
外源钙和水杨酸对弱光下观光木幼苗生理特性的影响
2023-05-22 09:11覃永华刘世男
北华大学学报(自然科学版) 2023年3期
覃永华,李 婷,杨 梅,刘世男
(1.广西壮族自治区林业勘测设计院,广西 南宁 530011;2.广西壮族自治区南宁树木园,广西 南宁 530031;3.广西大学林学院,广西 南宁 530004)
光是影响植物生长发育和生理功能的重要环境因子之一[1].在遮阴状态下,植物的可溶性蛋白、SOD、POD等显著降低,如油茶(Camelliaoleifera)、望天树(Parashoreachinensis)和野鸭椿(Euscaphisjaponica(Thunb.) Kanitz)等[2-5],而MDA和游离脯氨酸含量显著升高,如红芸豆(Phaseolusvulgaris)等[6];此外,植物的叶绿素荧光Fo、Fm、Fv/Fm也呈现不同程度的降低,如辣椒(Capsicumannuum)、兴安落叶松(LarixdahuricaTurcz.ex Trautv.)等[7-8].可见,弱光胁迫使植物生理功能受到影响,进而影响植物生长发育.研究表明,喷施激素、化学药剂等外源物质,能缓解植物在胁迫条件下的伤害.目前,关于水杨酸(SA)和氯化钙(CaCl2)增强植物抗逆性的作用已有很多报道[9-10].比如,有研究显示,SA可以缓解菊芋(Helianthustuberosus)所受的铝胁迫[11],还可以增强孔雀草(Tagetespatula)在镉胁迫下的抗性[12];CaCl2可以缓解绦柳(Salixmatsudana)幼苗盐胁迫[13],对元宝枫(Acertruncatum)高温胁迫也有缓解效应[14].此外,外源CaCl2和SA能够提高黄瓜(Cucumissativus)、番茄(Solanumlycopersicum)、甜瓜(Cucumismelo)对弱光的耐受性,降低弱光胁迫造成的伤害[15-18].
观光木(Tsoongiodendronodorum)为木兰科(Magnoliaceae)观光木属(Tsoongiodendron)常绿乔木,兼具用材、观赏和药用价值[19-20].由于砍伐过多及环境破坏,观光木的野外数量很少,已被列为国家珍稀濒危二级保护植物[21].目前,观光木研究主要集中在苗木培育、经营、抗逆性等方面[21-24].已有研究[25]表明,弱光会对观光木生长、生理造成显著影响,但关于观光木在弱光胁迫下的缓解方法还少有报道.本文旨在探索施加不同浓度CaCl2和SA对观光木缓解弱光胁迫的作用,研究在弱光胁迫下,施加外源CaCl2和SA对观光木抗氧化酶活性及叶绿素荧光参数的影响,确定观光木缓解弱光胁迫最适宜的CaCl2和SA浓度,为开展观光木在弱光逆境下的生长研究提供科学依据.
1 材料与方法
1.1 供试材料
试验材料为南宁树木园培育的6个月大观光木实生苗,选取生长状况良好,高度、地径一致的苗木.
1.2 试验设计
本试验在人工光照培养箱中进行.环境条件:光周期14 h/10 h,光照强度30 000 lx,温度25 ℃,相对湿度70%.适应3 d,每天同一时间分别喷施不同浓度CaCl2(0、5、10、20、50 mmol/L)和SA(0、1、2、5、10 mmol/L),同时以喷水为对照,每天1次,喷至叶面滴水,连喷3 d.然后将一部分苗木进行弱光处理,光照强度设置为6 000 lx,其他条件不变.每个处理3个重复,每个重复为8株苗,处理21 d后测定各项指标.
1.3 生理指标测定
丙二醛(MDA)含量采用硫代巴比妥酸法测定[26],超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)光还原法测定[27],过氧化物酶(POD)活性采用愈创木酚法测定[14],多酚氧化酶(PPO)活性采用邻苯二酚法测定[28],抗坏血酸过氧化物酶(APX)活性采用紫外吸收法测定[29],可溶性蛋白质含量采用考马斯亮蓝染色法测定[30],游离脯氨酸含量采用茚三酮法测定[30].
1.4 叶绿素荧光测定
采用便携式叶绿素荧光仪PAM-2500测定叶片初始荧光(Fo)和最大荧光(Fm)等参数,整理、计算得到光系统Ⅱ(PSⅡ)最大光化学效率(Fv/Fm)、非光化学淬灭系数(NPQ)等参数.
1.5 数据处理
采用Microsoft Excel 2016对数据进行整理并制图,利用SPSS 26.0对数据进行差异性及显著性检验.
2 结果与分析
2.1 不同浓度CaCl2和SA对弱光下观光木幼苗生理指标的影响
2.1.1 SOD
如图1 a所示,与正常光照相比,弱光条件下观光木幼苗叶片的SOD活性显著降低(P<0.05).对弱光下幼苗进行CaCl2处理SOD活性总体上升,随着CaCl2浓度的提高呈先升高后降低趋势.其中,10、20 mmol/L CaCl2处理SOD活性显著高于其他浓度处理,与弱光处理(即0 mmol/L CaCl2)相比显著上升(P<0.05),但与正常光照处理相比差异不显著;5、50 mmol/L CaCl2处理与弱光处理差异不显著.
对弱光下幼苗进行SA处理的SOD活性总体上升,随着SA浓度的提高呈先升高后降低的趋势.其中,SOD活性在2 mmol/L SA处理最高,10 mmol/L SA处理最低;2 mmol/L SA处理与其他SA浓度处理差异显著(P<0.05),但与正常光照处理差异不显著.
2.1.2 POD
如图1 b所示,与正常光照相比,弱光条件下观光木幼苗叶片的POD活性显著降低(P<0.05).对弱光下幼苗进行CaCl2处理的POD活性总体上升,随着CaCl2浓度的提高呈先上升后下降趋势,但不同CaCl2处理间差异不显著,与弱光处理、正常光照处理差异显著(P<0.05).
弱光下幼苗进行SA处理的POD活性总体上升,随着SA浓度的提高呈先升高后降低的趋势.其中,POD活性在2 mmol/L SA处理最高,10 mmol/L SA处理最低;2 mmol/L SA处理与其他SA浓度处理差异显著(P<0.05).
2.1.3 PPO
如图1 c所示,与正常光照相比,弱光下观光木幼苗叶片的总PPO活性显著降低(P<0.05).对弱光下幼苗进行CaCl2处理的PPO活性显著上升,但不同浓度处理间差异不显著.
对弱光下幼苗进行SA处理的PPO活性总体上升,且随着SA浓度的提高呈先上升后下降趋势.其中,PPO活性在2 mmol/L SA处理最高,10 mmol/L SA处理最低;2 mmol/L SA处理与其他SA浓度处理差异显著(P<0.05).
2.1.4 APX
如图1 d所示,与正常光照相比,弱光条件下观光木幼苗叶片的总APX活性显著降低(P<0.05).对弱光下幼苗进行CaCl2处理的APX活性显著上升,随着CaCl2浓度的提高呈先上升后下降趋势,不同浓度处理间差异显著(P<0.05).其中,10 mmol/L CaCl2处理的APX活性最高,而50 mmol/L处理最低,但仍显著高于弱光处理.
对弱光下幼苗进行SA处理的APX活性显著上升,随着SA浓度的提高呈先上升后下降趋势.不同浓度处理间差异显著(P<0.05),其中,APX活性在5 mmol/L SA处理最高,10 mmol/L SA处理最低.

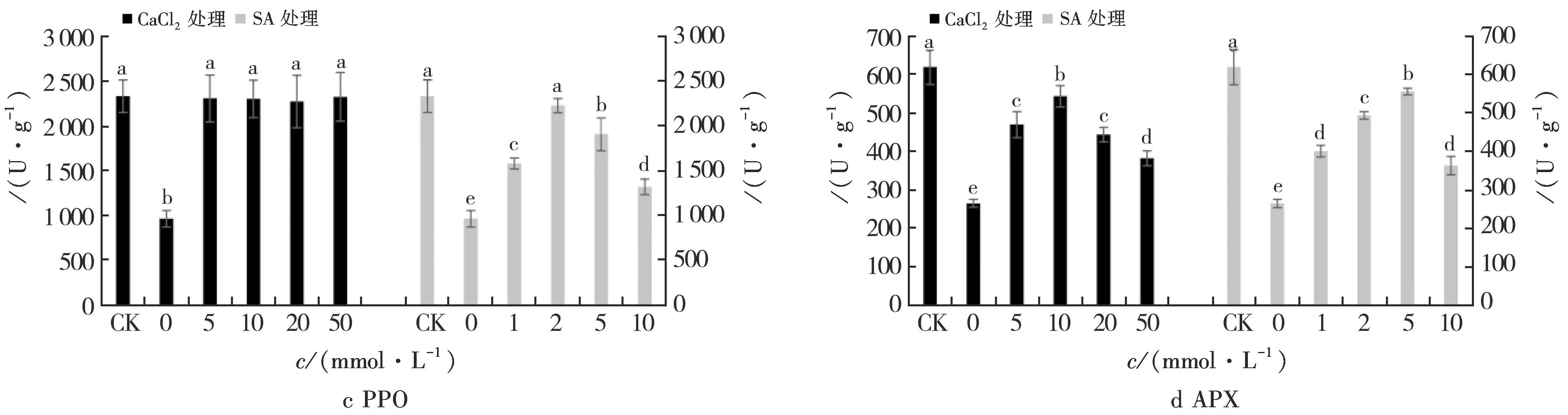
不同小写字母表示差异显著(P<0.05);相同小写字母表示差异不显著(P>0.05).图1 不同浓度CaCl2和SA对弱光下观光木幼苗叶片抗氧化酶活性的影响Fig.1 Effect of different concentrations of CaCl2 and SA on the antioxidant enzyme activities of Tsoongiodendron odorum seedlings under weak light
由2.1.1~2.1.4结果可以看出:弱光使观光木幼苗叶片中的SOD、POD、PPO、APX活性显著降低,喷施一定浓度CaCl2和SA后上述酶活性均提高.其中,10 mmol/L和20 mmol/L CaCl2处理下SOD活性与正常光照差异不显著,所有CaCl2处理下PPO活性均与正常光照时差异不显著,同时,10 mmol/L CaCl2处理POD和APX活性虽然低于正常光照处理,但高于其他CaCl2处理;此外,2 mmol/LSA处理SOD、POD、PPO活性与正常光照处理差异不显著.由此可见,喷施10 mmol/L CaCl2和2 mmol/L SA能有效缓解光照不足给观光木幼苗抗氧化酶带来的伤害.
2.1.5 MDA
如图2 a所示,与正常光照相比,弱光条件下观光木幼苗叶片的总MDA含量显著上升(P<0.05).对弱光下幼苗进行CaCl2处理的MDA活性显著上升,随着CaCl2浓度的提高呈先下降后上升趋势.其中,10 mmol/L CaCl2处理MDA含量最低,显著低于其他浓度处理和正常光照处理(P<0.05).
对弱光下幼苗进行SA处理的MDA含量显著下降(P<0.05),随着SA浓度的提高呈先下降后上升趋势.其中,2 mmol/L SA处理MDA含量最低,与5 mmol/L和10 mmol/L SA处理差异显著(P<0.05),但与正常光照和1 mmol/L SA处理差异不显著.
2.1.6 可溶性蛋白
如图2 b所示,与正常光照相比,弱光条件下观光木幼苗叶片的可溶性蛋白含量显著降低(P<0.05).对弱光下幼苗进行CaCl2处理的可溶性蛋白含量总体上升,随着CaCl2浓度的提高呈先上升后下降趋势.其中,10 mmol/L CaCl2处理的可溶性蛋白含量最高,显著高于其他浓度处理和弱光处理,与正常光照处理差异不显著.
对弱光下幼苗进行SA处理的可溶性蛋白含量也总体上升,随着SA浓度的提高呈先升高后下降趋势.其中,2 mmol/L SA处理的可溶性蛋白含量最高,与除5 mmol/L SA以外的处理差异显著(P<0.05).
2.1.7 游离脯氨酸
如图2 c所示,与正常光照相比,弱光条件下观光木幼苗叶片的游离脯氨酸含量显著升高(P<0.05).对弱光下幼苗进行CaCl2处理的可溶性蛋白含量总体下降,随着CaCl2浓度的提高呈先下降后上升趋势.其中,10 mmol/L CaCl2处理游离脯氨酸含量最低,仅与20 mmol/L CaCl2和正常光照处理差异不显著.
对弱光下幼苗进行SA处理的游离脯氨酸含量总体变化不大,其中,2 mmol/L SA处理下游离脯氨酸含量显著低于其他浓度处理(P<0.05),但仍显著高于正常光照处理(P<0.05).


不同小写字母表示差异显著(P<0.05);相同小写字母表示差异不显著(P<0.05).图2 不同浓度CaCl2和SA对弱光下观光木幼苗MDA、可溶性蛋白和游离脯氨酸的影响Fig.2 Effect of different concentrations of CaCl2 and SA on the MDA,soluble protein and activities of Tsoongiodendron odorum seedlings under weak light
由2.1.5~2.1.7可见,弱光条件使得观光木幼苗叶片中的MDA、游离脯氨酸含量显著升高,而可溶性蛋白含量显著降低,喷施一定浓度的CaCl2和SA后,MDA、游离脯氨酸含量显著下降,而可溶性蛋白含量显著上升.其中,10 mmol/L CaCl2处理的MDA、游离脯氨酸和可溶性蛋白含量与正常光照处理差异不显著;2 mmol/L SA处理的MDA含量与正常光照处理差异不显著,同时,2 mmol/L SA处理的可溶性蛋白含量低于正常处理,但仍显著高于其他浓度SA处理,而在相同处理下游离脯氨酸含量表现出相反的规律.可见,喷施10 mmol/L CaCl2和2 mmol/L SA能有效缓解光照不足对观光木幼苗MDA、可溶性蛋白、游离酸脯氨酸含量的胁迫影响.
2.2 不同浓度CaCl2和SA对弱光下观光木幼苗叶绿素荧光参数的影响
2.2.1 最大荧光Fm
如图3 a所示,与正常光照相比,弱光条件下观光木幼苗Fm显著降低(P<0.05).对弱光下幼苗进行CaCl2处理的Fm总体上升,随着CaCl2浓度的提高呈先升高后下降趋势.其中,10 mmol/L CaCl2处理Fm最高,仅与弱光和正常处理差异显著(P<0.05).
对弱光下幼苗进行SA处理的Fm总体上升,随着SA浓度的提高呈先升高后下降趋势.其中,2 mmol/L SA处理Fm最高,与弱光(即0 mmol/L SA)和5 mmol/L SA处理差异显著(P<0.05),但与1、5 mmol/L SA和正常光照处理差异不显著.
2.2.2 初始荧光Fo
如图3 b所示,与正常光照相比,弱光下观光木幼苗Fo显著下降(P<0.05).对弱光下幼苗进行CaCl2处理的Fo总体上升,随着CaCl2浓度提高而下降.其中,10 mmol/L CaCl2处理Fo最高,且显著高于与弱光和50 mmol/L CaCl2处理,与其他处理差异不显著.
对弱光下幼苗进行SA处理的Fo总体上升,随着SA浓度的提高呈先升高后下降趋势.其中,2 mmol/L SA处理Fo最高,仅与弱光和5 mmol/L SA处理差异显著(P<0.05).
2.2.3 最大光化学效率Fv/Fm
如图3 c所示,与正常光照相比,弱光下观光木幼苗Fv/Fm显著下降(P<0.05).对弱光下幼苗进行CaCl2处理的Fv/Fm总体上升,随着CaCl2浓度的提高呈先升高后下降趋势.其中,10 mmol/L CaCl2处理Fv/Fm最高,且显著高于弱光处理,与其他处理差异不显著.
对弱光下幼苗进行SA处理的Fv/Fm总体上升,随着SA浓度的提高呈先升高后下降趋势.其中,5 mmol/L SA处理Fv/Fm最高,与2 mmol/L SA处理差异不显著,但与其他处理差异显著(P<0.05).
2.2.4 非光化学淬灭系数NPQ
如图3 d所示,与正常光照相比,弱光下观光木幼苗NPQ显著升高(P<0.05).对弱光下幼苗进行CaCl2处理的NPQ总体下降,随着CaCl2浓度的提高呈先下降后上升趋势.其中,10 mmol/L CaCl2处理NPQ最低,但与其他处理差异不显著.
对弱光下幼苗进行SA处理的NPQ总体下降,其中,2 mmol/L SA处理NPQ最低,仅与弱光(0 mmol/L SA)和正常光照处理差异显著(P<0.05).
由2.2.1~2.2.4可见,弱光条件使得观光木幼苗叶片中的Fm、Fo和Fv/Fm显著降低而NPQ显著升高,喷施一定浓度的CaCl2和SA后,Fm、Fo、Fv/Fm显著上升,而NPQ显著下降.其中,10 mmol/L CaCl2处理Fm、Fo、Fv/Fm与NPQ与正常光照处理差异不显著;此外,2 mmol/L SA处理Fm、Fo含量与正常光照处理差异不显著.可见,喷施10 mmol/L CaCl2和2 mmol/L SA能有效缓解光照不足对观光木幼苗叶片叶绿素荧光参数造成的不良影响.


不同小写字母表示差异显著(P<0.05);相同小写字母表示差异不显著(P>0.05).图3 不同浓度CaCl2和SA对弱光下观光木幼苗叶绿素荧光参数的影响Fig.3 Effect of different concentrations of CaCl2 and SA on the chlorophyll fluorescence characteristics of Tsoongiodendron odorum seedlings under weak light
3 讨论与结论
植物在受到逆境胁迫后,体内会产生大量活性氧,耐受型物种可诱导相关氧化酶活性提高,以减轻胁迫伤害,而不耐受型物种中的保护系统遭到破坏[31],MDA含量积累,对生长造成伤害[31-32].本试验中,在弱光下,观光木幼苗叶片中的SOD、POD、PPO、APX活性和可溶性蛋白含量均明显降低,而MDA和游离脯氨酸含量显著增加,说明观光木受到弱光胁迫,抗性低.然而,在弱光下施加CaCl2和SA的观光木幼苗中SOD、POD、PPO、APX活性和可溶性蛋白含量升高,MDA和游离脯氨酸含量显著降低.可见,施加CaCl2和SA后可以缓解弱光对观光木造成的伤害,这与蒋晓婷[33]、李天来等[34]的研究结果一致.
Fv/Fm降低意味着植物光合作用被抑制,有可能是受到了逆境的伤害[35];NPQ的变化反映了非光化学耗散的能量,是植物光合机构的自我保护机制[36].本试验中,在弱光下,观光木幼苗叶片的Fv/Fm、Fm和Fo降低,而NPQ升高,表明观光木受到弱光伤害且叶片叶绿素荧光特性参数发生异常,这与刘凯歌等[37]和常静等[38]的研究结果一致.施加CaCl2和SA可使弱光下观光木幼苗叶片的Fv/Fm、Fm、Fo上升,而NPQ下降,表明CaCl2和SA都可以提高观光木在弱光下的抗逆性.
此外,在弱光下用CaCl2和SA处理观光木幼苗,其POD、SOD、APX活性,可溶性蛋白含量,Fv/Fm,Fo和Fm均随浓度的提高先升高再降低,而MDA、游离脯氨酸含量和NPQ随浓度提高先下降再升高,其中,PPO、POD、SOD、APX活性和可溶性蛋白含量随着浓度提高先增加再降低,MDA和游离脯氨酸含量随浓度提高先下降再升高.由此表明:适宜浓度的CaCl2和SA处理才可以有效缓解弱光胁迫,过高或过低浓度下缓解作用都会下降.
综上所述,弱光对观光木幼苗的抗氧化酶、叶绿素荧光等生理功能造成伤害,而喷施10 mmol/L CaCl2和2 mmol/L SA可以起到缓解作用且效果最优.本研究可为观光木的培育提供理论基础.
猜你喜欢
中老年保健(2022年3期)2022-11-21
河北果树(2020年4期)2020-11-26
科学与财富(2019年15期)2019-10-21
发酵科技通讯(2018年2期)2018-07-06
现代园艺(2018年2期)2018-03-15
现代园艺(2017年23期)2018-01-18
娃娃画报(2017年3期)2017-03-30
湖南林业科技(2017年1期)2017-02-06
动物营养学报(2015年10期)2015-12-01
幼儿智力世界(2015年3期)2015-06-16
