
豆野螟的生物防治研究进展与展望
2023-05-27 14:00王嘉乐朱佳晨江薰垣徐汉虹唐雪妹
环境昆虫学报 2023年2期
王嘉乐,朱佳晨,江薰垣,徐汉虹,唐雪妹*
(1. 广东省农业科学院农业质量标准与监测技术研究所/广东省农产品质量安全风险评估重点实验室/农业农村部农产品质量安全检测与评价重点实验室,广州 510640;2. 浙江大学昆虫科学研究所,杭州 310058;3. 华南农业大学植物保护学院/天然农药与化学生物学教育部重点实验室,广州 510642)
豆野螟MarucavitrataFabricius,又名豇豆荚螟、豆蛀螟、大豆卷叶螟、豆叶螟,属鳞翅目Lepidoptera草螟科Crambidae,是豇豆等豆类蔬菜的重要害虫。1995年以前,豆野螟曾用学名MarucaaquatilisDoisduval(林荣华等, 2000)和MarucatestulalisGryer。国外众多的翻译资料将MarucatestulalisGryer(豆荚野螟)译作豆荚螟,因而常与EtiellazinckenellaTreitschke(豆荚螟)混淆。林荣华等(2000)曾就该问题提请研究学者们注意,但在此后的相关研究报道中,仍然常见此类误用。
豆野螟喜产卵于寄主花(蕾)(田忠等, 2017),卵期3~5 d后,初孵幼虫在寄主花器内取食,2~3龄幼虫通过吐丝将受害花器与近花端的果枝相连,转移至豆荚内为害(柯礼道等, 1985)。老熟幼虫多钻入土缝内做土室结荚化蛹,少数在植株上或豆架杆内化蛹(王琳等, 2003)。目前已知豆野螟为害的寄主超过73种,主要包括豇豆Vignaunguiculata、绿豆V.radiata、四季豆Phaseolusvulgaris、豌豆Pisumsativum、木豆Cajanuscajan、大豆Glycinemax等荚面少毛的豆类作物。其中以豇豆受害最为严重,可致产量损失高达72%(Sharma, 1998)。近年来,随着高产优质品种的推广,豇豆栽培面积不断扩大,且全球气候变暖,国际贸易交流日益频繁,豆野螟在亚洲与非洲的发生日趋猖獗(Srinivasanetal., 2021)。对我国南方地区尤其是海南、广东、广西等(亚)热带气候地区的豆类蔬菜生产构成严重威胁,田间常见“十荚九蛀”的情形。
目前,化学农药仍是国内外防治豆野螟的主要方式,引发了豇豆等豆类蔬菜中尤为突出的质量安全问题。豆野螟幼虫具有钻蛀取食的习性,能有效躲避多种杀虫剂的喷施,导致施药效果不佳,国内外超范围使用农药的报道屡见不鲜(Schreinemachersetal., 2017)。且豇豆花果同期、采摘间隔短,成为了我国乃至全球农药残留超标问题最严重的食用农产品之一。自2010年海南“毒豇豆”事件以来,国家高度重视豇豆等蔬菜农药残留治理问题。2021年5月,农业农村部联合六部委印发了《食用农产品“治违禁 控药残 促提升”三年行动方案》,将“豇豆”列作农药残留突出问题重点治理蔬菜之首;2022年11月农业农村部办公室印发《豇豆农药残留突出问题攻坚治理方案》,大力推进豇豆病虫害绿色防控技术的应用与普及。
生物防治是豆野螟绿色防控体系的重要组成部分,通过利用害虫天敌(寄生性与捕食性昆虫)、病原微生物、植物等生物及其产物,能有效抑制豆野螟的种群数量和抗药性的产生(Sreelakshmietal., 2015)。其安全、绿色、可持续的优点突出,高度符合新时代下人民对农产品质量安全的新需求和新期待,因此近年来备受广大研究学者的重视。然而,豆野螟不属于我国重大农业害虫或主要粮食作物害虫,在国内害虫生物防治研究领域关注度较低。迄今,国内鲜有关于豇豆重要害虫豆野螟生物防治技术的系统综述,在指导田间绿色防控时缺乏重要参考。因此,本文将对国内外生物防治豆野螟的前沿技术与研究成果进行简要论述,主要围绕昆虫性信息素、天敌昆虫资源、昆虫病原微生物与植物源农药4个方面展开,以期为豆野螟生物防治技术的推广应用和基础性研究提供参考。
1 昆虫性信息素
性信息素是昆虫两性交流的重要信号分子,由性信息素腺分泌,可引起同种昆虫的异性群体产生生殖反应。因其具有物种特异、微量高效、无毒无害的特点,近三十年在欧洲地区被广泛应用于昆虫种群监测、害虫大规模诱捕、害虫交配干扰等方面,成效显著(Witzgalletal., 2010)。
目前已鉴定的豆野螟雌性性信息素共有3种组分,分别是(反,反)-10,12-十六碳二烯醛(EE10,12-16: Ald)、(反,反)-10,12-十六碳二烯醇(EE10, 12-16: OH)和反-10-十六碳单烯醛(E10-16: Ald)(Luetal., 2013; Schlägeretal., 2015)。 总体而言,EE10, 12-16: Ald为豆野螟性信息素的主要组分,EE10, 12-16: OH和E10-16: Ald的含量普遍较低。不同地理种群雌蛾释放的性信息素中, 各组分的比例不尽相同(图1)。例如, 在加纳、 泰国、 越南、 我国台湾的豆野螟种群的性信息素中未鉴定到E10-16: Ald(Schlägeretal., 2015),但该组分在我国武汉种群中大量存在, 在我国化州种群中的含量也显著高于EE10, 12-16: OH(Luetal., 2013)。由此推测,不同地理种群的遗传差异可能导致了豆野螟性信息素组成的分化,促进了地理隔离的形成。
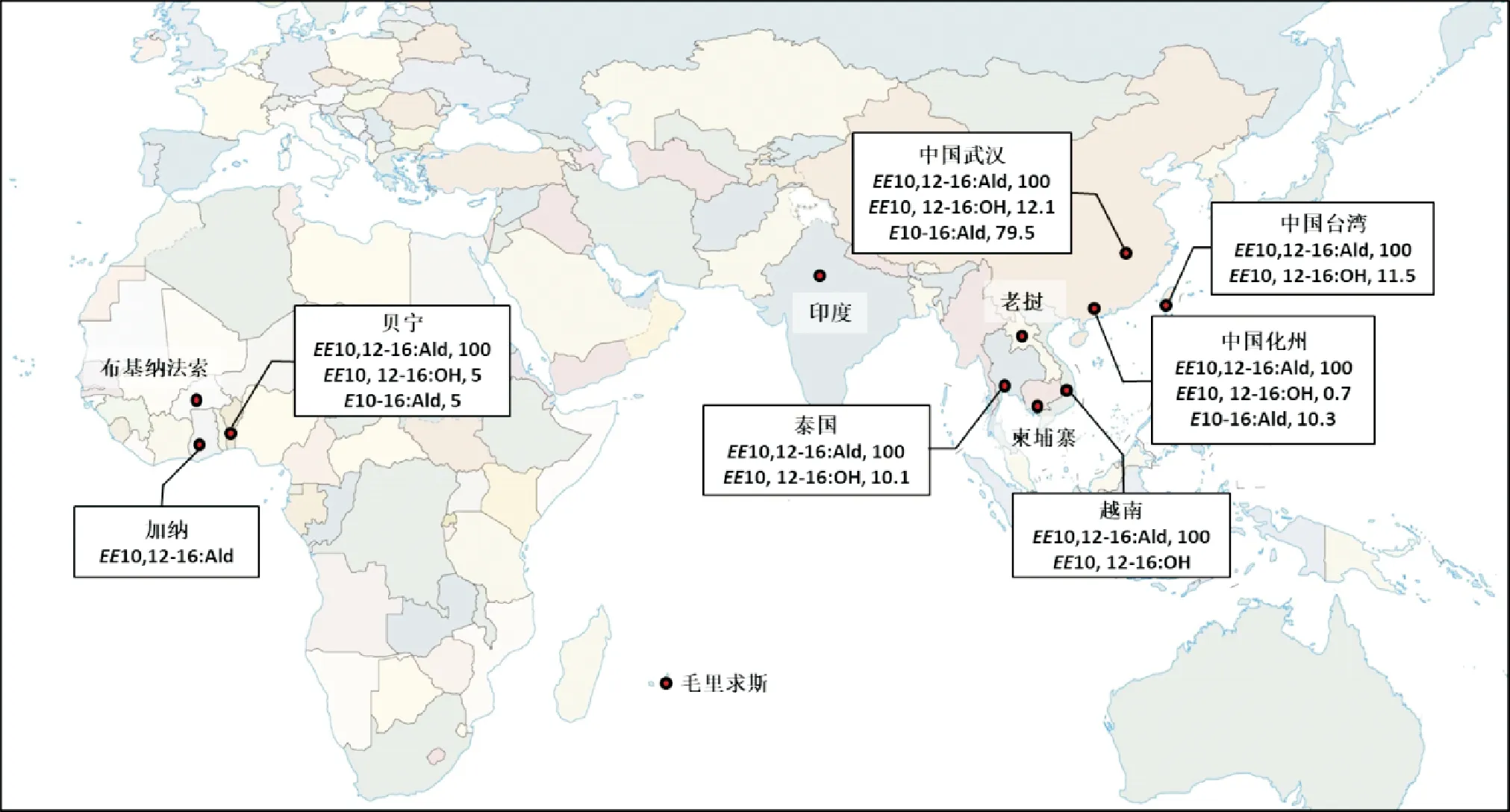
图1 不同地理种群豆野螟性信息素组成的差异Fig.1 Differences of sex pheromone components in different geographic populations of Maruca vitrata注:该图基于国家自然资源部标准地图服务网站下载的审图号为GS(2016)1663号的标准地图制作,且未对国家、行政区域界线或者范围、重要地理信息数据进行修改。Note: This figure was drawn based on the standard map No. GS (2016) 1663 downloaded from the standard map service website of the Ministry of Natural Resources, without modification of national and administrative boundaries or scope, as well as important geographical information data.
不同地理种群的豆野螟雄蛾对不同雌性性信息素的趋性也有所差异。在西非国家贝宁和加纳,以100∶5∶5的比例混合EE10, 12-16: Ald、EE10, 12-16: OH和E10-16: Ald,能有效诱集豆野螟雄蛾;而在布基纳法索,单独使用EE10, 12-16: Ald才是诱集雄蛾最有效的方式(Downhametal., 2003; Downhametal., 2004)。有趣的是,这两种使用性信息素的方法在毛里求斯(Unmole, 2009)及多个东南亚国家或地区,如泰国、越南、我国台湾等均无法吸引豆野螟雄蛾(Schlägeretal., 2012; Srinivasanetal., 2015)。使用3种人工合成性信息素ZE-10, 12: Ald(EE-10, 12: Ald的同分异构体)、EE10, 12-16: OH与E10-16: Ald,以100∶10∶5的比例混合制作诱芯,在印度和柬埔寨能显著提升豆野螟雄蛾的诱集数量(Bhanuetal., 2018; Kapoor and Shankar, 2019);但该方法在老挝、泰国、越南、我国台湾仍不适用(Srinivasanetal., 2019a)。在我国武汉和广州,分别以5∶0.3∶5和5∶5∶0.7的比例配制EE10, 12-16: Ald、EE10, 12-16: OH和E10-16: Ald三元组分诱芯时,田间诱捕豆野螟雄蛾的活性最高(陆鹏飞, 2007)。从分子水平上看,豆野螟雄蛾对不同性信息素组合的差异反应可能与性信息素结合蛋白(Pheromone binding protein,PBPs)关键位点的氨基酸突变有关(Malinietal., 2019)。
性信息素结合蛋白(PBPs)和普通气味结合蛋白(general odorant-binding protein, GOBPs)是昆虫气味结合蛋白(odorant binding protein, OBPs)家族的两个重要亚族,分别参与性信息素分子和寄主植物挥发物(信息素)的识别与结合。目前已知的豆野螟PBPs共3种,分别是在雄蛾中鉴定到的MvitPBP1、MvitPBP2和雌蛾中的MvitPBP3(Malinietal., 2013)。荧光竞争结合实验结果显示,MvitPBP1、MvitPBP3与性信息素EE10, 12-16: Ald的结合力最强,MvitPBP2则偏好E10-16: Ald(Maoetal., 2016)。Wang等(2014)从豇豆花中鉴定出17种能够引起豆野螟雌虫触角强烈的EAG反应的挥发性化合物,解释了豆野螟对寄主植物花(蕾)的产卵偏好性。Zhou等(2015)从分子生物学水平验证了MvitGOBP1和MvitGOBP2能与17种豇豆花挥发物发生不同程度的结合。Ai等(2021)鉴定的MvitOBP3则与4种豇豆花挥发物具有较强的结合亲和力。此外,MvitPBP3被检测出与MvitGOBP2相似的功能,能与豇豆花挥发物丁酸辛酯显著结合(毛阿萍, 2015)。因此,豆野螟性信息素和寄主植物的挥发性化合物均可作为引诱剂(或增效剂),用以调节豆野螟在寻找寄主植物或产卵时的选择行为,为生物防治提供新的思路。
现阶段,我国对昆虫性信息素等引诱剂的应用总体仍处于起步阶段。有鉴于不同地理种群(或亚种)的豆野螟性信息素通讯系统存在差异,全国各地在考虑利用性信息素防治豆野螟时,有必要对当地种群的性信息素组分进行再鉴定,进而推广因地制宜的豆野螟性信息素应用技术规程。
2 天敌昆虫资源
天敌昆虫在作物害虫生物防治中扮演着重要的角色。根据取食特点的不同,常见的天敌昆虫主要分为寄生性与捕食性两大类群。
2.1 寄生性天敌
寄生性天敌昆虫通常将卵产在寄主害虫的体内(或体外),其后代通过持续消耗寄主以满足自身生长和发育所需,从而有效控制田间害虫基数,是害虫生物防治的重要手段之一。据统计,自然界中已知的豆野螟寄生性天敌昆虫超过98种(Srinivasanetal., 2021)。其中,膜翅目寄生蜂是重要的天敌昆虫资源。豆野螟寄生蜂物种以茧蜂科数量最多,姬蜂科次之(表1)。
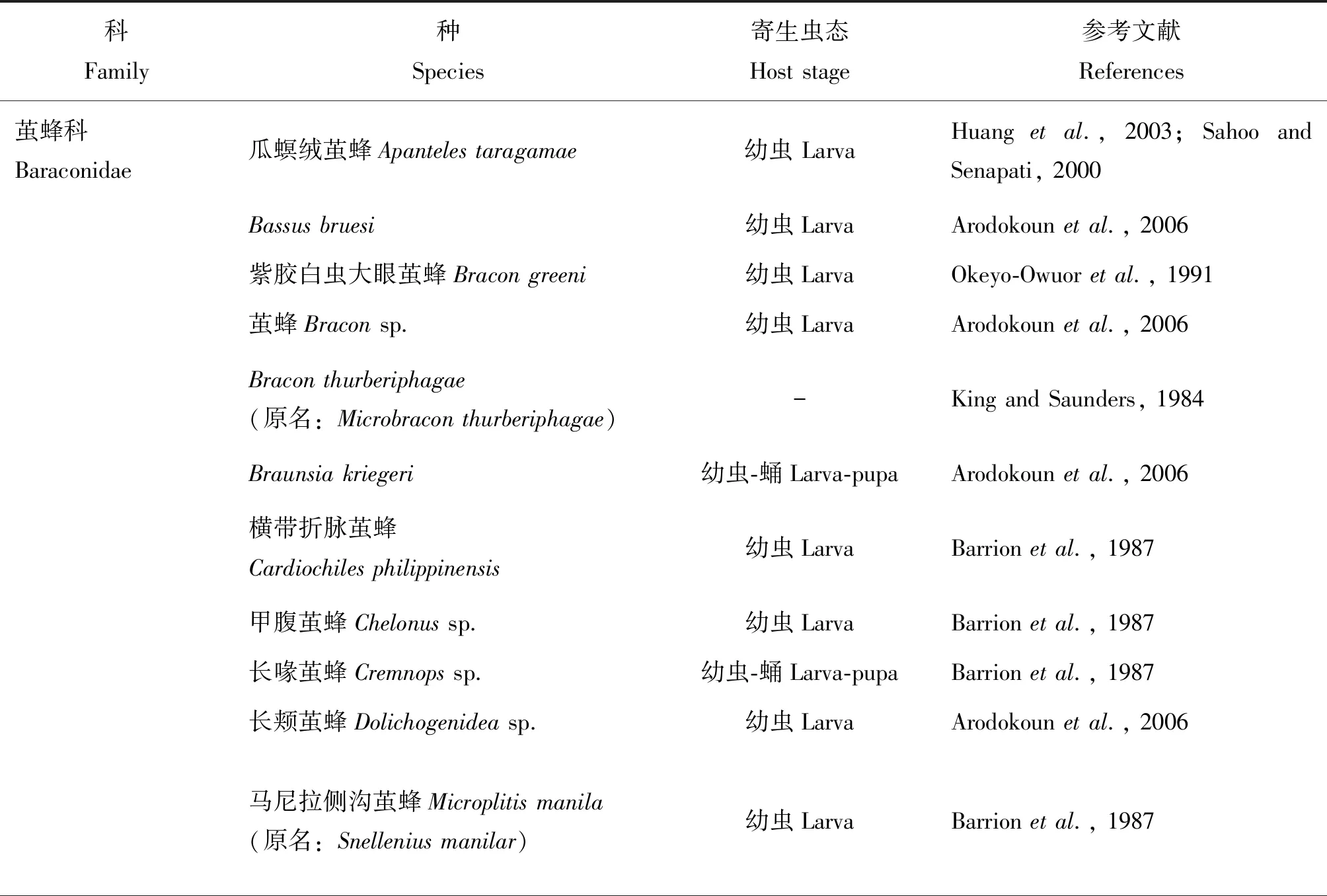
表1 国内外已报道的豆野螟寄生蜂物种

续表2 Continued table 2
此外,双翅目寄生蝇也是基于天敌昆虫生物防治豆野螟的有效途径,国内外研究报道中常见的豆野螟寄生蝇共13种(表2)。

表2 国内外报道的豆野螟寄生蝇种类
豆野螟的寄生性天敌昆虫资源虽然丰富,但至今未发现其专性寄生的天敌。瓜螟绒茧蜂Apantelestaragamae是豆野螟的优势寄生蜂,起源于印度-马来西亚地区,在我国海南、浙江等地均有分布(刘树生等, 1989; 刘奎等, 2007)。寄生时,雌蜂产卵在豆野螟1~2龄幼虫(2~3日龄)体内,老熟幼虫自3龄豆野螟幼虫体内钻出并吐丝结成白茧,约3~4 d后羽化成蜂(Dannonetal., 2010)。瓜螟绒茧蜂在我国台湾地区田菁上的自然寄生率可高达63%(Huangetal., 2003),在豆野螟生物防治领域具有广阔的应用前景。然而,目前国内外对瓜螟绒茧蜂的认识仍然十分有限。我国大陆仅在调查瓜螟天敌时,对其发生动态进行了简要报道(刘奎等, 2007);国际上围绕瓜螟绒茧蜂的生物学特性开展了一些研究,揭示了印度种群与台湾种群在寄主范围与寄生习性上的差异(Mohan and Sathiamma, 2007),但对其寄生机制(包括寄生因子等)仍缺乏深入了解。
为充分发挥寄生蜂天敌资源的应用价值,贝宁曾从我国台湾引进瓜螟绒茧蜂用于豆野螟的生物防治,但一直未能成功建立起本地的优势种群(Srinivasanetal., 2021)。直至2016年,贝宁再次引进了豆野螟的两种茧蜂Phanerotomasyleptae和Therophilusjavanus,并成功克服了环境等制约因素,建立了本地种群并将其应用推广至邻国布基纳法索(Baetal., 2019; Srinivasanetal., 2021)。其中,P.syleptae是目前已知为数不多的豆野螟卵-幼虫期寄生蜂,可在豆野螟幼虫钻蛀为害前完成寄生,而T.javanus是幼虫期寄生蜂,两者相辅相成,是豆野螟绿色防控体系建设的重要组成部分。但截至目前,P.syleptae和T.javanus在我国未见纪录。我国科研人员报道了黄眶离缘姬蜂Trathalaflavoorbitalis是豆野螟的重要寄生蜂,7-11月在长沙的自然寄生率达40%左右(肖芬等, 2009),该数据与7-8月在贵阳调查结果相近(忙定泽等, 2012)。
2.2 捕食性天敌
已知豆野螟捕食性天敌的种类较为丰富(表3),但实际活跃在豆类蔬菜田间的种群数量较少。除了室内日捕食卵量可达10~15粒的小花蝽若虫(汪自卿等, 1987)以外,其他捕食性天敌大部分以多种昆虫为食,在豆类蔬菜田间捕食豆野螟的幼虫。然而,豆野螟幼虫钻蛀豆荚取食的习性为天敌的捕食行为增加了难度,天敌可能因此转向捕食其他昆虫。因此,应用捕食性天敌绿色防控豆野螟存在一定的短板。
我国幅员辽阔、自然地理条件复杂,天敌昆虫物种资源极为丰富,且在赤眼蜂、瓢虫、蠋蝽等重要天敌昆虫的规模化繁殖与应用技术创新方面都取得了一系列理论突破。但迄今,我国除台湾地区鲜有关于豆野螟天敌资源的监测与调查研究,严重制约了基于天敌昆虫应用的豆野螟生物防治技术发展。
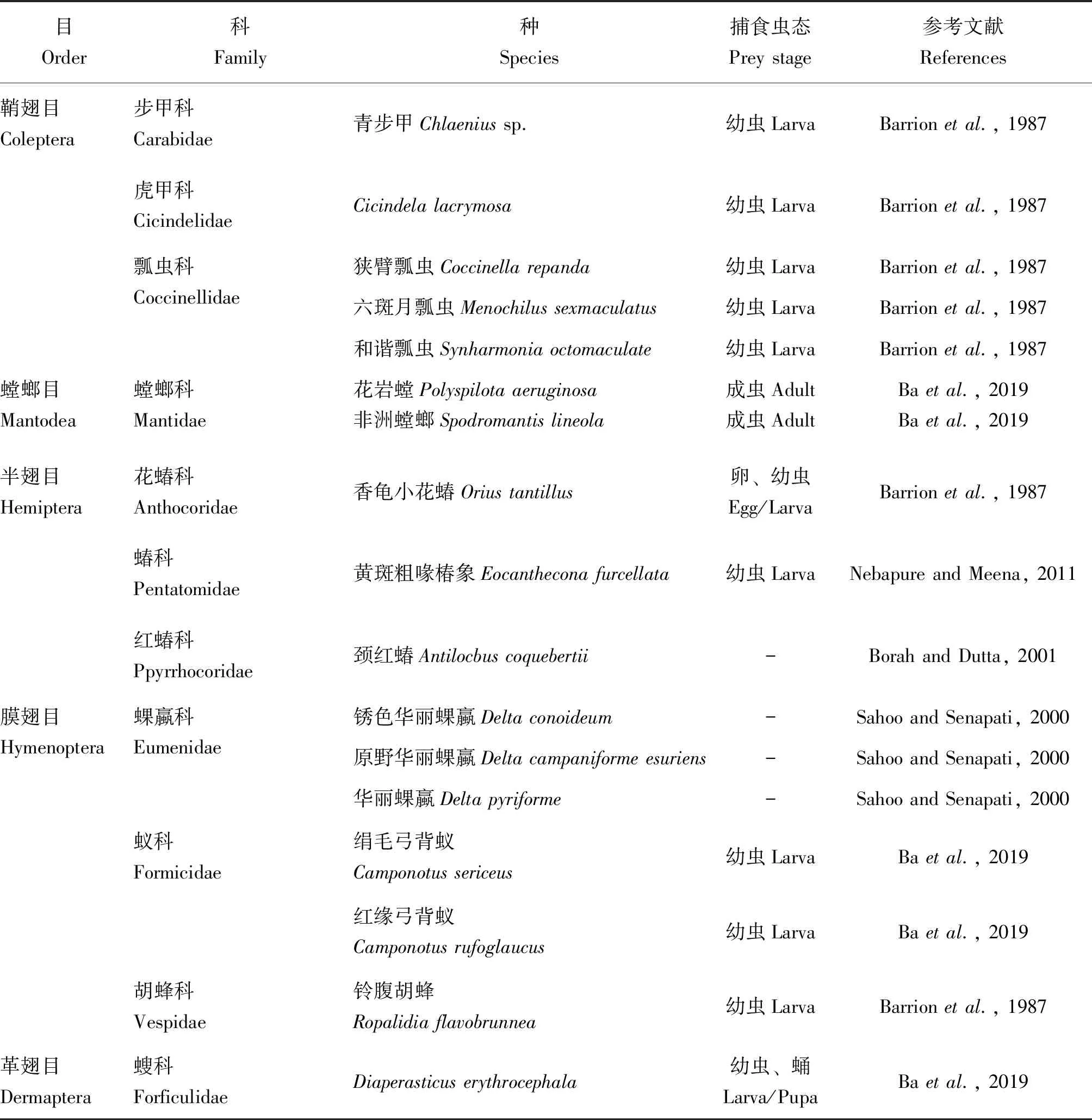
表3 国内外报道的豆野螟捕食性天敌昆虫种类
3 昆虫病原微生物
昆虫病原微生物是能够侵染昆虫使其致病,并在昆虫种群中流行的真菌、细菌、病毒等。它们为提高自身在寄主体内适应性,可调控寄主生理引起发烧等病理现象;而寄主在应对昆虫病原物侵入时也会产生免疫反应,改变其取食、繁殖、防御、群集、趋性等行为。因此,昆虫病原微生物作为调控害虫种群数量动态的重要因子,在害虫生物防治中发挥了关键作用。
3.1 真菌
昆虫病原真菌是昆虫病原生物中最大的类群,能有效防控豆野螟的发生。已有报道证实,球孢白僵菌Beauveriabassiana和金龟子绿僵菌Metarhiziumanisopliae对豆野螟具有中高致病性(Ekesietal., 2002; Mehintoetal., 2014; Srinivasanetal., 2014; Tumuhaiseetal., 2015);但其制剂对豆野螟的防治效果并不理想。在实验室条件下,仅越南报道了上述两种真菌制剂对豆野螟具有较高致病性(Srinivasanetal., 2014);印度和泰国报道的真菌制剂侵染效果则不佳(Sunithaetal., 2008, Yule and Srinivasan, 2013)。根据现有的田间试验数据,绿僵菌制剂的防治效果略高于白僵菌。在印度卡纳塔克邦,按照(1~2)×1012conidia/ha在豇豆上使用绿僵菌,可减少45%~68%的蛀果率(Rachappaetal., 2005);在柬埔寨的裙带豆上使用Real M-62©、Real M-69©的绿僵菌制剂可减少48%~77%(Srinivasanetal., 2019b)。但在印度泰米尔纳德邦,按照2.8×106CFU/g在黑吉豆上使用白僵菌,可挽回的蛀果损失不超过50%(Soundararajan and Chitra, 2011)。
3.2 细菌
豆野螟是多种昆虫病原细菌的自然寄主。苏云金芽孢杆菌Bacillusthuringiensis(Bt)、蜡样芽孢杆菌B.cereus、粪链球菌Streptococcusfaecalis和粘质沙雷菌Serratiamarcescens等均可侵染豆野螟(Otienoetal., 1983)。其中,Bt产生的伴孢晶体中内含δ-内毒素,可经虫体中肠蛋白酶水解进入血淋巴,引起败血症,致虫体死亡。在西非和我国台湾,豆野螟对Bt毒素Cry1Ab和Cry1Ca高度敏感(Srinivasan, 2008)。此外,不同地理种群的豆野螟对不同Bt亚种制剂的敏感性也有差异。在泰国,豆野螟对苏云金芽孢杆菌库尔斯塔克(Kurstaki)亚种制剂高度敏感(Yule and Srinivasan, 2013; Kumaretal., 2014);而在越南和我国台湾地区,则对苏云金芽胞杆菌鲇泽(aizawai)亚种制剂高度敏感(Srinivasanetal., 2014)。不同地理种群豆野螟对Bt敏感性的差异可能与其长期生境中的Bt水平有关,部分地理种群可能已经对Cry1Ab和Cry1Ca产生了一定的抗性。目前尚未建立豆野螟对Bt毒素的敏感基线。
Bt制剂应用于田间防治豆野螟的案例相当丰富。以裙带豆为例,在泰国使用Bt-aizawai(Zitarback F.C.©)和Bt-kurstaki(Redcat©),能分别减少36%~52%和36%~45%的蛀果率(Yule and Srinivasan, 2013);在柬埔寨使用Bt-aizawai(Xentari©)和Bt-kurstaki(Crymax©、E911©)则能减少46%~72%和48%~75%(Srinivasanetal., 2019b)。
3.3 病毒
目前已知的能侵染豆野螟的昆虫病原病毒数量不多。1986年,国内首次采集、分离到豆野螟颗粒体病毒(M.testulalisgranulosis viruses, MtGV)。幼虫在感染第3天后停止取食,四处爬动但行动和发育都变得迟缓,体色渐变成黄白色,体壁脆弱(叶爱珍等, 1998)。随后,研究人员发现大蜡蛾核型多角体病毒(Galleriamellonellanucleopolyhedrovirus,GmNPV)也能侵染豆野螟幼虫,但认为其在田间规模化应用的前景较为有限(Parthasarathyetal., 2004)。直至2004年,在我国台湾地区的田菁上首次发现了NPV特异性感染的豆野螟幼虫。被感染的幼虫行动缓慢不进食,虫身呈粉红色且体壁易破,表现为典型的NPV感染(Leeetal., 2007)。经表征分析,该病毒被命名为豆野螟多核衣壳型多角体病毒MaviMNPV(M.vitratamultiple nucleocapsid nucleopolyhedrovirus)。同其他NPV一样,MaviMNPV制剂对豆野螟低龄幼虫非常有效(Leeetal., 2007),可在幼虫转移至豆荚为害前实现有效控制,从而大幅地减少经济损失。2006年,MaviMNPV由国际热带农业研究所引入贝宁,贝宁的评估结果也证实了该病毒作为生物杀虫剂有效控制豆野螟的巨大潜力。当MaviMNPV的使用比例达到2×1013OBs/ha时,豆野螟幼虫死亡率高达88%(Srinivasanetal., 2009)。而瓜螟绒茧蜂被证实是MaviMNPV在不同地理种群中传播的途径之一(Srinivasanetal., 2009)。
4 植物源农药
植物源农药是从植物中提取的、能有效抑制农作物病虫害发生的活性成分。它们通常是植物中的多种有机物组合,对非靶标生物安全、对生态环境友好、不易产生抗药性,因此国内外学者从未停止将植物源农药应用于豆野螟生物防治上的探索。
印楝Azadiracbtaindica中提取的印楝素(Azadirachtin)是目前植物源农药产业化最为成熟的品种之一,具有较高的环境安全性。已有研究表明,在实验室条件下,印楝油及其商业化产品对豆野螟幼虫具有良好的杀虫活性(Kumaretal., 2014)。研究人员先后将印楝籽、叶的提取物,及其油、皂等多种产品应用在不同豆类作物上,豆野螟的田间防治效果各异(表4)。此外,多种豆野螟非嗜食植物提取物也对具有良好的种群抑制活性。飞机草Eupatoriumodoratum、马缨丹Lantanacamara、蟛蜞菊Sphagneticolacalendulacea乙醇提取物对豆野螟成虫和幼虫都有明显的趋避效果,蛀果率分别下降42.4%、35.4%和29.3%(曾玲等, 2001);胡椒Piperguineense和大蒜Alliumsativum的提取物能降低豆野螟卵的孵化率(Ekesi, 2000)。在田间试验中,麻疯果油(Pillaietal., 2013)、水黄皮皂(Sambathkumaretal., 2015)、木薯Manihotesculenta和面包果Artocarpusaltilis(Oparaeke, 2006)等植物提取物均能有效减少蛀果。然而,上述研究中涉及的多种(亚)热带植物对生境要求高,且现阶段其生理学研究基础较为薄弱,要实现这些植物源农药的商业化量产,仍需面临多重的挑战。
综上,印楝制剂是现阶段生物防治豆野螟的常用的植物源农药。在豆类蔬菜的早花期,印楝油与MaviMNPV配合使用的防治效果更佳(Sokameetal., 2015),是豆野螟“治花保荚”关键防治策略的重要组成部分之一。
5 总结与展望
豆野螟的生物防治技术是保障我国“菜篮子”质量安全的迫切需求。现阶段,利用性信息素进行诱捕的方法在贝宁、加纳、印度、柬埔寨等地取得了一定的成效。但是豆野螟不同地理种群(或亚种)的性信息素组成呈多态性,适用于当地的诱捕方法和经验不一定具有可复制性。在天敌昆虫方面,P.syleptae和T.javanus两种寄生蜂在贝宁和布基纳法索的成功引种和应用,为推广“以虫治虫”的豆野螟生物防治方法提供了范例。作者认为,寄生蜂资源的有效利用是从源头上治理豇豆等质量安全问题的突破口。一方面,寄生蜂能准确定位、寄生隐蔽在豇豆花苞和豆荚内的幼虫,破解了化学农药无法直接作用于虫体的难题;另一方面,寄生蜂能连续多天多次寄生,可持续抑制田间龄期大小不一的豆野螟种群,避免了在豇豆连续开花结荚期间多次用药的困境。豆野螟的昆虫病原微生物制剂和植物源农药的研发工作已积累了一定基础,尤其是在MaviMNPV和印楝制剂配合使用的方法上取得了较好的成绩。但是生物源农药的推广和应用高度依赖于“产研融合”的进程,生产企业在降低成本、提质增效、扩大产能等方面,仍有较大提升空间。
豆野螟的生物防治手段多样,但在国内的适用性仍缺乏具体实践和论证,相关应用基础研究仍处于起步阶段。我国科技人员可以从以下几方面着手,助力豆野螟绿色防控体系建设:
(一)继续研究豆野螟性信息素合成与化学感受机理。了解国内不同地理种群性信息素通讯系统的差异;研究豆野螟性信息素生物合成与化学通讯系统的基因调控机理、遗传和进化规律;进一步筛选、开发寄主植物的特异性挥发物作为引诱剂或与性信息素诱芯复配,增加田间诱捕能力。
(二)发掘我国境内豆野螟寄生性天敌昆虫资源,系统开展利用基础研究。充分发挥我国生物多样性的大国优势,明确国内豆野螟的优势寄生蜂(蝇)种类及其自然控害作用;研究寄生性天敌对豆野螟的控害机理及关键因子,解决寄生性天敌昆虫在规模化繁育、保藏与田间释放保护中的技术问题。
(三)加强昆虫病原物微生物与植物源农药新制剂的研发及其配套的使用技术。优先建立豆野螟低龄幼虫对Bt毒素的敏感基线;明确制剂与天敌昆虫、常用化学农药和其他生物源农药的相互增效或拮抗作用;解决生物农药资源在人工扩繁过程中的技术问题;研究制剂的增效途径、储存条件优化、施用后的抗逆能力等。
相信在国家政策的推动与国家自然科学基金的双重助力之下,我国生物防治技术的高质量发展势如破竹,未来可应用于豆野螟生物防治的手段也将取得创新性的突破,有助于化学农药的减量控害,为保障人民群众“舌尖上的安全”,推动“十四五”时期农业绿色转型作出重要贡献。
猜你喜欢
中老年保健(2022年7期)2022-09-20
基层中医药(2021年4期)2021-07-22
——管氏肿腿蜂的应用技术
江苏林业科技(2020年3期)2020-08-31
湖北农机化(2020年4期)2020-07-24
中国与非洲(法文版)(2019年7期)2019-07-31
现代园艺(2018年2期)2018-03-15
现代园艺(2017年21期)2018-01-03
现代农业(2016年6期)2016-02-28
上海蔬菜(2016年5期)2016-02-28
新疆农垦科技(2014年10期)2014-02-28
