
不同品种大球盖菇原生质体单核化及单核体的特性
2023-05-18 07:42:20杨焕玲姜淑霞陈明杰李正鹏
上海农业学报 2023年2期
周 峰,杨焕玲,2*,赵 妍,姜淑霞,陈明杰,李正鹏,2,查 磊,2,李 玉**
(1 上海市农业科学院食用菌研究所,上海 201403;2 上海国森生物科技有限公司,上海 201403;3 山东农业大学植物保护学院,泰安 271018)
大球盖菇(Stropharia rugosoannulata)又名皱环球盖菇、酒红色球盖菇、赤松茸[1-2],不仅含有丰富的营养物质,味道鲜美,深受消费者喜爱[3],而且在实现“乡村振兴”战略中同样发挥重要作用,可以提高农田利用率、增加经济效益、改善土壤品质等[4]。 大球盖菇作为新兴发展产业面临很多亟待解决的问题,如高温环境下烧菌、易开伞,育种周期长,野生资源少,种质资源匮乏等。 为满足市场需求,丰富大球盖菇的种质资源库,定向育种迫在眉睫。 相较于传统常规的食用菌育种方法,原生质体技术开辟了食用菌育种的新天地,单核原生质体具有相对稳定而集中的亲本性状,可以减少筛选杂交后代的工作量,同时不受栽培季节限制,有效地缩短了育种周期[5]。 目前原生质体技术在食用菌育种方面也广泛应用。 高明侠等[6]将紫芝和赤芝原生质体使用电融合方法获得高多糖的灵芝。 王淑珍等[7]利用灵芝与糙皮侧耳的原生质体融合,选育木栓质化较弱的灵芝新菌株。 燕晓翠等[8]通过电融合糙皮侧耳和酿酒酵母原生质体的方法,获得了生长较快且有效降解木质素的新菌株。 郑锦荣[9]采用原生质体融合技术获得3 株金针菇与巨大口蘑融合的新菌株。 蔡佺佑[10]采用原生质体融合育种方法,以秀珍菇和巨大口蘑为亲本菌株选育出2 株新菌株。 原生质体融合的方法也应用于大球盖菇育种[11]。 朱静娴[12]利用原生质体育种技术驯化出3 株大球盖菇耐高温菌株。 任纪帆等[13]通过原生质体融合和单孢杂交选育出‘山农球盖3 号’耐高温高产菌株。 单核原生质体特殊的遗传背景,在理论上是研究其基因定位和遗传性状的重要材料,在育种上是形成以单核原生质体为材料的新方向,具有广阔的应用和推广前景[14-16]。
原生质体单核化的关键是原生质体的制备及再生,影响原生质体制备的因素有很多,主要包括酶的种类及浓度、反应温度、酶解时间、渗透压稳定剂种类及浓度等,不同的食用菌有其特定的最佳条件。 研究表明,真姬菇、褐色双孢蘑菇、蛹虫草、桑黄、草菇等食用菌原生质体制备的条件各不相同[17]。 本试验通过不同品种大球盖菇原生质体单核化,获得单核体,探究单核体菌丝形态特征,为利用原生质体技术选育新品种做铺垫,同时为大球盖菇的遗传和育种工作提供不同品种的单核体材料。
1 材料与方法
1.1 材料
1.1.1 供试菌株
本研究所用供试菌株及其来源见表1。

表1 供试菌株及其来源Table 1 Tested strains and their sources
1.1.2 主要仪器
智能食用菌培养箱(型号:ZJX-300A),杭州钱江仪器设备有限公司;洁净工作台(型号:VS-1300LU),苏净集团苏州安泰空气技术有限公司;均质仪(型号:32BL80,产地:美国);超级恒温槽,上海比郎仪器有限公司;倒置显微镜(产地:菲律宾);电子天平,梅特勒-托利多仪器有限公司。
其他试材:血球计数板,无纺布,普通漏斗,0.20 μm 细菌过滤器,砂芯漏斗,50 mL 离心管,20 mL 离心管。
1.1.3 主要试剂
溶壁酶购自广东省微生物研究所;甘露醇购自生工生物工程上海股份有限公司;PDA 粉(马铃薯葡萄糖琼脂)、PDB 粉(马铃薯葡萄糖肉汤)购自美国碧迪医疗器械(上海)有限公司。
1.1.4 培养基类型
固体培养基(加富PDA 培养基):PDA 粉39 g,KH2PO43 g,MgSO4·7H2O 1.5 g,胰蛋白胨2 g,酵母粉2 g,水1 L。 液体培养基:PDB 粉24 g,胰蛋白胨2 g,KH2PO43 g,MgSO4·7H2O 1.5 g,VB10.1 g,水1 L。再生培养基:PDA 粉39 g,胰蛋白胨2 g,酵母粉2 g,MgSO4·7H2O 1.5 g,K2HPO41.5 g,KH2PO41.5 g,VB10.1 g,VB60.1 g,甘露醇109 g,水1 L。 镜检培养基:PDA 粉0.39 g,吐温-20 20 μL,水100 mL。 以上培养基均在121 ℃高压灭菌15 min 后冷却备用。
1.2 试验方法
1.2.1 固体菌丝的制备
在无菌超净工作台中,将4 ℃冰箱保存的大球盖菇菌种25 ℃活化2 d 后,取3—5 mm 大小的菌丝块接种到固体培养基上,于25 ℃恒温黑暗条件下培养14 d,备用。
1.2.2 液体菌丝的制备
将长势良好的菌落转至均质仪,并加入50 mL 液体培养基打碎,分别接种于盛有50 mL 液体培养基的250 mL 三角瓶中,25 ℃恒温培养箱黑暗静置培养3 d,备用。
1.2.3 渗稳剂的制备
甘露醇稳渗液:称取10.9 g 甘露醇溶解于100 mL 蒸馏水中,配成0.6 mol∕L 的甘露醇稳渗液,121 ℃高压灭菌15 min 后备用。
1.2.4 酶液的制备
称取0.12 g 溶壁酶溶于8 mL 0.6 mol∕L 渗透压稳定剂,使酶液含量为1.5%,经0.20 μm 细菌过滤膜过滤除菌后备用。
1.2.5 原生质体的制备
收集并过滤液体培养3 d 的菌丝,无菌水冲洗后用无菌吸水纸吸干多余水分,将菌丝转移至酶液含量为1.5%的液体中,轻微振荡使菌丝均匀分散开,置于30 ℃水浴锅中酶解3 h,每隔1 h 轻微震荡,使其与酶液充分接触。 用G-2 砂芯漏斗过滤除去酶解液中残留的菌丝,将收集的滤液放在50 mL 离心管中,4 ℃、4 000 r∕min 离心15 min,弃上清,沉淀用0.6 mol∕L 的甘露醇洗涤2 次,即得到纯化的原生质体。
1.2.6 原生质体再生
将原生质体用0.6 mol∕L 的甘露醇稀释至适当浓度,用血球计数板计数原生质体总数。 吸取100 μL稀释好的原生质体悬液于再生培养基上,用涂布棒涂布均匀。 同时用无菌水胀破原生质体后,以同样方法涂布于再生培养基上,以消除非原生质体所形成的菌落带来的误差。 25 ℃恒温黑暗条件下倒置培养3—5 d 可见再生菌落,并统计原生质体再生菌落数。 计算各个品种原生质体的再生率:
原生质体再生率=[(原生质体再生菌落数-对照组再生菌落)∕原生质体总数] ×100% (1)
1.2.7 原生质体单核化
将原生质体再生的单个菌落(长出如针尖大小时挑出)接种在镜检培养基上,统计单个菌落总数。25 ℃培养3—5 d 后使用显微镜观察锁状联合情况,若无锁状联合则为单核体,并统计单核化率。
1.2.8 菌丝形态特征
将不同品种的大球盖菇菌株及其单核体接种于镜检培养基上,置于25 ℃恒温培养箱中黑暗培养7 d,观察菌落大小、长势、颜色等特征,分析出发菌株与单核体菌落形态的差异性。 每个试验设置4 个重复。
2 结果与分析
2.1 不同大球盖菇品种的原生质体再生率
将制备的原生质体用0.6 mol∕L 的甘露醇洗涤后得到纯净的原生质体,如图1 中黑色圆圈标出的即为原生质体。

图1 显微镜下的大球盖菇原生质体(20 ×)Fig.1 Protoplasts of Stropharia rugosoannulata under microscope(20 ×)
由表2 可知,‘土肥’品种原生质体再生能力最强,3 d就已经长满培养皿,并且菌丝浓白,气生菌丝发达,再生率最高,达1.37%;‘高邮’和‘大T’品种再生菌落均4 d 长满培养皿,‘高邮’菌丝较稀疏且分散,再生率为1.13%,菌落由白色浓密菌丝组成;‘大T’菌丝蔓延扩展,呈扩散状,菌落较大,菌丝先由透明变为白色,在试验的3 个品种中再生率最低,为0.90%。

表2 不同品种大球盖菇原生质体再生率Table 2 Regeneration rate of protoplasts of different strains of Stropharia rugosoannulata
2.2 不同品种大球盖菇的原生质体单核化率
从表3 可以看出,供试验的3 个大球盖菇品种的单核化率‘大T’品种的最高,达到85.2%,‘土肥’的单核化率次之,为66.0%,‘高邮’的单核化率最低为59.8%。 单个菌落接种在镜检培养基上,可能由于培养基较薄,营养物质不够充足,生长的菌丝较细,在倒置显微镜下难以观察锁状联合,待菌丝生长5 d 后才能清晰观察到有无锁状联合。

表3 不同品种大球盖菇原生质体单核化率Table 3 Monokaryotization rate of protoplasts of different strains of Stropharia rugosoannulata
2.3 不同品种大球盖菇的单、双核体菌落特性
观察菌落大小、长势、颜色、形态等特征,统计不同品种大球盖菇单、双核菌丝体菌落形态的特性。 如图2 及表4,大球盖菇的不同品种菌落形态差别明显,而同一品种的单核体和双核体在菌落形态上差别较小。 使用镜检培养基,提高了培养基的透明度,观察更为清晰明显,但生长速度较普通PDA 培养基慢。‘土肥’品种菌丝生长非常稀疏,菌落颜色为白色,有同心轮纹,微凸起,边缘整齐,单核和双核体的菌落形态略微有差别,双核体的菌落边缘有气生菌丝生长,但单核体边缘菌丝匍匐,生长不旺盛,多为基内菌丝。 ‘大T’品种菌丝生长旺盛,浓密,长势良好,边缘整齐,菌落呈放射状,圆形,平坦,单双核无明显差异。‘高邮’品种单双核菌丝体差异较大:单核体菌落呈放射状,平坦,基内菌丝多,气生菌丝较少,菌丝稀疏;双核体菌丝生长旺盛,菌落白色,边缘不整齐,呈绒毡状,气生菌丝发达,菌丝浓密。
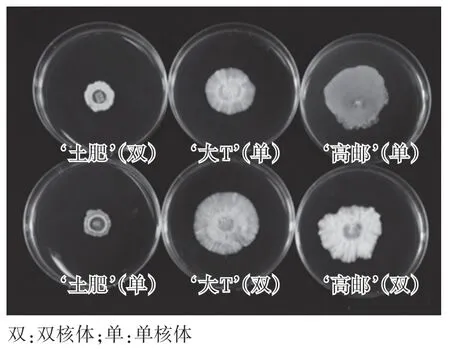
图2 不同品种大球盖菇的菌落形态Fig.2 Colony morphology of different strains of Stropharia rugosoannulata

表4 不同品种大球盖菇的菌落特性Table 4 Colony characteristics of different strains of Stropharia rugosoannulata
3 讨论与结论
大球盖菇在温度较高的环境下原基难分化、菌盖易开伞,品质和产量均急剧下降[12],为解决大球盖菇发展的瓶颈,可通过快速定向育种的途径丰富大球盖菇种质资源,单核原生质体成为探究遗传背景及开展原生质体技术育种的重要前提。
原生质体再生率的高低是原生质体技术应用的关键。 本研究采用0.6 mol∕L 的甘露醇做渗透压稳定剂,制备菌龄3 d 的菌丝和含量为1.5%的酶液,在30 ℃恒温水浴锅中酶解3 h,滤液在4 ℃、4 000 r∕min条件下离心15 min,再生培养基在普通的PDA 培养基的基础上加入VB1、VB6,以促进进原生质体再生。邹彰毅等[18]在再生培养基中添加纤维二糖和VB1,提高了姬松茸原生质体的再生率,此结果在本试验中再次得到验证。 大球盖菇中供试验的3 个不同品种再生率均较高,‘土肥’品种再生率最高,达1.37%,‘高邮’品种再生率次之为1.13%,‘大T’品种的再生率最低为0.90%。 许梅[19]用双层平板法得到大球盖菇再生率为0.96%,远低于‘土肥’和‘高邮’两个品种,进一步说明大球盖菇的再生率与不同品种相关,本试验原生质体再生的配方为最佳条件。 茶树菇再生菌落中异核体与单核体存在时间差,8—10 d 异核体菌落先生成,10 d 之后产生单核体菌落[20]。 但是,本试验表明大球盖菇原生质体再生速度较快,原生质体单核化率高,且单核体和双核体再生的菌落几乎同时形成,这与闫培生等[11]和Yan 等[21]的结果相符。
原生质体单核化是快速获得单核体杂交亲本的有效手段,已从许多食用菌中通过单核化获得单核体。 本研究获得了不同品种大球盖菇的原生质体单核体,供试验的三个大球盖菇品种的单核化率‘大T’品种高达85.2%;‘土肥’品种单核化率次之,为66.0%;‘高邮’品种单核化率最低,为59.8%。 ‘大T’品种单核化率是已报道过的食用菌中单核化率最高的,这也丰富了食用菌原生质体单核化和遗传育种研究的内容。 ‘土肥’和‘高邮’品种的单核体与双核体在菌落形态及生长速度上有所差别,‘土肥’品种菌丝生长局限,非常稀疏,菌落颜色为白色,有同心轮纹,微凸起,边缘整齐,单核和双核体的菌落形态略微有差别,双核体的菌落边缘有气生菌丝生长,但单核体边缘菌丝匍匐,生长不旺盛,多为基内菌丝。 ‘高邮’品种菌丝蔓延生长,边缘不整齐,菌落白色,培养皿上单双核菌丝体差异较大,单核体菌落呈放射状,平坦,基内菌丝多,气生菌丝较少,菌丝稀疏,双核体菌落呈绒毡状,气生菌丝发达,菌丝浓密。 目前,已使用本试验选育的原生质体单核体进行了高温菌种的选育,并获得高温大球盖菇品种[12]。
将不同品种大球盖菇原生质体单核化可获得遗传背景单一的单核体,初步探究单核体的菌落特性,后续对其进行分类确定交配型,不仅可为构建大球盖菇遗传体系提供原材料,同时还可为定向育种提供更多材料,进一步丰富大球盖菇种质资源。
猜你喜欢
四川蚕业(2022年2期)2022-11-19 02:10:30
房地产导刊(2020年6期)2020-07-25 01:31:24
四川蚕业(2020年3期)2020-07-16 08:09:38
农家科技中旬版(2018年8期)2018-10-15 00:49:38
食用菌(2017年5期)2017-10-19 03:02:28
食药用菌(2016年6期)2016-03-01 03:24:29
大连工业大学学报(2015年4期)2015-12-11 04:06:50
特产研究(2015年1期)2015-04-12 06:36:11
中国房地产·市场版(2014年7期)2014-08-07 12:59:58
食品工业科技(2014年23期)2014-03-11 18:19:06
