响较大。
有研究发现,松材线虫在单异活体培养时繁殖力较拟松材线虫B.mucronatus强,与致病力呈现正相关性(Tomminen, 1993),这与多数关于松材线虫种群增殖研究的结论相一致(Yuetal., 2012),离体培养时强毒松材线虫种群完成一代的速度比弱毒松材线虫种群和拟松材线虫种群要快得多,较短的生活周期和更高的繁殖率意味着具有更高的种群增长率(徐红梅等, 2017)。但也有研究发现,拟松材线虫在单异活体变温培养条件下,种群增长较松材线虫快(王文琪, 2006; 谢立群等, 2007)。来自北欧的无致病力拟松材线虫较来自美国南部的松材线虫在低温培养下有着更快的繁殖速率(Wangetal., 2005)。由此可见,松材线虫和拟松材线虫的种群增殖与温度的关系仍有待进一步研究。且目前在已有的关于不同温度下松材线虫和拟松材线虫毒力和繁殖力的研究中,多以单异活体(菌食)培养下种群的繁殖量与其毒力相关联;而在病害发生过程中,松材线虫在感病寄主体内(植食)的繁殖能力应能更准确的反映种群的寄生性与致病力的关系。近年松材线虫已向我国北部地区如辽宁等省快速递进(郑雅楠等, 2021)。然目前关于不同温度环境对松材线虫种群繁殖影响的报道仍尚少见。因此,本研究选取不同致病力的松材线虫和拟松材线虫虫株,对其在不同温度下的单异活体培养和寄主体内生存条件下的种群繁殖及其与温度的关系进行探讨,以期解析松材线虫和拟松材线虫在菌食和植食条件下如何响应不同温度而使种群繁衍的生殖对策,为揭示松材线虫的生态适应性和入侵机理提供参考依据。
1 材料与方法
1.1 供试线虫虫株及培养
供试的松材线虫虫株为ZJ和YW4,拟松材线虫虫株为15#和JNL10,均保存于江苏省有害生物入侵预防与控制重点实验室。供试线虫的寄主和来源见表1。
通过马铃薯葡萄糖琼脂(Potato Dextrose Agar,PDA)平板在25℃无光照条件下培养灰葡萄孢Botrytiscinerea4~5 d,待菌丝长满PDA后接入供试线虫,培养5~6 d,待灰葡萄孢菌丝被吃光后采用贝尔曼漏斗法收集线虫(Viglierchioetal., 1983)。将收集的线虫悬浮液离心(4 000 r/min)3 min,弃上清;再用双蒸水(double distilled H2O,ddH2O)清洗3次,4 000 r/min离心,每次3 min,备用。
1.2 供试植物
供试植物为生长状况一致的2年生健康黑松Pinusthunbergii,培养于南京林业大学温室,培养温度控制在28~32℃,相对湿度控制在65%~75%。
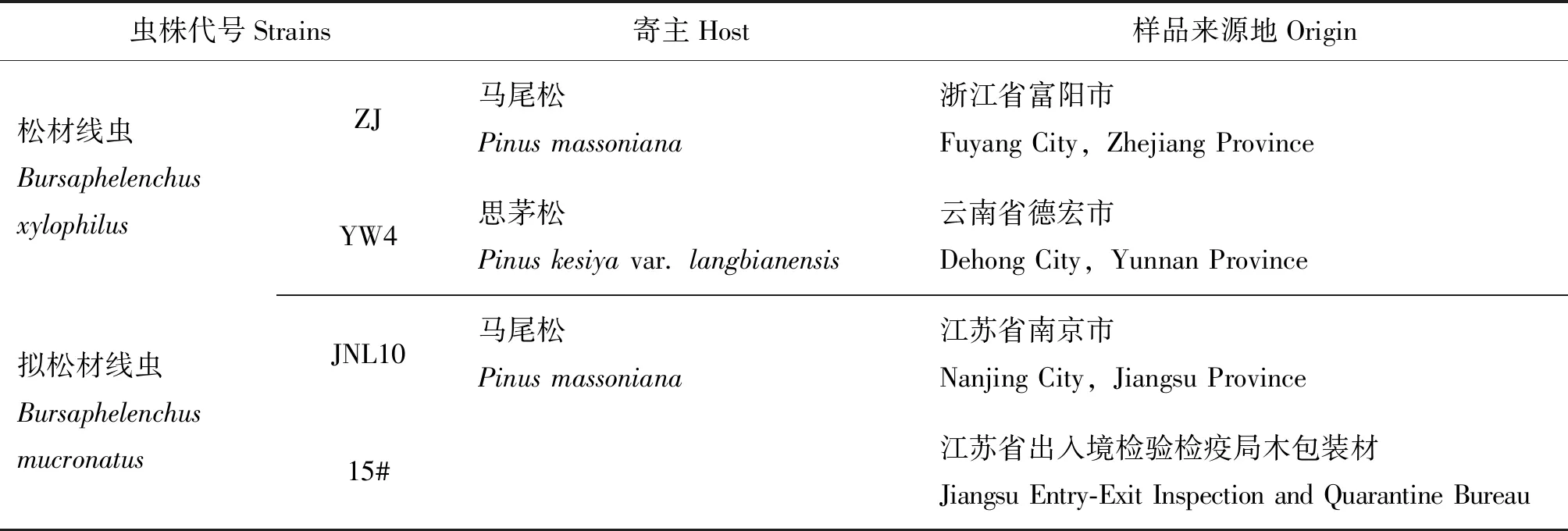
表1 供试松材线虫和拟松材线虫的寄主和来源
1.3 供试线虫致病力的测定
将供试的4个线虫虫株制备成虫悬液(3 000条/mL),以皮接法对黑松盆栽苗进行人工接种,用无菌刀片在近地约10 cm的茎干处割开树皮深达木质部,用镊子取无菌棉球塞入,每株接1 mL的虫悬液,以接种1 mL ddH2O的松树作为对照,封口膜做成漏斗状将切口及棉球包裹住,保证虫液不渗漏。每个处理设置8个重复。根据针叶颜色变化程度,将松树的感病程度划分为以下5个等级:0级:所有针叶均健康呈绿色;Ⅰ级:小部分针叶开始变黄;Ⅱ级:一半左右的针叶变黄或萎蔫;Ⅲ级:大部分针叶褐变萎蔫;Ⅳ级:所有针叶失绿枯萎。在接种后的第20、40和60 d统计松苗的发病率和感病指数,发病率和感病指数按如下公式进行计算(方中达,2007):
发病率(%)=发病株树/总株树×100
感病指数=Σ(各级株树×等级数)/(总株树×发病最高等级数)×100
1.4 不同温度下单异活体培养的线虫卵孵化率的测定
收集经灰葡萄孢培养的4个供试线虫虫株,将虫悬液倒入培养皿中,加入ddH2O静置10 min。后将上层液轻轻倒入到另一培养皿中,线虫的卵会因其特有的粘性附着在原培养皿的底部。重复以上步骤5次,除去虫悬液中的卵。将只含有线虫的虫悬液置于25℃培养箱中培养4 h使线虫产新的卵,收集线虫卵用于试验。
将收集到的供试线虫虫卵置于恒温梯度培养箱(EYELA MTI-201)中,设置15、20、25、30和35℃进行恒温培养,培养皿上层吸附潮湿的吸水纸保湿,各处理设置3次重复。培养48 h后于蔡司显微镜(Zeiss, LSM710,德国)下观察记录线虫卵孵化情况,并计算卵孵化率(初始卵的数量为60~100枚)。
1.5 单异活体培养的线虫繁殖量与后代雌雄性比的测定
挑取各供试线虫虫株的3条雌成虫和3条雄成虫于灰葡萄孢培养基上,置于恒温梯度培养箱中,设置15、20、25、30和35℃进行培养,各处理设置3次重复。培养8 d后,将灰葡萄孢菌丝块切碎,在体式显微镜(Zeiss, Leica MZ16,德国)下测定雌虫和雄虫数量并计算雌雄性比。
1.6 不同温度下寄主黑松中线虫繁殖量和雌雄性比的测定
测定接种前各供试线虫虫悬液的雌雄性比后,将线虫接种到2年生黑松上,接种量和接种方法同1.3所述。接种后的黑松置于恒温梯度培养箱内,分别设置15、20、25、30和35℃恒温培养,每隔3 d浇水一次。培养12 d后,采用贝尔曼漏斗法收集松苗中的线虫,洗净后于显微镜下统计线虫繁殖量和雌雄性比。
1.7 数据分析
使用Microsoft Excel(Microsoft Corp.,Redmond,WA,USA)计算重复的平均值和标准差(standard deviations,SD)作为统计结果,并使用Prism 5软件(北京),采用Duncan多重比较法检验各处理的差异显著性,显著性检验水平为P<0.05。
2 结果与分析
2.1 松材线虫和拟松材线虫的致病力
供试松材线虫和拟松材线虫虫株间的致病力存在差异(表2)。接种后20 d,感染松材线虫的黑松出现发病症状,其中接种ZJ虫株的发病率为75%,感病指数25;接种YW4虫株的发病率为50%,感病指数18。而此时接种拟松材线虫的所有黑松未表现出感病症状。接种后40 d,接种松材线虫ZJ虫株的所有黑松均感病,感病指数69;接种松材线虫YW4虫株的黑松发病率为75%,感病指数34;接种拟松材线虫15#虫株的黑松发病率为12.5%,感病指数6.25;而接种拟松材线虫的JNL10的黑松未表现出症状。接种后60 d,接种松材线虫ZJ虫株的所有黑松100%发病,感病指数85;接种松材线虫YW4虫株的黑松感病率为75%,感病指数34;接种拟松材线虫15#虫株的黑松感病率为12.5%,感病指数6.25;而接种拟松材线虫JNL10虫株的黑松仍未表现出症状。接种无菌水的黑松始终表现健康。可见,供试4个线虫虫株的毒力从高到低依次为:松材线虫ZJ>松材线虫YW4>拟松材线虫15#>拟松材线虫JNL10,且拟松材线虫JNL10对黑松没有致病力(表2)。
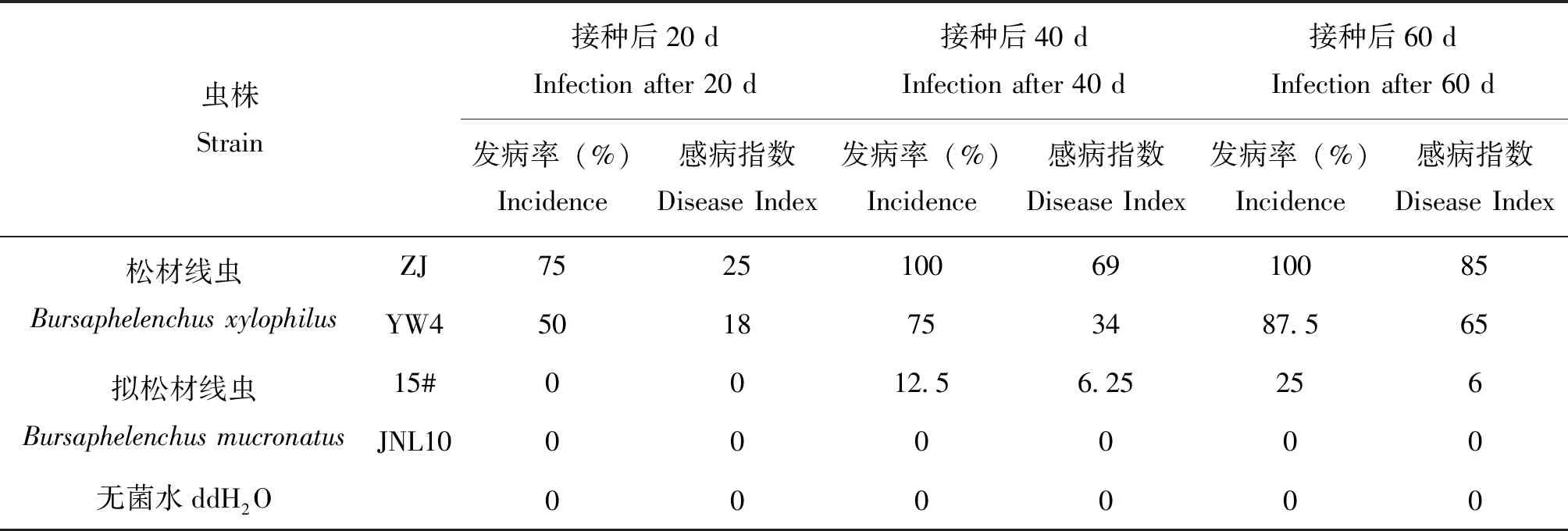
表2 松材线虫和拟松材线虫对黑松致病力测定结果
2.2 不同温度对单异活体培养的松材线虫和拟松材线虫卵孵化率的影响
不同致病力的松材线虫与拟松材线虫在不同温度下的卵孵化结果(图1)表明:供试的4个虫株在5个温度梯度下的孵化率均表现为正态分布的格局。在15℃各个虫株均有少量幼虫孵化出;且随着温度的升高,孵化率逐渐增高,25℃时孵化率达到最大值,之后随着温度升高,孵化率有所下降。孵化率最高的是松材线虫强毒虫株ZJ,在5个温度下均高于其他虫株;在25℃时,其孵化率为73.3%,是各处理中最高的。其次是无毒的拟松材线虫JNL10,在20℃和25℃时的孵化率都仅次于ZJ虫株。4个虫株中孵化率最低的为松材线虫弱致病力虫株YW4,在15℃时其孵化率仅为11.4%,是各处理中最低的,在20℃和25℃,其孵化率也低于其他3个虫株。说明在单异活体培养条件下,供试的松材线虫和拟松材线虫卵的孵化率与致病力并无明显相关性。
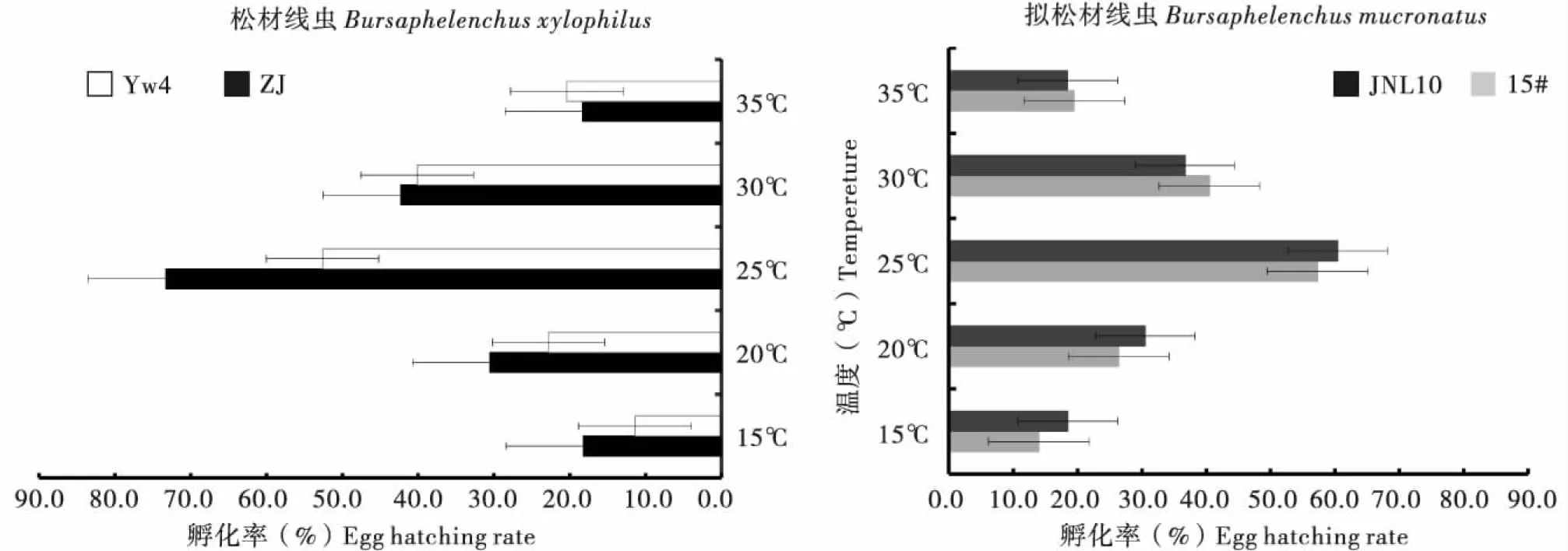
图1 不同温度对松材线虫和拟松材线虫虫株卵孵化率的影响Fig.1 Effects of diferent temperatures on the egg hatching rate of Bursaphelenchus xylophilus and Bursaphelenchus mucronatus
2.3 不同温度下单异活体培养的松材线虫和拟松材线虫种群繁殖量的变化
供试虫株在灰葡萄孢上不同温度下的繁殖量(8 d)测定表明,松材线虫和拟松材线虫在15℃下仅有极少量的2龄幼虫,说明在15℃时,线虫的发育受到抑制,无法正常生长。而在20、25、30和35℃下,各处理都有较多的雌雄成虫产生。供试的4个虫株在20℃时繁殖量较低;在25℃时,繁殖量较大幅度的提升,其中具致病力的拟松材线虫15#虫株增加最多,达到初始虫量的16倍左右。4个虫株出现最大繁殖量的温度梯度有所不同,其中拟松材线虫弱毒虫株15#和无毒虫株JNL10的最大繁殖量出现在25℃,松材线虫强毒虫株ZJ的最大繁殖量出现在35℃,而松材线虫弱毒虫株YW4出现在30℃,且高于30℃时强毒虫株ZJ的繁殖量。观察同时发现,在25~35℃时松材线虫强毒虫株的繁殖量间以及拟松材线虫具毒力虫株的繁殖量间无显著差异(图2)。由此可见,供试的松材线虫和拟松材线虫虫株在菌食条件下有着各自最适的生长温度;松材线虫的弱毒虫株和拟松材线虫同样可耐受高温;且致病力强的虫株并未表现出高的繁殖能力。
2.4 不同温度对单异活体培养的线虫种群雌雄性比的影响
观察发现,在灰葡萄孢培养条件下,除15℃外,松材线虫和拟松材线虫后代中雌成虫数量都多于雄成虫,大部分处理的雌雄性比比值都大于2.0,最小的性比也达到1.85。20℃时各个虫株都有着相对于其它温度下最高的性比,其中拟松材线虫弱毒虫株15#的性比为2.66,为各个处理中最大的。而随着温度的升高,各虫株性比不断下降,后代雌成虫在种群中的比例逐渐降低,雄成虫的数量则相应增加。20℃和25℃时,拟松材线虫的性比均高于松材线虫;30℃和35℃时,松材线虫和拟松材线虫的性比差别不大(除30℃时,拟松材线虫无毒虫株JNL10的性比高于其他虫株)(表3)。
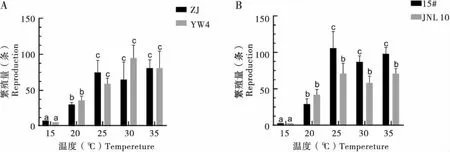
图2 不同温度下在灰葡萄孢上培养的松材线虫和拟松材线虫的繁殖量Fig.2 Reproductive capacity of Bursaphelenchus xylophilus and Bursaphelenchus mucronatus cultivated on Botrytis cinerea at different temperatures注:A,松材线虫;B,拟松材线虫。下图同。Note: A, Bursaphelenchus xylophilus; B, Bursaphelenchus mucronatus. The same below.
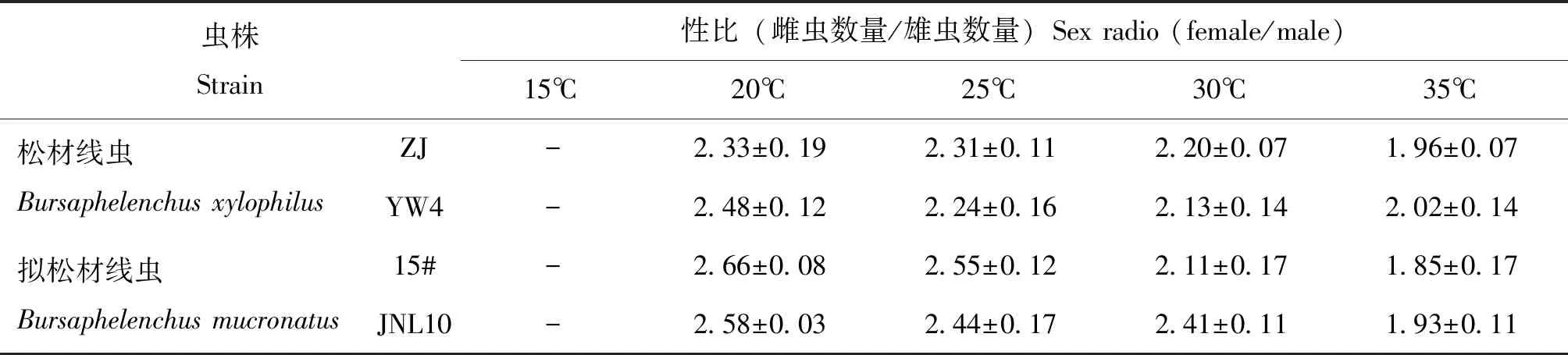
表3 不同温度下单异活体培养的松材线虫和拟松材线虫后代的性比变化
2.5 寄主黑松中松材线虫和拟松材线虫种群在不同温度下的繁殖量变化
松材线虫和拟松材线虫接种黑松12 d后的繁殖量测定表明:松材线虫强致病力虫株ZJ较弱致病力虫株YW4和其它2个拟松材线虫虫株表现出很高的繁殖力;在15℃下,松材线虫在寄主黑松中繁殖量约为500条,拟松材线虫在寄主黑松中繁殖量约为280条,均大大高于它们在灰葡萄孢上的繁殖力;随着温度的上升,强毒虫株ZJ的繁殖量不断增加,在30℃的黑松中达到了最高,较初始虫量增长了5倍;在35℃时其繁殖量有所下降,但还是保持在较高的水平。松材线虫弱毒虫株YW4在25℃有着最高的繁殖量,且在各个温度下,YW4虫株的繁殖量均低于强致病力的松材线虫ZJ虫株。拟松材线虫在黑松体内的繁殖力明显低于松材线虫,且也低于其在灰葡萄孢上的繁殖力;其中繁殖量最低的为无致病力的拟松材线虫JNL10,在25℃时最大繁殖量为452条,较初始虫量只增长1倍左右;拟松材线虫有毒虫株15#的繁殖量随温度增高有所增加,但在20℃至35℃时无显著性差异。表明低致病力和无致病力的拟松材线虫虫株在黑松体内并不具有种群繁殖优势;在寄主体内,不同温度条件下的松材线虫和拟松材线虫的繁殖力与其致病性存在正相关的关系,即在寄主松树中,致病力越强的虫株,其繁殖力越大。结果同时表明,4个线虫虫株在35℃时的繁殖量都出现一定的下降,说明高温对种群的增殖有一定的抑制作用(图3)。
图3 不同温度下黑松体内松材线虫和拟松材线虫的繁殖量Fig.3 Reproductive capacity of Bursaphelenchus xylophilus and Bursaphelenchus mucronatus in Pinus thunbergii at different temperatures
2.6 不同温度对寄主黑松中松材线虫和拟松材线虫种群性比的影响
松材线虫和拟松材线虫4个种群的初始性比为2.2~2.4,接种黑松12 d后发现,各线虫种群的性比较接种前都有较大程度的下降(表4)。与单异活体培养下较高的性比不同,在黑松体内各个虫株的种群性比都低于1.5。其中松材线虫虫株ZJ和YW4种群在15~35℃下大多处理的性比均低于1.0,出现了雄成虫数量多于雌成虫的情况;相对于供试的拟松材线虫虫株,松材线虫虫株的性比处于较低值。从各温度处理来看,在寄主松树中松材线虫不同毒力虫株的性比在20~30℃较大些,但总体随温度变化的幅度不大;而拟松材线虫15#和JNL10种群的性比率则随着温度的升高而增大,变化趋势明显,均在35℃时最大,其中无致病力的JNL10种群的性比率最高达1.42。在各温度处理下的寄主松树中,拟松材线虫无论是毒力虫株还是无毒虫株其性比均大于1.0,而其繁殖量则处于较低的水平(表4)。
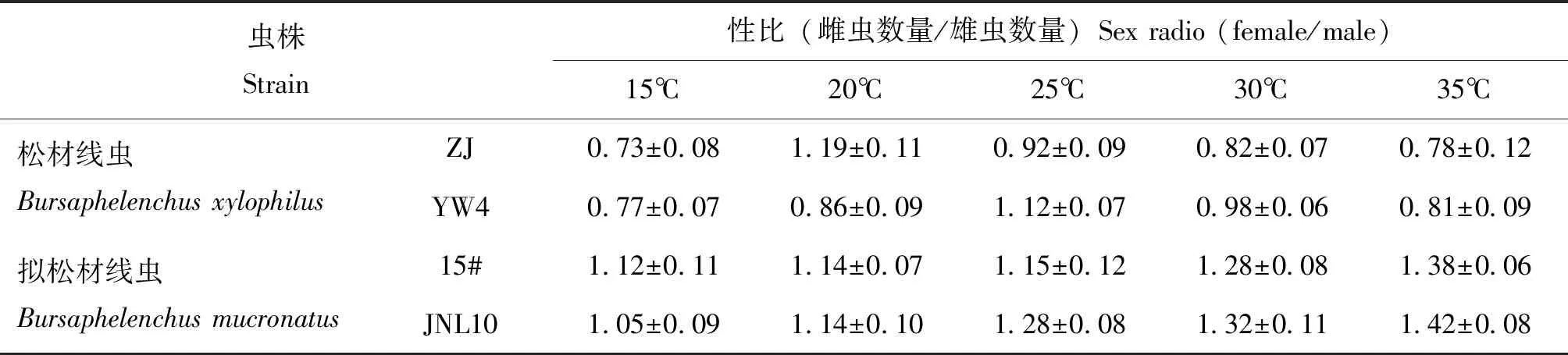
表4 不同温度下寄主松树中松材线虫和拟松材线虫后代的性比变化
3 结论与讨论
温度对入侵生物的生长繁殖和定殖扩张至关重要。且在无资源因素限制的环境中,种群增长率与生物世代周期、产卵量和雌虫存活率等参数密切相关(谢立群等, 2007)。温度对松材线虫增殖有着重要作用(张星耀等, 2003;Zhaoetal., 2007; 盛若成等, 2019;)。因此,研究影响松材线虫种群增殖的因子有助于掌握病害的发生发展规律。本研究探讨了松材线虫和拟松材线虫的卵孵化对不同温度的响应,表明供试虫株的最适卵孵化温度均为25℃;而在15℃和35℃下,线虫卵的孵化受到抑制,孵化率降低。研究还发现25℃下48 h孵化出的幼虫大多数处于J2阶段,而在35℃下孵化率虽低但已有较多J3幼虫出现,由此可推测,35℃下线虫生殖世代周期缩短,这与张建平等的研究结论相吻合(张建平, 2002)。供试的4个虫株,致病力虽有较大差异,但在单异活体培养下卵的孵化率与致病力并未呈现一致性,对温度的适应性也无明显差异,在不同温度梯度下,呈现出相同的变化趋势。
本研究发现在单异活体培养的菌食条件下,拟松材线虫弱毒种群的繁殖量超过松材线虫,且松材线虫和拟松材线虫各虫株有着各自的最适生长温度。松材线虫强毒虫株ZJ的离体培养最适温度为35℃,在供试4个虫株中耐热性最高;拟松材线虫的最适增长温度相对较低,繁殖量最高的拟松材线虫弱毒虫株15#最适温度为25℃,随着温度升高其繁殖量有所下降但仍保持在相对较高的水平。说明在菌食条件下不同毒力的松材线虫和拟松材线虫的繁殖力对温度的响应不同,且致病力与繁殖力并未呈现正相关性。这与Wang等观察到的结果有所不同(Wangetal., 2005)。研究还发现当培养温度在15℃时,单异活体培养的松材线虫和拟松材线虫种群的繁殖受到抑制,线虫种群表现出不能增长。当培养温度达到30℃时,松材线虫强毒虫株ZJ种群增长较快,说明较高温有利于松材线虫繁殖。
松材线虫和拟松材线虫为两性生殖线虫,理论上其后代性比为1∶1。Rutherford等研究发现拟松材线虫单异活体培养时种群性比率最高为2.0,在某些重复中,性比出现过极端的50∶1的情况(Rutherfordetal., 1990)。本研究中,在单异活体各不同温度培养时,不同毒力的松材线虫和拟松材线虫种群的性比率多介于2.0~3.0之间。在20℃下,各个种群的性比是最高的,随着温度的升高性比呈现出不断下降的趋势,雄虫在种群中所占的比例不断提高。与此同时,种群的繁殖量较20℃时也有着较大的提高。本研究对离体恒温梯度培养下的线虫增殖观察时间为8 d,若延长观察时间,通过较长时间的世代繁殖,种群的性比可能会有一定变化。
在自然条件下,松材线虫多以寄主植物薄壁细胞为营养来源(Matsunagaetal., 2003; Mamiyaetal., 2004)。Yamaguchi的研究表明,在25~30℃下,黑松幼苗中松材线虫的繁殖是稳定的,且随着温度的升高呈指数增长;但当温度在20℃以下不足以促进松材线虫的发育时,松材线虫繁殖就可能受到宿主防御系统的抑制(Yamaguchietal., 2020)。本研究发现,在寄主黑松体内的植食条件下,在15℃下,松材线虫种群的繁殖量约为500条,拟松材线虫的繁殖量约为280条;随着温度的升高线虫种群的繁殖量上升;松材线虫多数处理的雌雄性比低于1.0,且随温度升高不断微调下降;而拟松材线虫种群的繁殖量随温度升高增长缓慢,其性比1.0~1.5,呈现上升趋势。Mamiya等研究发现松材线虫在低于10℃会出现滞育现象,夏季的高温和生长季的干旱有利于松材线虫病的发生(Mamiyaetal., 2004)。本研究说明松材线虫和拟松材线虫对温度的响应不同,且随温度升高,树体中线虫的繁殖量增加。已有多项研究预计由于全球变暖导致的温度变化将会进一步加剧松材线虫病的发生和扩展(Hirataetal., 2017; Ikegamietal., 2018; Gruffuddetal., 2019)。本研究还表明在寄主松树中,线虫在较低温下仍能维持一定的种群数,且松材线虫的低温适应性高于拟松材线虫,松材线虫的这种温度适应性有助于解释其引起的病害在我国南北方的扩展蔓延态势。
本研究中各虫株在寄主中的繁殖量和其致病力呈现显著的正相关性。即在寄主松树体内松材线虫的种群繁殖力明显高于拟松材线虫,且毒力愈强差异愈大。高繁殖量的生物通常通过提高后代产量与投入能量的比值以达到提高生殖效率的目的(Mamiyaetal., 1977)。松材线虫和拟松材线虫种群的后代性比随生存条件变化而产生相应的调整,这种能力可能是种群繁殖力产生差异的原因。在营养条件充分时,线虫种群中雌虫数量的增加可使得种群数量增长,但雌虫消耗的营养随之增多,因此在线虫种群中存在着一种“平衡”以制约雌虫的增长。推测强致病力的松材线虫在遇到寄主及温度胁迫时可通过降低种群中的雌虫比和增加雄虫比使得种群保持持续增长,这与其在真菌营养下的菌食阶段显著不同。而拟松材线虫在寄生条件下,种群性比率保持较高,其种群数量则增长缓慢。由此推断,松材线虫种群可以通过调节雌雄虫在种群中所占的比例来应对寄主和环境条件的改变,使得种群增长保持在相对较高的水平而促进病害的发生。本研究也暗示随着营养和环境压力的增加,那些能够使性比下降、雄虫数量在种群中所占比例增加的种群将更具繁殖力,同时也更具致病力。
猜你喜欢
贵州农业科学(2023年5期)2023-05-17
美食(2022年12期)2022-12-09
中国蚕业(2022年1期)2022-03-09
今日农业(2021年6期)2021-11-27
绿色中国(2019年13期)2019-11-26
美食(2019年7期)2019-09-10
科学大众·小诺贝尔(2018年12期)2018-02-22
现代园艺(2017年21期)2018-01-03
现代园艺(2017年21期)2018-01-03
生物灾害科学(2015年2期)2015-03-11
