
极耐热α-L-鼠李糖苷酶的性质及其在异槲皮素制备中的应用研究
2023-03-09 04:27:14张珊珊卢昌宁赵林果
食品与生物技术学报 2023年1期
卢 姗,张珊珊,邹 康,卢昌宁,赵林果
(南京林业大学 化学工程学院,江苏 南京 210037)
异槲皮素(isoquercitrin),又名异槲皮素-3-O-葡萄糖苷[1],具有抗氧化[2]、抑制肿瘤与癌症[3]、调节免疫功能[4]、降低血糖血脂[5]等药理作用,还可用于抗抑郁药物的研发[6],但其在植物中的含量极低,无法使用传统的提取方法进行大规模制备[7]。研究表明,芦丁(rutin)与异槲皮素拥有相同的母核结构,仅在异槲皮素葡萄糖基的C6 位多连接了一个鼠李糖[8],其在植物中含量较高,如槐花米中的含量高达28%[9],但其药理活性、生物利用度等均不如异槲皮素[10]。因此,可以使用芦丁作为转化的底物制备异槲皮素。相较于化学法,生物转化法因其条件温和、高效环保、副产物少等优点,已成为大规模制备异槲皮素的一种较为可行的途径[11]。值得注意的是,生物转化法转化芦丁制备异槲皮素的关键是寻找高效的能作用于α-1,6-鼠李糖苷键的α-L-鼠李糖苷酶。
α-L-鼠李糖苷酶(EC 3.2.1.40)属于糖苷水解酶,可特异性切除糖苷类化合物非还原末端的α-L-鼠李糖基[12],在食品工业和医药领域有着广泛的应用[13-17]。目前已研究了多种细菌[18]和真菌[19]来源的α-L-鼠李糖苷酶,其中大部分α-L-鼠李糖苷酶最适反应温度在40~60 ℃[18-20],属于中温酶,较难满足一些工业生产所需。另一方面,天然产物大多是水难溶或水不溶的化合物,在实际的工业生产中,常借助高温增加底物溶解度,提高传质速率,从而提高转化效率,同时较高的反应温度还能降低体系黏度和避免杂菌污染等[21],但这对参与反应的酶蛋白的热稳定性有着很高的要求。研究发现嗜热菌来源的酶蛋白普遍表现出优异的热稳定性[22],在较高的反应温度下也能长时间保持良好的催化活力。然而,目前仅报道过少数嗜热菌来源的α-L-鼠李糖苷酶,如嗜热菌Thermomicrobium sp.[23]、Dictyoglomus thermophilum[24-25]和Thermotoga petrophila DSM 13995[26]等。因此,嗜热菌来源的α-L-鼠李糖苷酶资源值得进一步研究和开发。
在前期研究中,作者已从NCBI 数据库中筛选到嗜热菌Sulfolobus islandicus(NC_021058.1)来源的α-L-鼠李糖苷酶SisRha 的基因序列,对其进行了重组质粒的构建及表达。在此基础上,作者首先优化了重组酶SisRha 的表达条件,接着测定其酶学性质,然后优化了重组酶SisRha 制备异槲皮素的工艺条件。研究结果丰富了现有的α-L-鼠李糖苷酶资源,为嗜热菌来源的α-L-鼠李糖苷酶的研究奠定了基础,同时提供了一种高反应温度下转化芦丁制备异槲皮素的方法。
1 材料与方法
1.1 菌种与质粒
α-L-鼠李糖苷酶基因sisRha 来源于嗜热菌Sulfolobus islandicus(NC_021058.1),基因片段的全长为2 652 bp,可编码883 个氨基酸,理论蛋白质相对分子质量约为1.005 4×103,预测的等电点pI 为6.14。目的序列的全基因合成由上海捷瑞生物工程有限公司进行,表达宿主菌株Escherichia coli(E.coil)BL21(DE3)购自南京诺唯赞生物技术有限公司。
1.2 主要试剂
乙腈、甲醇、乙醇、二甲基亚砜(DMSO)、磷酸氢二钾、磷酸二氢钾及其他常见生化试剂(均为国产分析纯):国药集团(上海)化学试剂有限公司产品;酵母提取物、蛋白胨:英国Oxoid 公司产品;麦芽糊精:南京姜华化玻璃有限公司产品;芦丁:上海甄准生物科技有限公司产品;异槲皮素标准品:成都曼思特生物技术有限公司产品;硫酸卡那霉素、异丙基-β-D-硫代半乳糖苷(IPTG):生物工程上海股份有限公司产品;蛋白质电泳缓冲液、预混蛋白质电泳Marker:南京天为公司产品;对硝基苯酚鼠李糖糖苷(pNPR)、考马斯亮蓝G-250:美国Sigma 公司产品;Bradford 蛋白质含量检测试剂盒:日本TaKaRa 公司产品;PAGE 凝胶快速制备试剂盒、质粒提取试剂盒:上海雅酶生物科技有限公司产品。
1.3 培养基
LB 培养基:1.0 g/dL 胰蛋白胨,0.5 g/dL 酵母提取物,1.0 g/dL 氯化钠。
TB 培养基:A 液为1 g/dL 麦芽糊精(或其他碳源),1.2 g/dL 胰蛋白胨,2.4 g/dL 酵母提取物;B 液为磷酸二氢钾2.3 g/dL,磷酸氢二钾16.43 g/dL。A液和B 液体积比为9∶1。
1.4 主要仪器与设备
SX-500 型全自动高压蒸汽灭菌器:日本TOMY公司产品;HH-4 型恒温水浴锅:国华电器(常州)有限公司产品;SHP-250 型生化培养箱:上海精宏实验设备有限公司产品;U-1800 型分光光度计:日本Hitachi 公司产品;超声波细胞破碎机:宁波新芝生物科技有限公司产品;5415R 型高速离心机、温度梯度PCR 仪:德国Eppendorf 公司产品;AIR TECH US-1300L-U 型超净工作台:苏州安泰空气技术有限公司产品;凝胶成像系统:美国BIO-RAD 公司产品;Agilent 1260 型高效液相色谱仪:美国安捷伦科技有限公司产品。
1.5 实验方法
1.5.1 重组菌的构建及表达 将重组质粒pET-28-sisRha 热激转化至E.coli BL21(DE3)的感受态细胞中,采用含有质量浓度50 μg/mL 卡那霉素的LB 平板培养基进行筛选,37 ℃过夜培养后长出阳性转化子,挑取单菌落至5 mL LB 试管中,于37 ℃摇床过夜培养,使用30%(体积分数)的甘油将菌种保存于-80 ℃冰箱。取5 μL 菌液加入含50 μg/mL卡那霉素抗性的5 mL LB 试管中,180 r/min、37 ℃摇床过夜复苏。
将500 μL 复苏后的菌液转接到50 mL LB 培养基中,37 ℃、180 r/min 摇床培养至OD600为0.6~0.8,使用0.01 mmol/L 异丙基-β-D-硫代半乳糖苷(IPTG)诱导表达,28 ℃、180 r/min 摇床培养8 h。8 000 r/min、16 ℃离心10 min 后收集菌体。加入5 mL 25 mmol/L 的磷酸盐缓冲液重悬菌体,超声破碎后于10 000 r/min、16 ℃离心15 min,测定上清液酶活力。
1.5.2 酶活力的测定 以5 mmol/L pNPR 为底物,水解得到的对硝基苯酚(pNP)与碳酸钠发生显色反应,在405 nm 的波长下测定产物的吸光度。反应体系为:10 μL 500 mmol/L 柠檬酸-磷酸氢二钠缓冲液,20 μL 底物,65 μL 去离子水,混匀预热后加入5 μL 酶,在酶的最适温度和pH 下反应5 min,以不加酶液的反应体系作空白对照[22]。酶活力单位定义参照文献[22]。pNP 标准曲线为y=2.376x+0.048,决定系数R2=0.999 0。酶活计算公式为:
式中:E 为α-L-鼠李糖苷酶酶活力,U/mL;c 为酶反应后的pNP 浓度,μmol/L;t 为酶与底物反应时间,min;Vs为反应体系的体积,mL;Ve为反应体系中酶液的体积,mL;N 为酶液的稀释倍数。
1.5.3 重组酶SisRha 的表达条件优化 首先优化TB 培养基的碳源种类,将复苏菌液500 μL 分别接种至5 瓶50 mL 含质量浓度1.00 g/dL 不同碳源(葡萄糖、蔗糖、甘油、菊粉、麦芽糊精)的TB 培养基中,37 ℃、180 r/min 摇床培养至OD600为0.6~0.8,于0.02 mmo/L IPTG 中诱导,28 ℃、180 r/min 摇床培养48 h 后分别测定粗酶酶活力,酶活力测定方法见1.5.2。然后对TB 培养基的碳源质量浓度(0.10、0.25、0.50、1.00、2.00、4.00 g/dL)、诱导温度(20、25、28、32、37、42 ℃)以及IPTG 浓度(0、0.005、0.010、0.050、0.100、0.250 mmol/L)进行优化。重组菌的培养与酶活力测定方法与上述一致。
1.5.4 重组酶SisRha 的纯化 粗酶液置于60 ℃水浴锅内热处理30 min,10 000 r/min、16 ℃离心15 min 后留上清液。采用镍离子亲和层析柱纯化上清液,用含不同浓度咪唑的磷酸缓冲液洗脱并收集洗脱液,再经透析后得到纯酶。采用十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE)分析纯酶,考马斯亮蓝法测定蛋白质质量。
1.5.5 重组酶SisRha 的酶学性质
1)最适温度和最适pH 分别将重组酶置于70~100 ℃(不同温度间隔5 ℃)的水浴温度和pH 为4.0~8.0(不同pH 间隔0.5)的缓冲液中反应后测定吸光度,并计算相对酶活力,每次只改变一个变量,以测得的最高酶活力为100%,确定最适温度和最适pH。
2)热稳定性和pH 稳定性 将重组酶分别置于80、85、90 ℃水浴锅 中分别保温5、10、15、30、60、90、120 min 后,在其最适温度和最适pH 下测定剩余酶活力(相对酶活力),探究其热稳定性;70 ℃下将酶置于pH 为4.0~7.5(不同pH 间隔0.5)的缓冲溶液保温1 h 测定剩余酶活力(相对酶活力),探究其pH 稳定性。
3)金属离子耐受性 向反应体系中分别添加终浓度为1 mmol/L 和3 mmol/L 的Fe3+、Ca2+、Na+、K+、Mg2+、Zn2+等金属离子盐溶液,以不含金属离子的酶活力为100%,计算不同金属离子体系的相对酶活力。
4)有机溶剂和鼠李糖耐受性 向反应体系添加体积分数0、1%、2%、5%、10%、15%、20%、25%、30%、40%的甲醇、乙醇、二甲基亚砜(DMSO),以不含有机溶剂的酶活力为100%,测定有机溶剂对酶活力的影响;添加浓度0、25、50、100、150、200、250 mmol/L 的鼠李糖,以不加鼠李糖的酶活力为100%,探究鼠李糖(即产物)对酶活力的影响。
5)动力学参数测定和底物特异性分析 将重组酶加至浓度0.2~1.0 mmol/L(不同浓度间隔0.1 mmol/L)的人工底物pNPR 中反应5 min 后测定其吸光度,按照米氏动力学方程计算其Vmax、Km及kcat;再分别以50 mmol/L 的芦丁、淫羊藿苷、朝藿定C、橘皮苷、柚皮苷为底物,于最适条件下测定重组酶SisRha 的酶活力,以底物pNPR 的酶活力为100%,探究该酶对不同天然底物的底物特异性。
1.5.6 产物HPLC 检测 所用色谱柱为Agilent E clipse XDB-C18(5 μm,4.6 mm×250 mm),流量0.8 mL/min,进样量10 μL,检测波长368 nm,柱温40 ℃;流动相为甲醇与0.1%(质量分数)甲酸水混合液(体积比为40∶60)。芦丁出峰时间为13.091 min,异槲皮素为14.545 min[11]。以不同质量浓度异槲皮素及相对应的色谱峰面积绘制标准曲线,为y=21 456 x-245.42,决定系数R2=0.999 5。
1.5.7 重组酶SisRha 转化芦丁的条件优化
1)转化温度和pH 优化 构建100 μL 的反应体系,底物芦丁为2 mmol/L,加酶量0.1 U/mL,柠檬酸-磷酸氢二钠缓冲液浓度50 mmol/L,去离子水补足至100 μL。分别在70~90 ℃(不同温度间隔5 ℃)的水浴锅和pH 4.5~7.0(不同pH 间隔0.5)的缓冲液中反应2 h 后,加入400 μL 甲醇终止反应,样品离心后,由0.22 μm 滤膜过滤,HPLC 检测异槲皮素的浓度并计算其产率,确定最适转化温度和pH。
2)加酶量优化 在最适转化温度和pH 下,加入0、0.02、0.04、0.10、0.20、0.40、0.60、0.80、1.00 U/mL 的重组酶SisRha,反应2 h 后HPLC 检测,确定最适加酶量。
3)反应历程检测 在最适转化温度、pH 和加酶量的条件下,分别于0、10、20、30、40、50、60、70、80、90、120 min 时取样,加入400 μL 甲醇终止反应,样品离心后,由0.22 μm 滤膜过滤后,HPLC 检测,探究底物浓度与转化时间的关系。
2 结果与分析
2.1 重组酶SisRha 的表达条件优化
重组菌目的蛋白质的表达量条件优化的结果如图1 所示。通过对比不同碳源培养条件下SisRha的相对酶活力,发现菊粉为最合适的碳源(见图1(a));在对碳源含量进行优化时,培养基中菊粉的质量浓度为0.50 g/dL 和1.00 g/dL 时,重组酶SisRha 的相对酶活力最高且二者几乎相同(见图1(b));当诱导温度为37 ℃(见图1(c))且不添加诱导剂时(图1(d)),重组酶的酶活力最高,即使极微量的IPTG(0.005 mmol/L)都会降低目的蛋白质的表达量,这可能是由于培养基中微量的乳糖足以启动目的基因的转录,其本底表达即可获得极高的表达。说明该酶的表达不需要依赖诱导剂IPTG,这在实际应用中更有前景。综上所述,重组酶SisRha 的最佳表达条件为:以0.50 g/dL 的菊粉作TB 培养基的碳源,不添加诱导剂,于37 ℃摇床培养48 h,表达量最高可至0.92 U/mL,为LB 培养基中的最佳表达量(0.16 U/mL)的5.75 倍。
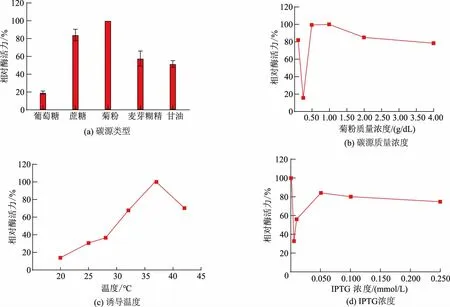
图1 重组酶SisRha 在TB 培养基中的表达条件优化Fig.1 Optimization of expression conditions for recombinant enzyme SisRha in TB medium
2.2 重组酶SisRha 的纯化及SDS-PAGE 分析
通过超声破碎、热处理、亲和层析等方法对重组酶SisRha 进行了纯化,并对粗酶液(超声破碎)、热处理的酶液以及亲和层析后的纯酶进行SDSPAGE 分析,结果如图2 所示,通过热处理,能去除大量非目的蛋白质,很大程度上降低了目的蛋白质后续的纯化难度,通过亲和层析,获得了较纯的目的蛋白质,在1.0×105左右有单一的目的条带。如表1 所示,纯酶的比活力为10.76 U/mg,纯化倍数是粗酶的12.92 倍,得率(纯酶总酶活力/粗酶总酶活力×100%)为44.35%。获得的纯酶可应用于后续的酶学性质测定及芦丁的转化实验。

图2 重组酶SisRha 的SDS-PAGE 分析Fig.2 SDS-PAGE analysis of recombinant enzyme SisRha

表1 重组酶SisRha 的纯化及蛋白质质量Table 1 Purification and protein content of recombinant enzyme SisRha
2.3 重组酶SisRha 的酶学性质
2.3.1 重组酶SisRha 的反应条件测定
1)最适反应温度及热稳定性 重组α-L-鼠李糖苷酶SisRha 来源于嗜热菌Sulfolobus islandicus,其最适生长温度在80 ℃以上,该重组酶很有可能为高温酶。因而本研究中使用人工底物pNPR 对重组酶SisRha 的最适温度进行测定,结果表明,重组酶SisRha 最适温度为90 ℃(见图3(a)),比重组α-L-鼠李糖苷酶PRHA2 的最适温度高出30 ℃[27]。
在实际应用中确定催化反应温度时,除了考虑酶能保持较高的反应速率外,还应考虑该温度下是否具有较为优良的热稳定性,其热稳定性结果如图3(b)所示,重组酶SisRha 在80 ℃下保温2 h 后酶活力几乎没有改变,而嗜热菌来源的α-L-鼠李糖苷酶TpeRha 在同样温度下保温2 h 后酶活力损失50%左右[26],说明该酶具备优异的热稳定性。

图3 重组酶SisRha 的最适反应温度及热稳定性Fig.3 Optimum temperature and thermostability of recombinant enzyme SisRha
2)最适反应pH 及pH 稳定性 pH 环境的不同,不仅会很大程度上影响酶的催化反应速率,还会对酶蛋白的稳定性产生一定的影响。因此,本研究中对重组酶SisRha 的最适反应pH 和该酶在不同pH 条件下的稳定性进行了测定,结果如图4 所示。与RhmA 最适反应pH 7.8 不同[23],重组酶SisRha 的最适pH 为5.5(见图4(a)),且该酶在pH 4.5~6.5 时相对酶活力保持在80%以上。70 ℃下,重组酶SisRha 在pH 4.5~7.0 的缓冲体系中孵育1 h,均保留80%以上的残余酶活力(见图4(b)),这与该酶最适pH 测定曲线的趋势相同,进一步证实该酶在弱酸环境下稳定性较好。

图4 重组酶SisRha 的最适pH 和pH 稳定性Fig.4 Optimum pH and pH stability of recombinant enzyme SisRha
2.3.2 金属离子对重组酶SisRha 催化活性的影响部分金属离子和螯合剂对重组酶SisRha 酶活力的影响如表2 所示,少数金属离子对重组酶SisRha 具有微弱的激活作用,Mn2+具有相对明显的激活作用,Cu2+和Zn2+则一定程度上抑制SisRha 的酶活力;螯合剂EDTA 对重组酶SisRha 的酶活力没有显著的影响,因此金属离子对重组酶SisRha 的催化活性和三维结构不起作用[28]。

表2 金属离子对重组酶SisRha 的影响Table 2 Effects of metal ions on recombinant enzymes SisRha
2.3.3 重组酶SisRha 的有机溶剂和鼠李糖耐受性在工业生产过程中,常添加一定浓度的有机溶剂增加底物的溶解度,常用的助溶剂如甲醇、乙醇、二甲基亚砜(DMSO)等。本研究中以pNPR 为底物,在反应体系中分别加入不同体积分数的甲醇、乙醇、DMSO 测定了重组酶SisRha 的有机溶剂耐受性。结果如图5(a)所示,重组酶SisRha 对DMSO 的耐受性优于甲醇和乙醇,DMSO 体积分数为10%时,残余酶活力(相对酶活力)大于50%。
在糖苷水解酶反应过程中,产物糖的积累会对酶的催化效率产生一定程度的抑制作用,因此,对产物糖的耐受能力的大小,一定程度上决定了糖苷水解酶能水解的底物浓度大小以及催化效率的高低。为确定重组酶SisRha 对产物鼠李糖的耐受能力,本研究中以pNPR 为底物,在反应体系中分别加入不同终浓度的L-鼠李糖,测定重组酶SisRha 的鼠李糖耐受性。结果显示,约添加20 mmol/L 鼠李糖时,该酶保留有50%左右的残余酶活力(相对酶活力,见图5(b))。
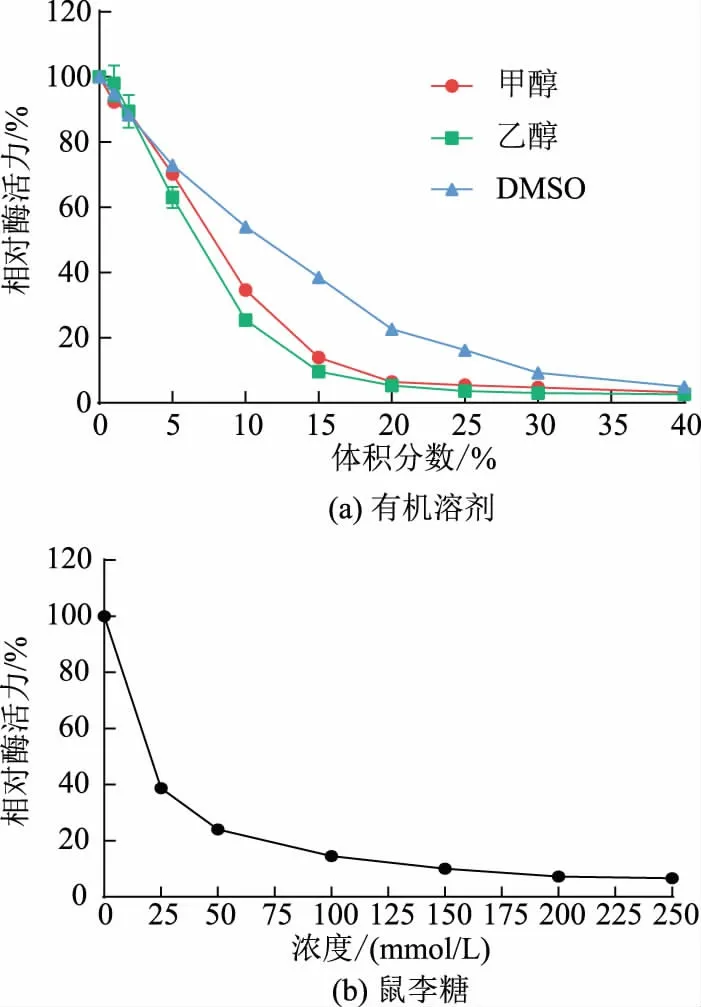
图5 重组酶SisRha 的有机溶剂和鼠李糖耐受性Fig.5 Tolerance of organic solvent and rhamnose on recombinant enzyme SisRha
2.3.4 重组酶SisRha 的动力学参数及底物特异性分析 为确定重组酶SisRha 的底物特异性,首先选取人工底物pNPR、pNPG、pNPX、pNPAraf、pNPArap作为底物进行反应,以确定重组酶SisRha 能切除的糖苷类型,结果如表3 所示。重组酶SisRha 仅作用于人工底物pNPR(与鼠李糖相连),比活力为25.06 U/mg,而对pNPG(与葡萄糖相连)、pNPX(与木糖相连)、pNPAraf(与阿拉伯呋喃糖相连)和pNPArap(与阿拉伯吡喃糖相连)等其他人工底物无作用。由此可以确定重组酶SisRha 为鼠李糖苷酶,仅专一识别鼠李糖苷键,而对其他种类的糖苷键没有催化作用。
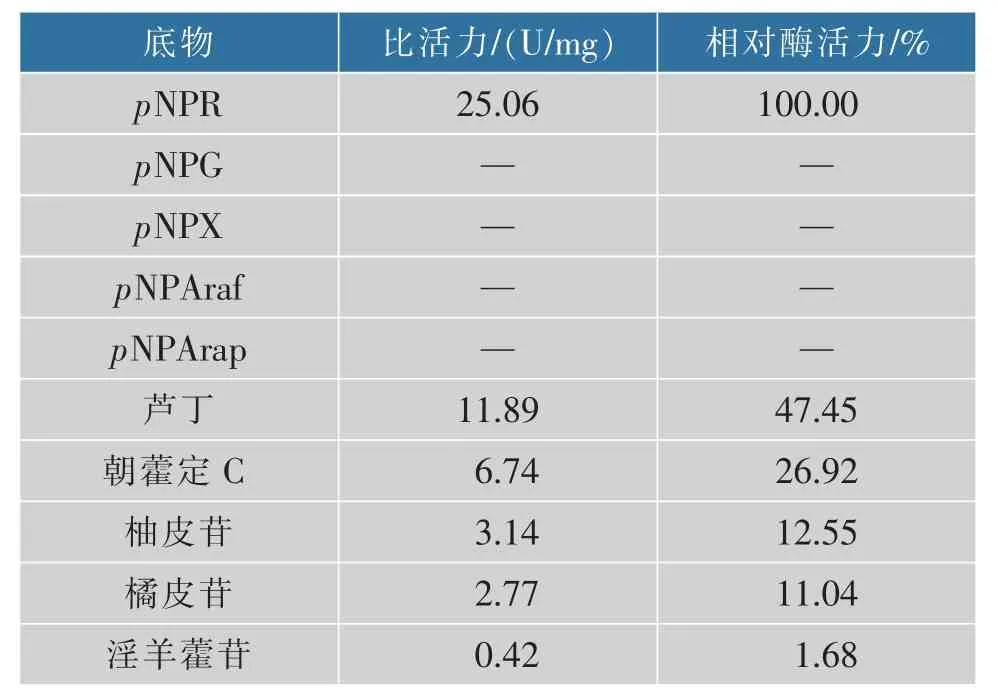
表3 重组酶SisRha 的底物特异性Table 3 Substrate specificity of recombinant enzyme SisRha
随后测定了重组酶SisRha 对几种典型的含有鼠李糖基的黄酮类化合物的催化能力,相较于人工底物pNPR(100%),重组酶SisRha 对芦丁(α-1,6-鼠李糖苷键)的催化效率最高,表现出47.45%的相对酶活力,朝藿定C(α-1,2-鼠李糖苷键)次之,对橘皮苷(α-1,6-鼠李糖苷键)和柚皮苷(α-1,2-鼠李糖苷键)只有大约10%的相对酶活力,对淫羊藿苷(α-1-鼠李糖苷键)几乎无酶活力。这是由于不同的糖苷键类型及与母核连接的位置会影响酶的催化活性中心与底物的结合,从而影响酶的催化活性[29]。芦丁和橘皮苷都通过α-1,6-鼠李糖苷键与β-D-葡萄糖连接,芦丁的糖基连在黄酮的C3 位,而橘皮苷的糖基连在C7 位,两者的空间结构差异导致酶的催化活性有所不同。相较于切除α-1,2-鼠李糖苷键,重组酶SisRha 对α-1,6-鼠李糖苷键的切除能力更强,并对连在C3 位置的糖基的切除效果更好,这与RhmA 的底物特异性结果刚好相反[23]。随后又进一步研究了重组酶SisRha 对pNPR 的动力学常数,以确定其底物亲和力和催化效率。结果如表4所示,重组鼠李糖苷酶SisRha 对人工底物pNPR 亲和力较好,Km值为(0.15±0.03)mmol/L,Vmax值为(6.26±0.51)U/mg,Kcat值为(10.48±0.86)s-1;Kcat/Km为(73.28±11.56)L/(s·mmol)。

表4 重组酶SisRha 的反应动力学参数Table 4 Kinetic parameters of recombinant enzyme SisRha
2.4 重组酶SisRha 转化芦丁的条件优化
由2.3.4 的结果可知,在天然底物的催化中,重组酶SisRha 对芦丁的催化能力最强,因此本研究中选取芦丁作为转化底物,对该酶水解芦丁制备异槲皮素的反应条件进行进一步探究。首先优化了重组酶SisRha 转化芦丁的反应温度、pH 以及加酶量,结果如图6 所示。当反应温度为85 ℃(见图6(a)),反应pH 为5.0 时(见图6(b)),芦丁的转化效率最高。这与重组酶SisRha 的最适反应温度和pH 相比均有所下降,主要是由于芦丁转化时间比pNPR 反应时间长,转化过程中既要使酶发挥最佳的催化活性,又要确保酶的稳定性不能太差。而该酶的酶学性质表明85 ℃时既能发挥相对较高的催化活性还能避免酶活力的大量损失;另一方面,不同底物与酶的催化活性中心结合情况不同,导致最适反应pH 也有所差异。在此基础上又探究了重组酶SisRha 的加酶量对芦丁转化率的影响,结果表明仅添加0.40 U/mL 重组酶SisRha,可将2 mmol/L 底物芦丁几乎完全转化,摩尔转化率高达98.56%(见图6(c)),相较于酸水解法转化率提高了50%左右[30]。
为探究重组酶SisRha 转化芦丁生成异槲皮素的反应中底物与产物的变化趋势,对反应历程进行了监测。结果如图6(d)所示,反应初始时,反应体系内仅有2 mmol/L 芦丁存在,随着反应时间的不断增加,底物芦丁的浓度逐渐减少,同时伴随着产物异槲皮素浓度的逐渐上升。至反应60 min 左右,产物异槲皮素的浓度达到最高水平,并保持稳定,而底物芦丁则几乎完全转化(见图6(d))。

图6 重组酶SisRha 转化芦丁的条件优化与反应历程Fig.6 Optimization of conversion conditions for rutin with recombinant enzyme SisRha and the reaction process
3 结语
作者表达定性了一种嗜热菌Sulfolobus islandicus来源的α-L-鼠李糖苷酶SisRha,重组酶SisRha 最适反应温度为90 ℃,具有优异的热稳定性,并且在pH 4.5~7.0 的环境下能保持优良的稳定性。重组酶SisRha 具有切除α-L-鼠李糖苷键(与苷元直接相连)、α-1,6-鼠李糖苷键、α-1,2-鼠李糖苷键的能力。该酶对天然底物芦丁的催化效率较高,利用该酶水解芦丁制备异槲皮素的适宜反应条件为用酶量0.40 U/mL,在85 ℃、pH 为5.0 的反应体系中反应60 min。此条件下,可将2 mmol/L 底物芦丁几乎完全转化,摩尔转化率可达98.56%。本研究不仅丰富了现有α-L-鼠李糖苷酶资源,为嗜热菌来源的α-L-鼠李糖苷酶的研究奠定了基础,同时还提供了一种高温条件下转化芦丁制备异槲皮素的方法,为生物酶法制备异槲皮素的研究提供了技术支撑。
猜你喜欢
现代食品科技(2023年12期)2024-01-09 01:22:38
作文与考试·小学高年级版(2023年22期)2023-03-14 14:32:40
西南石油大学学报(自然科学版)(2019年5期)2019-12-20 07:00:52
中国洗涤用品工业(2019年4期)2019-05-11 09:27:26
中成药(2017年3期)2017-05-17 06:09:13
食品与发酵工业(2015年11期)2015-12-25 02:01:06
药学研究(2015年11期)2015-12-19 11:10:54
生物加工过程(2015年1期)2015-11-11 04:01:25
分析科学学报(2015年3期)2015-10-18 02:25:40
广西科技大学学报(2015年4期)2015-02-27 12:22:22
