
吸虱亚目非典型线粒体基因组研究进展
2023-05-27 14:00孙佳宁董文鸽
环境昆虫学报 2023年2期
孙佳宁,董文鸽
(大理大学病原与媒介研究所,云南大理 671000)
吸虱亚目Anoplura隶属于节肢动物门Arthropoda昆虫纲Insecta虱目Phthiraptera,全世界共15科50属540种(Durden &Musser, 1994),中国共11科22属96种(金大雄, 1999)。吸虱是寄生于真兽类哺乳动物体表专性寄生虫,以宿主血液为食,具有极强的宿主特异性。吸虱不仅对真兽类哺乳动物(野生兽类和家畜)产生直接危害,还在近缘动物宿主之间传播和储存人兽共患病病原体,如鼠疫、野兔热、鼠型斑疹伤寒等,在流行病学上具有保存和扩展疫源地的意义。吸虱中的体虱Pediculushumanus在全球范围内广泛分布,因体虱可直接传播战壕热(Trench fever)、回归热(Pelapsing fever)和流行性斑疹伤寒(Epidemic typhus)等人类疾病,在历史上受到高度关注(金大雄, 1999; Durden &Timm, 2001; Nafstad &Gronstol, 2001; Toshinorietal., 2002)。
随着分子生物学技术的飞速发展,目前吸虱亚目已有15种吸虱的线粒体基因组完成测序,其线粒体基因组数据均已上传至NCBI数据库(https://www.ncbi.nlm.nih.gov/),这15种吸虱的线粒体基因组均发生不同程度的裂化,其基因组结构和基因排列顺序有极强的自身特殊性,明显区别于其它节肢动物(刘静和边迅, 2021)。通常来说,动物典型的线粒体基因组结构是双链闭合环状的DNA分子,其大小为15~20 kb(Boore, 1999)。吸虱亚目昆虫线粒体基因组不再是典型的双链闭合环状DNA分子,而是裂化成数目不等的线粒体微环染色体(Minichromosomes或Minicircular mitochondrial chromosomes)。除虱目的吸虱亚目外,细角亚目(Cameronetal., 2011; Songetal., 2018)和象虱亚目(Shaoetal., 2015)中也发现线粒体基因组裂化。此外,缨翅目Thysanoptera(Dickeyetal., 2015)、啮虫目Psocoptera(Weietal., 2012; 陈世春, 2014; Fengetal., 2019)、线虫(Armstrongetal., 2000; Gibsonetal., 2007)、轮虫(Sugaetal., 2008)、中生动物(Watanabeetal., 1999)、刺胞动物(Voigtetal., 2008; Smithetal., 2011)、原生生物(Noseketal., 1998; Fanetal., 2002; Burgeretal., 2003; Marandeetal., 2005)、植物(Sugiyamaetal., 2005)以及真菌(Burger &Lang, 2003)中也发现裂化线粒体基因组。
1 吸虱亚目裂化线粒体基因组测序现状
吸虱亚目昆虫获得的首个线粒体全基因组序列是体虱(Shaoetal., 2009),自此之后陆续对更多吸虱亚目昆虫的线粒体基因组进行了测序。截止2021年8月,共获得7科7属15种吸虱亚目昆虫的线粒体基因组序列,且均已上传至GenBank数据库。迄今为止已获得虱科虱属3种吸虱(体虱、头虱P.capitis和黑猩猩虱P.schaeffi)(Shaoetal., 2009; 2012; Herdetal., 2015),阴虱科阴虱属1种吸虱(耻阴虱Pthiruspubis)(Shaoetal., 2012),血虱科血虱属3种吸虱(野猪血虱Haematopinusapri、猪血虱Ha.suis和驴血虱Ha.asini)(Jiangetal., 2013; Songetal., 2014),多板虱科多板虱属3种吸虱(亚洲多板虱Polyplaxasiatica、棘多板虱Po.spinulosa和弯多板虱Po.reclinata)(Dongetal., 2014b; 2021),甲胁虱科甲胁虱属2种吸虱(红姬甲胁虱Hoplopleuraakanezumi和克氏甲胁虱Ho.kitt)(Dongetal., 2014a),微胸虱科微胸虱属1种吸虱(羊驼虱Microthoraduspraelongiceps)(Shaoetal., 2017),猴虱科猴虱属2种吸虱(红疣猴虱Pedicinusbadii和钝猴虱Pe.obtusus)(Fuetal., 2020)(表1)。其中4个为完整的线粒体基因组,11个为部分线粒体基因组(编码区缺少部分基因或未获得全长非编码区)(图1)。
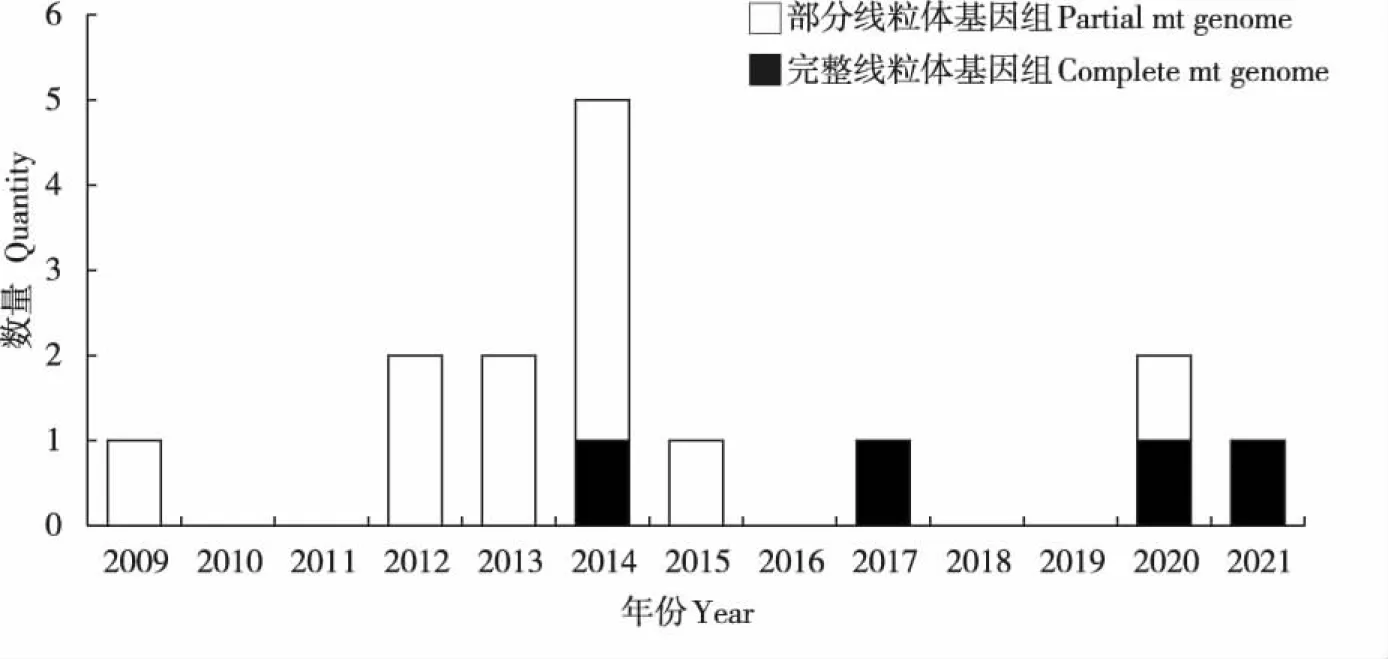
图1 近10年测序的吸虱亚目线粒体基因组数量Fig.1 Number of mitochondrial genomes Anoplura sequenced in recent 10 years
2 吸虱亚目裂化线粒体基因组基本特征
2.1 基因组结构和组成
吸虱亚目的线粒体基因组不再以典型单个环状染色体呈现,而是裂化成数目不等的线粒体微环染色体(图2),每个微环染色体由1个编码区和1个非编码区组成。吸虱线粒体基因组同样由37个基因(包括13个蛋白质编码基因、22个tRNA基因和2个rRNA基因)组成,这37个基因不均匀的分布于裂化的线粒体微环染色体上,每个微环染色体上有1~7个基因。但在部分物种中37个基因并未找全,如耻阴虱,在Shao等(Shaoetal., 2012)首次对其线粒体基因组测序时未找到其trnK、trnT和nad4基因,后续Fu等(Fuetal., 2020)再次对耻阴虱进行测序,找到了trnK和nad4基因,仍未找到trnN基因;克氏甲胁虱未找到trnH、trnF和nad5;红姬甲胁虱未找到trnF、trnG、trnH、trnL1(tag)、trnM、trnQ、nad1、nad3和nad5这9个基因(Dongetal., 2014a)。
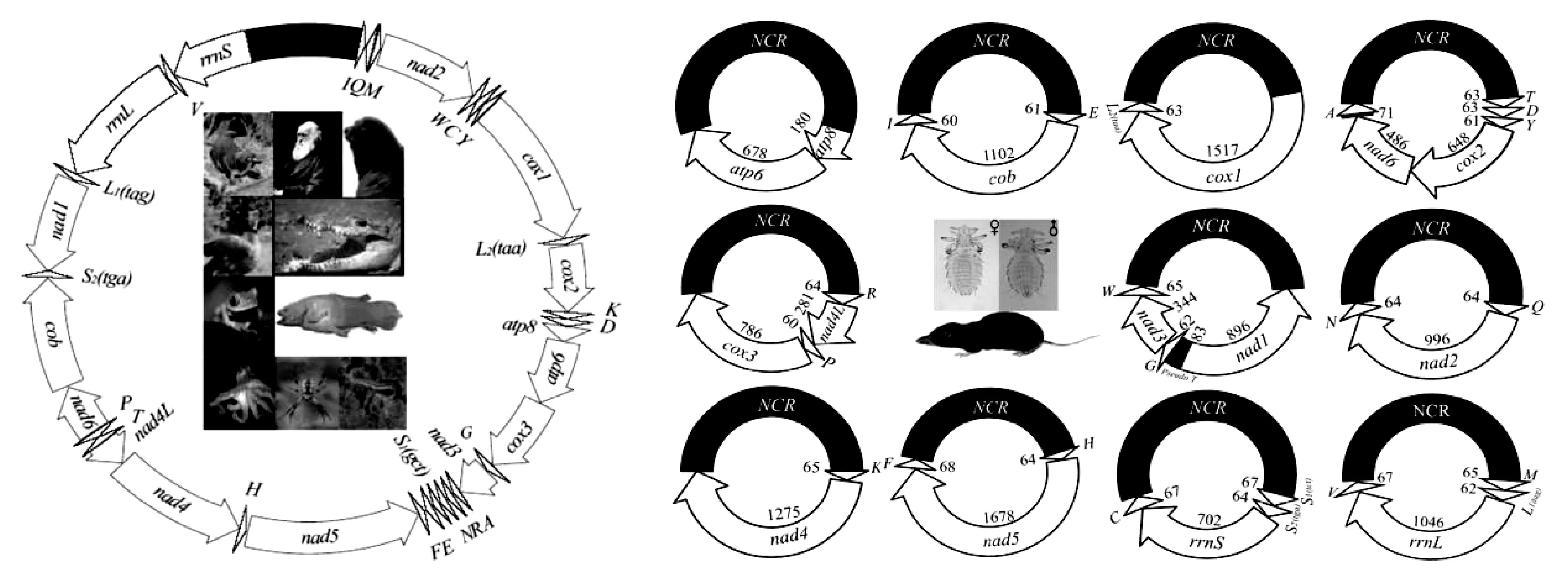
图2 大多动物典型线粒体基因组(左)和吸虱非典型线粒体基因组(右)(董文鸽, 2014; Dong et al., 2021)Fig.2 Typical mitochondrial genome of most animals (left) and atypical mitochondrial genome of sucking lice(right) (董文鸽, 2014; Song et al., 2021)
吸虱亚目线粒体基因组微环染色体的数目在9~20个不等,猪虱和驴血虱(Jiangetal., 2013; Songetal., 2014)微环染色体数目最少,仅有9个,体虱和头虱(Shaoetal., 2009; 2012)微环染色体数目最多,达20个。同属内吸虱的微环染色体数目基本一致,如多板虱属的亚洲多板虱、棘多板虱和弯多板虱均为11个微环染色体(Dongetal., 2014b; 2021);甲胁虱属的红姬甲胁虱和克氏甲胁虱均为11个微环染色体(Dongetal., 2014a);血虱属的猪虱和驴血虱均为9个微环染色体(Jiangetal., 2013; Songetal., 2014)。但也存在特殊情况,如虱属的体虱和头虱均为20个微环染色体,而黑猩猩虱只裂化成了18个微环染色体(Shaoetal., 2009; 2012; Herdetal., 2015);猴虱属的红疣猴虱和钝猴虱分别有14个和12个微环染色体(Fuetal., 2020)。由此可见,同属吸虱的线粒体基因组裂化微环染色体数目大部分一致,但也会存在差异,差异程度并不大。书虱线粒体基因组裂化成2~3个微环染色体(Weietal., 2012; 陈世春, 2014; Fengetal., 2019),茶黄硬蓟马ScirtothripsdorsailsSA1线粒体基因组裂化成1个大环和1个只含有2个基因的小环(Dickeyetal., 2015)。由此可见,吸虱亚目线粒体基因组裂化环数明显多于书虱和蓟马。这是否与吸虱吸食宿主血液的生活习性有关?需进一步验证。
2.2 RNA基因
吸虱亚目的tRNA基因大多为典型的三叶草结构,与大多数昆虫一样(Cameron, 2014),吸虱亚目的部分吸虱(如体虱、头虱、阴虱、黑猩猩虱、猪虱、猴虱、甲胁虱、亚洲多板虱和棘多板虱)的trnS1(tct)基因缺少D臂(Shaoetal., 2009; 2012; Jiangetal., 2013; Dongetal., 2014a; 2014b; Herdetal., 2015; Fuetal., 2020),但弯多板虱、驴血虱和羊驼虱的trnS1(tct)基因拥有完整的三叶草结构(Songetal., 2014; Shaoetal., 2017; Dongetal., 2021)。此外,羊驼虱的trnA和trnR基因(Shaoetal., 2017),黑猩猩虱的trnQ基因缺少DHU臂(Herdetal., 2015),红疣猴虱trnA基因缺少TΨC臂(Fuetal., 2020)。tRNA基因的氨基酸接受臂和反密码子臂的长度分别为7 bp 和5 bp,非常保守极少发生变异。而DHU臂和TΨC臂变异程度相对较大,分别为3~4 bp和2~8 bp。DHU环和TΨC环变异最大分别为3~16 bp和3~19 bp,反密码子环较为保守,除耻阴虱(Shaoetal., 2012)的trnA基因为9 bp外,其余均为7 bp。
吸虱昆虫的tRNA基因大多遵循碱基配对原则,但也存在基因错配现象,在15种吸虱tRNA基因中存在G-U、U-U、A-G、A-C、C-U、A-A、C-C、G-G错配,其中以G-U错配为主,约占全部基因错配数量的72.3%,每种吸虱的tRNA基因均存在G-U错配。G-G错配最少,占全部基因错配数量的0.6%,仅在猪虱的trnE基因以及棘多板虱的trnH和trnR基因中出现(Jiangetal., 2013)。氨基酸接受臂、DHU臂、TΨC臂和反密码子臂均存在基因错配,其中氨基酸接受臂上的基因错配最多约占39.1%,TΨC臂最少约占15.8%,这一现象与线粒体基因组未发生裂化的鳞翅目昆虫相同(王维等, 2013)。每个吸虱的tRNA基因存在碱基错配数目也不同,钝猴虱全部22个tRNA基因存在错配数目各为33个,而野猪血虱的全部tRNA基因存在错配数目,高达63个,是吸虱亚目中错配数目最多的吸虱(Jiangetal., 2013; Fuetal., 2020)。嗜虫目的嗜卷书虱Liposcelisbostrychophila21个tRNA基因存在55处错配,小眼书虱14个tRNA基因存在42处错配。细角亚目的4种住牛虱tRNA基因错配数目34~44个。较其他线粒体基因组未发生裂化的昆虫,线粒体基因组发生裂化的昆虫碱基错配数偏高(Weietal., 2012; 陈世春, 2014; Songetal., 2018; 单良陈玉, 2021)。即使同属内的吸虱,基因错配的数目差异也会较大,如血虱属的野猪虱(63个基因错配)和驴血虱(47个基因错配),相差 16个。若从吸虱宿主的角度分析,猪虱的宿主家猪和野猪均属于偶蹄目动物,而驴血虱的宿主马属于奇蹄目动物。宿主同为猪的猪血虱和野猪血虱tRNA基因错配数目仅差3个。即使存在不同数目的基因错配,但基因错配可以通过编辑矫正,并不会影响tRNA基因的转运功能(Varani &Mcclain, 2000)。
吸虱亚目的trnL1(tag)和trnL2(taa)基因存在较长的等同序列。其中耻阴虱、体虱、头虱、黑猩猩虱和钝猴虱的trnL1(tag)和trnL2(taa)基因,除的第3位反密码子外,其余碱基均相同(Shaoetal., 2012; Herdetal., 2015; Fuetal., 2020)。此外部分吸虱的存在2~3段较长的等同序列,如羊驼虱、棘多板虱和疣猴虱存在两段较长的等同序列,猪虱存在3段较长的等同序列(Shaoetal., 2017; Fuetal., 2020; Dongetal., 2021)。亚洲多板虱、弯多板虱和驴血虱均存在一段较长的等同序列分别为28、25和15 bp(Songetal., 2014; Dongetal., 2014b; 2021)。然而甲胁虱属的吸虱并没有较长的等同序列,红姬甲胁虱未能找到trnL1(tag)基因,无法进行比较。克氏甲胁虱trnL1(tag)和trnL2(taa)基因等同序列长度仅为7 bp(Dongetal., 2014a),缺少较长等同序列这一特点与芒角亚目的亚库巴果蝇Drosophilayakuba(Clary &Wolstenholme, 1985),丝角亚目的尖叫虱Bothriometopusmacrocnemis(Cameronetal., 2007)和鸽虱Campanulotesbidentatuscompar(Covacinetal., 2006)以及钝角亚目袋鼠虱Heterodoxusmacropus(Shaoetal., 2001)这些线粒体基因组未发生裂化的昆虫相似(表1)。
吸虱亚目昆虫的rRNA基因包括rrnS和rrnL基因,rRNA基因的长度都十分保守且有AT偏向性,rrnS基因平均长度为723 bp,rrnL基因平均长度为1 119 bp。所有吸虱的rRNA基因均呈AT偏向,亚洲多板虱rrnS和rrnL基因的AT含量在所有吸虱中最低,分别为55.3%和60.4%(Dongetal., 2014b);黑猩猩虱最高,分别为73.2%和72.3%(Herdetal., 2015)。其它未发生裂化的昆虫rRNA基因AT含量通常高于吸虱(Lietal., 2018; Fuetal., 2021)。虱属、血虱属和多板虱属均有3个种可在NCBI获得线粒体基因组序列,比较它们的rrnS和rrnL基因相似度发现,虱属相似度最高,分别为84.2%和86.3%;其次为血虱属,相似度分别为78.2%和77.1%;多板虱属相似度最低,分别为61.3%和68.9%。
2.3 非编码区
吸虱裂化线粒体基因组,每个微环染色体均有1个非编码区,如体虱(Shaoetal., 2009)的线粒体基因组裂化成20个微环染色体,意味着体虱的线粒体基因组同时存在20个非编码区。但驴血虱的R-nad4L-rrnS-C微环染色体除了1个较长的非编码区(1 951 bp),同时还存在1个440 bp的非编码区(Songetal., 2014)。这种情况也在弯多板虱nad1-G-nad3-W、红姬甲胁虱R-nad4L-P-cox3-A-T和克氏甲胁虱D-Y-cox2-T微环染色体中出现,除1个较长的非编码区外还分别有83、49和28 bp的非编码区(Dongetal., 2014a; 2021)。15种吸虱中仅4种吸虱获得了与其微环染色体数目对应数量的全长非编码区(Songetal., 2014; Shaoetal., 2017; Fuetal., 2020; Dongetal., 2021),部分吸虱仅获得了线粒体基因组中某些微环染色体的全长非编码区。如头虱的cox1和K-nad4微环染色体;黑猩猩虱cox1微环染色体;野猪虱的Q-nad1-T-G-nad3-W和rrnS-C微环染色体;亚洲多板虱的cox1-L2(taa)和M-L1(tag)-rrnL-V微环染色体;棘多板虱的M-L2(taa)-rrnL-V和S1(tct)-S2(tga)-rrnS-C微环染色体;红姬甲胁虱的rrnS和Y2(ata)-rrnL-V微环染色体;克氏甲胁虱的rrnS,M-L1(tag)-rrnL-V和I-cox1微环染色体(Shaoetal., 2012; Jiangetal., 2013; Dongetal., 2014a; 2014b)。耻阴虱全部10个微环染色体均获得了全长非编码区,家猪虱线粒体基因组9个微环染色体中的8个微环染色体均获得全长非编码区,而人体虱和红疣猴虱每个微环染色体仅获得了部分非编码区,未能获得全长非编码区(Shaoetal., 2009; 2012; Jiangetal., 2013; Herdetal., 2015; Fuetal., 2020)。
在同一物种不同微环染色体上的全长非编码区均有较高的相似性,首个获取全部微环染色体全长非编码区的驴血虱(Songetal., 2014),9个非编码区之间的相似性达96%。亚洲多板虱的2个全长非编码区之间的相似性达96%,棘多板虱的2个全长非编码区之间的相似性也高达98%(Dongetal., 2014b)。红姬甲胁虱的2个全长非编码区之间的相似性最低,仅78.7%,克氏甲胁虱的3个全长非编码区之间相似性在87%~93%之间(Dongetal., 2014a)。书虱线粒体基因组由2~3个微环染色体组成,与吸虱不同,书虱的每个环中含有较多的非编码区,如嗜虫书虱Liposcelisentomophila的2个环中共有38处非编码区,其中3个非编码区存在相似序列,故将其认为控制区(陈世春, 2014)。象虱Haematomyzuselephantis获得全长的4个微环染色体的非编码区之间也有高度相似(>97%)(Shaoetal., 2015)。
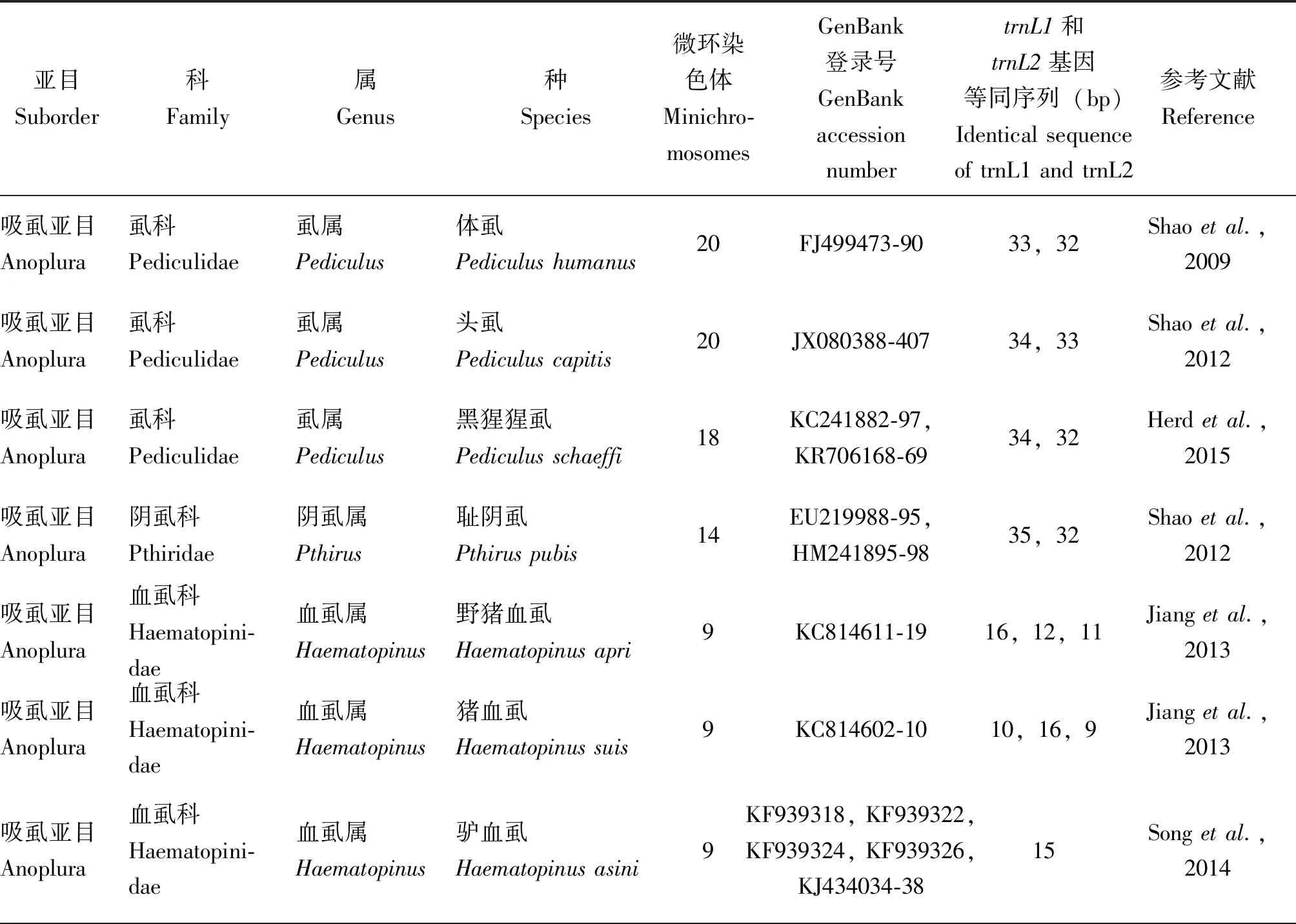
表1 寄生虱和果蝇trnL1(tag)和trnL2(taa)基因等同序列的长度
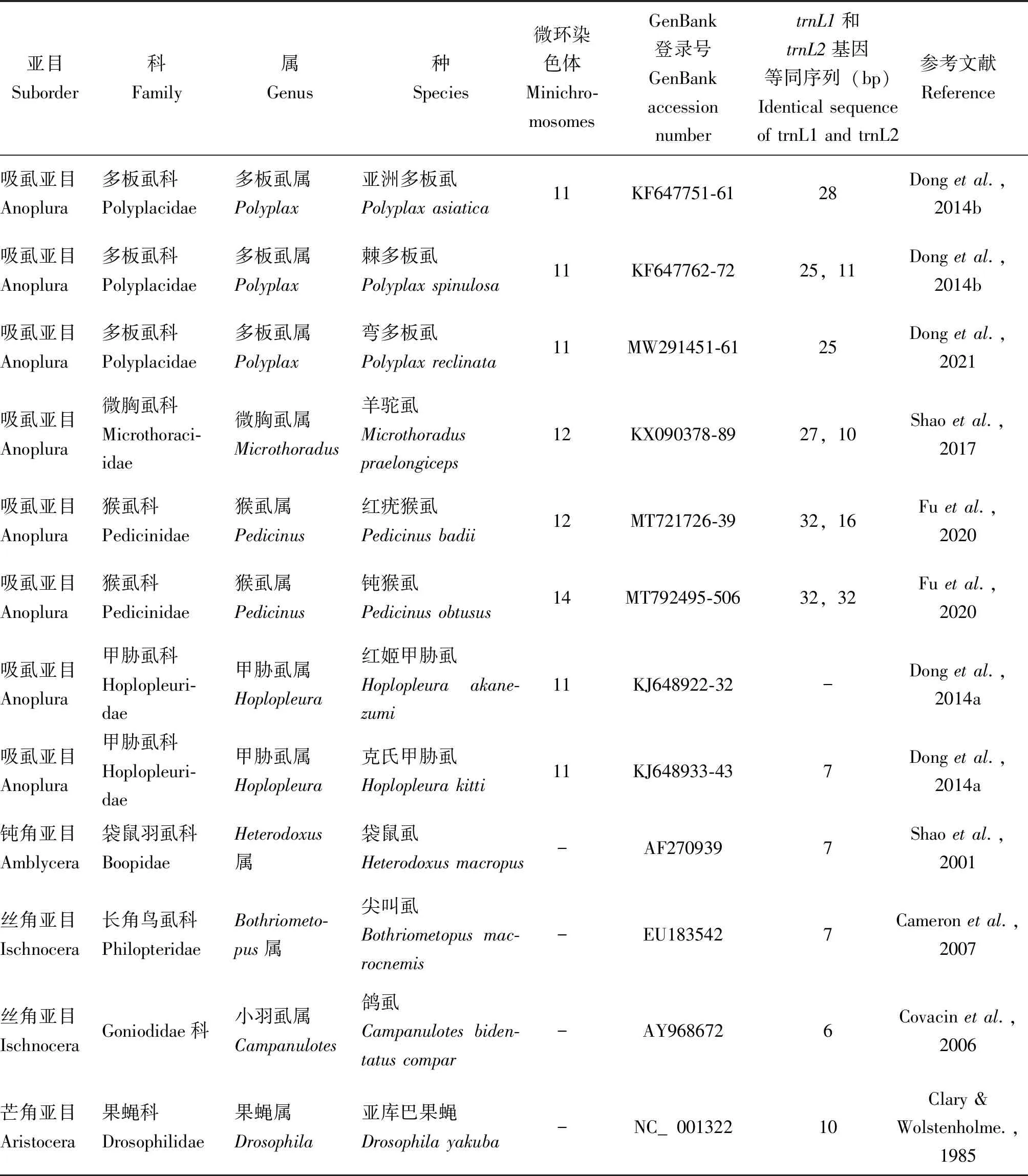
续表1 Continued table 1
在吸虱的每个微环染色体的编码区上游和下游的两端,均会存在一段较为保守的序列。在编码区5′端上游的非编码区存在一段长度45~203 bp的 AT富集区(AT-rich motif),AT含量为59%~100%。编码区3′端下游的非编码区存在一段长度33~90 bp的GC富集区(GC-rich motif),GC含量为56.7%~82%。由于这两个保守序列在非编码区的两端,非编码区又称为控制区,负责线粒体基因组的复制和转录过程(Taanman, 1999)。Shao等人认为,微环染色体的转录可能开始于AT富集区,终止于GC富集区(Shaoetal., 2017)。
吸虱全长非编码区在955~3 265 bp不等,其中驴血虱的每个微环上的非编码区长度均在2 kb以上,相较于其它吸虱,是非编码区最长的物种。即使在同一物种内,不同微环染色体上的非编码区长度也有差异,如羊驼虱最长非编码区和最短非编码区相差了450 bp,而阴虱最长非编码区和最短非编码区之间相差 627 bp(Shaoetal., 2012)。与大多数昆虫一样,这些非编码区长度差异大且存在重复次数不同的串联重复序列,或不存在串联重复序列;或是在不同微环染色体的非编码区中插入或删除不同数目的碱基序列。非编码区的进化速率明显快于编码区,具有许多不保守的特征,如串联重复序列、潜在的发夹结构等(Jiangetal., 2013; Dongetal., 2014b; Songetal., 2014; 李爱玲等, 2014; Shaoetal., 2017)。
3 祖先线粒体核型推测
3.1 概念
人们通常将亚库巴果蝇的线粒体基因组的排列方式作为昆虫的线粒体基因组最原始的排列方式(魏书军和陈学新, 2011),而吸虱和象虱的线粒体基因组均发生裂化,明显区别于大多数昆虫的线粒体基因组。为了更加方便、准确地描述它们的线粒体基因组,Shao等首次提出“线粒体核型(Mitochondrial karyotype)”这一概念,来描述像吸虱以及象虱由多个微环染色体组成的线粒体基因组(Shaoetal., 2015)。吸虱亚目线粒体核型指:1)微环染色体的数量;2)微环染色体的拓扑结构是线型还是环型;3)每个微环染色体的基因含量和基因排列。
3.2 推测方法
吸虱亚目的昆虫线粒体基因组均已裂化成不同数目的微环染色体,其特殊的线粒体基因组结构导致没有合适的进化模型用于分析并重建吸虱祖先线粒体核型。Shao等根据6科6属12种吸虱和象虱构建的系统发育树与Light等根据cox1、EF-1α和18S基因,对8科17属49种吸虱分别构建系统发育树以及Kim根据39种形态特征对 15科吸虱构建的系统发育树的结果一致(Kim, 1988; Lightetal., 2010; Shaoetal., 2017)。吸虱亚目的吸虱在大约750万年前分化成两个主要的进化支:一大进化支为棘虱科Echinophthiridae,鄂虱科Linognathidae,甲胁虱科的甲胁虱属和Pterophthirus属,多板虱科的拟鄂虱属Linognathides、新血虱属Neohaematopinus和树鼩虱属Sathrax;另一大进化支为血虱科,多板虱科的多板虱属和Lemurpediculus属,甲胁虱科的钩板虱属Ancistroplax,虱科,阴虱科和猴虱科。Shao等构建的系统发育树的基础上,利用简约法(Parsimony method)(Gordonetal., 2009; Shaoetal., 2017)初步推测吸虱祖先线粒体核型。后续Fu等(2020)和Dong等(2021)利用同样的方法分别对寄生于灵长类动物体表的吸虱(虱属,阴虱属,猴虱属)和多板虱属吸虱的祖先线粒体核型进行了推测。
若某一基因特征为吸虱祖先线粒体核型需满足:1)其存在于吸虱两大进化支中的至少1个进化支,且存在于外群象虱中;或2)其同时存在于吸虱的两大进化支中。若某一基因特征为寄生于灵长类动物体表的吸虱祖先线粒体核型需满足:1)其同时存在于虱属、阴虱属和猴虱属;或2)虱属物种以及虱属物种或阴虱属物种共有;或3)3个属中的1个或多个物种以及其他寄生虱。若某一基因特征为多板虱属吸虱的祖先线粒体核型:1)其存在于至少1种多板虱属吸虱中且存在于1种或多种非多板虱属吸虱中;或2)同时存在于全部的3种多板虱(亚洲多板虱、棘多板虱和弯多板虱)中。
3.3 推测结果
最终Shao等(2017)推测吸虱祖先线粒体核型:1)由11个不同的微环染色体组成;2)每个微环染色体由1个编码区和1个非编码区构成,且编码区包含1~6个基因(图3-A)。Fu等(2020)根据体虱、头虱、阴虱、黑猩猩虱、钝猴虱、红疣猴虱的线粒体基因组对寄生在灵长类动物体表寄生虱的祖先线粒体核型进行了初步推测,但这6种吸虱的线粒体基因组数据量用于推测祖先线粒体核型并不充足,且tRNA基因的移动性较强,因此仅推测出蛋白质编码基因和rRNA基因在祖先核型中的排列方式:1)由12个不同的微环染色体组成;2)每个微环染色体由1个编码区和1个非编码区构成;3)每个微环染色体包含1~2个蛋白质编码基因或rRNA基因(图3-B)。Dong等(2021)根据多板虱属的亚洲多板虱、棘多板虱和弯多板虱,并结合其余12种吸虱和象虱对多板虱属吸虱的祖先线粒体核型进行推测:1)由11个微环染色体组成;2)每个微环染色体由1个编码区和1个非编码区构成,且编码区包含2~7个基因;3)其中7个微环染色体仅有1个蛋白质编码基因或rRNA基因;4)trnS1(tct)-trnS2(tga)可能在rrnS基因的上游或nad1基因的上游,位置尚不能确定(图3-C)。
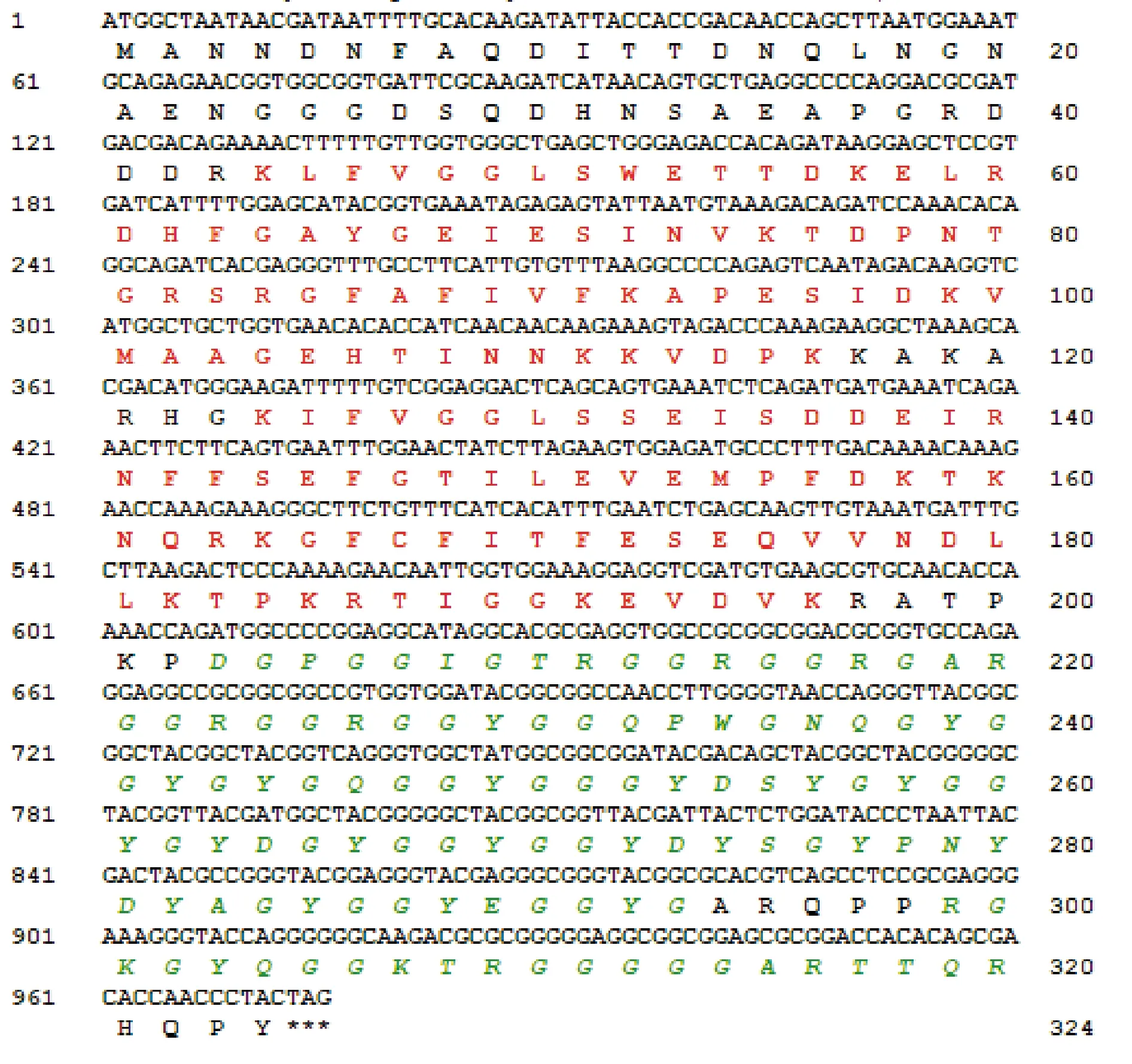
图3 吸虱亚目祖先线粒体核型推测Fig.3 Inferred ancestral mitochondrial karyotype of Anoplura
在不考虑tRNA基因的情况下,对比3个祖先线粒体核型发现,每个微环染色体均由1个编码区和1个非编码区构成。蛋白质编码基因和rRNA基因的位置基本一致,分布于10个微环线粒体上。其中7个微环线粒体染色体有1个蛋白质编码基因或rRNA基因,剩余3个微环染色体均有2个蛋白质编码基因:atp8-atp6,nad4L-cox3,nad1-nad3。但cox2和nad6基因在3个祖先线粒体核型中存在差异,吸虱祖先线粒体核型和多板虱属祖先线粒体核型一致,cox2和nad6基因在同一微环染色体中,而在寄生在灵长类动物体表的吸虱祖先线粒体核型中,这两个基因各自独立存在于不同的微环染色体中。
由于获得吸虱的线粒体基因组数量较少,对于祖先线粒体核型的推测结果尚不够精确,寄生于灵长类动物体表吸虱的祖先线粒体核型由于数据量不足且tRNA基因的较强活动性,只能初步推测出蛋白质编码基因和rRNA基因的位置。
4 展望
随着测序技术不断地更新换代,获得的吸虱亚目线粒体基因组的数量逐渐增多。目前已经对7科 7属15种吸虱的线粒体基因组进行了测序分析,但相对于全世界吸虱亚目15科50属540种(Durden &Musser, 1994)来说,仅对全部吸虱亚目物种的3%完成了线粒体基因组的测序。虽然目前对15科中的7科进行了测序,但从属这一分类阶元来看,较为单一,每科仅有1个属,且每个属仅有1~3种。这7科中的多板虱科物种最为丰富,有189种,但仅4种多板虱获得了线粒体全基因组;甲胁虱科共有157种,目前仅对2种甲胁虱的线粒体基因组进行了测序,而且这2种甲胁虱的37个基因均未能找全,红姬甲胁虱未能找到9个基因(nad1、nad3、nad5、trnF、trnG、trnH、trnL1(tag)、trnM和trnQ),克氏甲胁虱未能找到3个基因(nad5、trnF和trnH)(Dongetal., 2014a)。基因未能找全这一现象同样在耻阴虱中出现,Shao等2012年对耻阴虱线粒体基因组首次进行测序,未能找到nad4,trnK和trnN3个基因。Shao推测,未能找到的原因可能有两个:一是这3个基因所在的微环染色体要比耻阴虱其它微环染色体的丰度低;二是用于扩增耻阴虱线粒体基因组的引物不够保守。Shao更倾向于第2个原因,引物不保守这一原因也是两种甲胁虱未能找全基因推测的原因(Shaoetal., 2012)。今后可尝试设计更为保守的引物,增加试验虫体的数量以找到未能发现的基因。
每种吸虱的线粒体基因组均发生不同程度的裂化,基因不均匀的分布在每个微环染色体上,几乎每个微环染色体上至少有1个蛋白质编码基因或者rRNA基因,tRNA基因随机分布在每个微环染色体上。也有微环染色体仅包含1个基因,蛋白质编码基因、rRNA基因或tRNA基因。吸虱亚目的昆虫tRNA基因大部分为典型的三叶草结构,与大多数昆虫一样,trnS1(tct)基因常缺少D臂。tRNA基因存在基因错配现象主要为G-U错配,相比于其他线粒体基因组未发生裂化的昆虫,每个吸虱物种的tRNA基因错配数量要更多。吸虱的trnL1(tag)和trnL2(taa)基因存在较长的等同序列,这是明显区别于其他单个环状线粒体基因组的昆虫的特点,但甲胁虱属的物种缺失较长的等同序列(Dongetal., 2014a),甲胁虱这一特点是否为甲胁虱属固有特征,缺少等同序列是否意味着甲胁虱属吸虱为吸虱亚目进化较慢的物种?需对更多甲胁虱属物种进行测序,以进一步验证。此外,每个微环染色体上均有一段非编码区,目前虽获得了16种吸虱的线粒体基因组,但仅4种吸虱扩增出全部全长非编码区,大多吸虱只能获得编码区两端部分非编码区,或1~3个微环染色体的全长非编码区。由于非编码区的AT含量较高,设计的引物可能有较大局限性,导致非编码区扩增较困难(李志勇等, 2009; 阿尔祖古丽·买买提吐尔逊等, 2021)。
吸虱特殊的裂化线粒体基因组结构,吸虱不能再以亚库巴果蝇的基因排列作为假设祖先基因排列方式,2017年Shao等首次基于吸虱和象虱的8个蛋白质编码基因(atp6、atp8、cox1、cox2、cox3、cob、nad4L和nad6)和2个rRNA基因(rrnS和rrnL)构建的系统发育树,通过简约法初步推测了吸虱祖先线粒体核型(Shaoetal., 2017)。后续Fu等和Dong等陆续对寄生于灵长类动物的吸虱和多板虱属吸虱的祖先线粒体核型进行了推测(Fuetal., 2020; Dongetal., 2021)。由于获得线粒体基因组的物种过少,且tRNA基因移动性较强,寄生于灵长类动物的吸虱仅能推测出蛋白质编码基因和rRNA基因在祖先线粒体核型中的位置。同样,多板虱属的trnT基因的具体位置也尚不能准确确定。若想进一步验证推测祖先核型的准确性,或推测出tRNA基因的具体位置,需要对更多的吸虱线粒体基因组进行测序,以完善数据。
本文综述了当前已测序的15种吸虱线粒体基因组的结构和组成、RNA基因、非编码区以及对吸虱祖先线粒体核型推测的方法和结果。这15种吸虱均有裂化线粒体基因组,其微环染色体的数量、以及每个微环染色体的基因结构和基因顺序彼此不同,甚至在同属中亲缘关系较近的物种间也存在差异,这与大多数动物极其保守的单染色体线粒体基因组形成鲜明对比。这很好地弥补了动物线粒体基因组传统认识的不足和局限,对理解线粒体的起源和进化以及物种形成和进化都具有重要意义。
猜你喜欢
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
宁夏医学杂志(2020年3期)2021-01-21
科学之谜(2019年3期)2019-03-28
科学之谜(2018年8期)2018-09-29
恋爱婚姻家庭·养生版(2016年9期)2016-09-07
哈尔滨医药(2015年2期)2015-12-01
中央民族大学学报(自然科学版)(2015年2期)2015-06-09
中国当代医药(2015年9期)2015-03-01
应用海洋学学报(2014年3期)2014-11-22
