
丁酸钠与沸石混合添加对蛋鸡盲肠内容物体外发酵的影响
2023-02-13 09:49莫庆楠冯英杰彭晨慕雪李裕卫刘昌林詹周荣吴国云臧一天
饲料工业 2023年1期
■莫庆楠 冯英杰 彭晨慕雪 李裕卫 刘昌林 詹周荣 吴国云 臧一天*
(1.江西农业大学动物科学技术学院,南昌市动物健康与安全生产重点实验室,江西南昌 330045;2.南昌市农业科学院,江西南昌 330299;3.景德镇市农业科学研究所,江西景德镇 333000)
人们对鸡蛋的需求量日益增大,促使蛋鸡养殖产业规模不断扩大。集约化蛋鸡养殖已成为主要生产方式,这种养殖方式提高生产效率同时也加剧了禽舍内臭气的排放[1]。有研究表明氨气是臭气中的主要成分,占总臭气量的70%~90%[2]。氨气是一种带刺激性臭味的无色碱性气体,可损伤蛋鸡的呼吸道和眼结膜,增加患病死淘风险[3-4];同时高浓度氨气还会降低蛋鸡采食量,最终导致产蛋率下降[5]。因此减少蛋鸡饲养过程中的氨气排放十分必要。
目前,主要的蛋鸡源头氨气减排措施是改善饲粮配比和使用饲料添加剂[5-6]。朱春红等[6]在研究中提到日粮中添加有机酸可实现氨气减排目的。丁酸钠作为有机酸盐的一种,溶于水时多以丁酸根的形式存在,这点与丁酸类似。目前已有许多相关研究报道丁酸钠作为减排添加剂的效果,丁酸钠可以抑制蛋鸡盲肠内脲酶和尿酸酶活性,减少尿素和尿酸降解[7]。此外,王安平[1]通过试验证明,在蛋鸡盲肠内容物的体外发酵试验中添加0.15%的丁酸钠减排效果最佳。沸石是一种天然硅铝酸盐矿物,具有多孔洞结构,比表面积大,对氨具有吸附性[8-9]。目前沸石常用作饲料添加剂应用于动物生产中,Fendri 等[10]研究发现给蛋鸡饲喂含1%或2%沸石的日粮后可有效提高鸡蛋品质;而Lon-Wo等[11]的研究更直接表明,在饲粮中添加3%天然沸石可有效降低蛋鸡粪便中的氨挥发。罗一鸣等[12]的研究中也有类似发现,在鸡粪堆肥的过程中添加沸石可吸附氨态氮,减少氨气挥发。
王权[13]曾在其研究中提到,在好氧堆肥过程中添加矿物和化学物质可加速堆肥进程减少环境污染,但单一添加剂的促进作用具有局限性。钟智康[5]在其试验的前期研究中也有类似发现,单一使用丁酸钠调控蛋鸡减排氨气的效率仅有24.6%。因此,混合添加使用多种添加剂可能是一个更加高效且经济的减排方法,这在Wu-Haan 等[14]的研究中得到了印证,在稍微降低粗蛋白的蛋鸡日粮中混合添加酸化剂(CaSO4)和沸石可以有效减少氨气排放,减排效率达39%。而丁酸钠作为有机酸化剂的一种,常被作为饲料添加剂应用于实际生产中[15]。丁酸钠很可能也具有促进沸石减少氨气排放的效果。据此推测丁酸钠与沸石混合使用可以更加有效地达到减排目的。但丁酸钠和沸石混合使用的具体减排效果以及减排的机理并未有相关报道。本研究旨在探究丁酸钠与沸石混合添加对蛋鸡盲肠内容物体外发酵的影响,采用体外发酵技术,比较不同添加处理后,盲肠内容物体外发酵的产氨量、脲酶活性、尿素含量以及微生物群落的差异,从而解决混合添加丁酸钠与沸石是否优于单独添加丁酸钠或沸石的问题,并尝试探究丁酸钠与沸石混合减排的机理。为后续探究丁酸钠与沸石混合添加在蛋鸡上的应用积累数据,并为蛋鸡的绿色生产提供参考。
1 材料与方法
1.1 试验材料
1.1.1 试剂和设备
丁酸钠:98%;NH4HCO3:分析纯;CoCl2·6H2O:分析纯;FeCl2·6H2O:98%;Na2S·9H2O:分析纯;刃天青:90%;酒石酸钾钠:99%;苯酚钠:98%(上海麦克林生化科技有限公司);NaHCO3:分析纯;Na2HPO4:分析纯;KH2PO4:分析纯;CaCl2·2H2O:分析纯;MnCl2·4H2O:分析纯;NaOH:分析纯;H2SO4:分析纯;甲苯:分析纯;尿素:分析纯;次氯酸钠:分析纯(西陇科学股份有限公司);MgSO4·7H2O:分析纯(天津市福晨化学试剂厂);尿素氮测试盒(C013-1-1,南京建成生物工程研究所有限公司);沸石(200目,江西冠菌生物科技有限公司)。
立式压力蒸汽灭菌器(LDZX-50KBS,上海申安医疗器械厂);气浴恒温振荡摇床(CHA-S,江苏荣华仪器制造有限公司);自动双重水蒸馏器(SZ-93,上海银泽仪器设备有限公司);紫外分光光度计(Q/TBCR1,上海精科实业有限公司);高速冷冻离心机(Sorvall ST 8R,赛默飞世尔科技);微型螺旋混合仪(G-560E,Scientific Industries);电子天平(ATY124,日本岛津);玻璃发酵瓶(100 mL,徐州乾通玻璃制品有限公司);超净工作台(YJ-VS,无锡一净净化设备有限公司);超低温冰箱(DW-60W151EU1,青岛海尔集团)。
1.1.2 试验动物与发酵底物
在江西农业大学附近农场选购10只30周龄、体重(1.5±0.1)kg、采食产蛋正常的白来航蛋鸡用作发酵菌源制备。同时选取农场使用的无抗蛋鸡日粮用作发酵底物,日粮为粉料形式玉米-豆粕型日粮,且符合中华人民共和国农业行业标准《鸡饲养标准》(NY/T 33—2004)中规定的营养标准。日粮组成及营养水平如表1所示。

表1 蛋鸡日粮组成及营养水平
1.2 试验方法
1.2.1 试剂配制
缓冲溶液:NaHCO3105.00 g,NH4HCO312.00 g,用去离子水定容至3 000 mL。常量元素溶液:Na2HPO45.70 g,MgSO4·7H2O 0.60 g,KH2PO46.20 g,用去离子水定容至1 000 mL。微量元素溶液:MnCl2·4H2O 10.00 g,CaCl2·2H2O 13.20 g,FeCl2·6H2O 0.80 g,CoCl2·6H2O 1.00 g,用去离子水定容至100 mL。还原剂溶液:Na2S·9H2O 335.00 mg,1 mol/L NaOH 2 mL,用去离子水定容至50 mL。0.1%刃天青溶液:在100 mL去离子水中溶解100.00 mg刃天青。
1.2.2 菌源获取
根据江西农业大学动物实验伦理要求规定,将试验蛋鸡肌肉注射戊巴比妥钠麻醉后宰杀,立即剖开腹腔,将盲肠近端结扎固定并从结扎外侧剪断。迅速将盲肠取出转移至超净工作台,快速取出所有盲肠内容物并将其混匀,按照1∶3(W∶V)比例加入39 ℃预热的缓冲溶液充分搅匀,经5 层无菌纱布过滤,滤液在39 ℃下持续通入CO2制成体外发酵菌源待用。
1.2.3 试验分组
丁酸钠添加量参考王安平[1]在体外发酵试验中得出的0.15%最佳添加量;沸石添加量参考Fendri 等[10]和Lon-Wo 等[11]的饲喂试验,同时考虑采食和消化过程中的损耗,决定添加1.5%沸石,并且通过预试验验证了添加1.5%的沸石在体外发酵中有减少氨气排放的效果,最终确定沸石添加量为1.5%。试验分为4组:对照组(CT组),单一丁酸钠添加组(A组),单一沸石添加组(B 组)及混合添加组(C 组),每组4 个重复。参考王安平[1]的研究方法:干物质消化率按70%计算折算为采食饲料的量。在各处理的发酵瓶中添加1 000.00 mg 模拟盲肠内容物的粉末,最终折算为添加采食量3 334.00 mg 的日粮作为发酵底物。A组添加采食饲料质量比0.15%(5.00 mg)的丁酸钠;B 组添加采食饲料质量比1.5%(50.01 mg)的沸石;C组则混合添加0.15%(5.00 mg)的丁酸钠和1.5%(50.01 mg)的沸石;CT组不作添加处理。
1.2.4 体外发酵与样品采集
体外发酵产气技术参考王安平[1]的发酵方法。
将缓冲溶液预热至39 ℃,分别量取超纯水474 mL,缓冲溶液237 mL,常量元素溶液237 mL,微量元素溶液0.12 mL,刃天青溶液1.22 mL,混合均匀并持续通入CO210 min,然后加入50 mL还原剂溶液,继续通入CO2至溶液无色,制得1 000 mL接种液。将提前制备好的菌源与接种液按体积比1∶2混匀,制得体外发酵液。
在已加入发酵底物的发酵瓶中加入60 mL 上述体外发酵液,通入CO2排尽瓶中空气后密封。随后将密封好的发酵瓶放置于39 ℃的气浴恒温振荡摇床中,振荡培养12 h。发酵结束,将发酵瓶立即置于冰水终止发酵。将发酵瓶中的气体抽取干净并通过H2SO4溶液吸收固定氨气,另外采集10 mL 发酵液保存于超低温冰箱用于后续分析,取样2 mL 发酵液于液氮中保存,进行16S rDNA基因高通量测序工作。
1.2.5 样品指标测定
1.2.5.1 氨气产量测定
采用纳氏比色法,取1 mL硫酸吸收液于试管中,稀释至25 mL后加入酒石酸钾钠1 mL,再加入0.5 mL纳氏试剂,混匀后静置10 min,使用紫外分光光度计420 nm 处测定吸光值。绘制标准曲线后计算各样品氨气产量。
1.2.5.2 尿素含量测定
采用南京建成生物工程研究所有限公司的尿素氮测试盒(二乙酰肟比色法),具体操作参照试剂盒说明书。
1.2.5.3 脲酶活性测定
采用靛酚蓝比色法,取1 mL离心上清液加入1 mL甲苯,静置15 min 后加入10%尿素溶液10 mL、柠檬酸缓冲溶液20 mL。混匀后置于37 ℃气浴恒温振荡摇床中培养24 h。培养结束取3 mL过滤后的培养液于50 mL 容量瓶中,加入17 mL 超纯水、3 mL 次氯酸钠、20 mL苯酚钠,混匀后静置20 min显色,定容。使用紫外分光光度计于578 nm 处测定吸光值,根据回归方程计算脲酶活性。脲酶活性用1 mL发酵液24 h降解尿素生成铵态氮的量表示。
1.2.5.4 16S rDNA分析
DNA 的提取与PCR 扩增:使用FastDNA®Spin Kit for Soil DNA提取试剂盒,完成DNA提取。PCR扩增区域为338F_806R,引物对应16S rDNA 的V3~V4区,上游引物338F:3'-ACTCCTACGGGAGGCAGCAG-5',下游引物806R:3'-GGACTACHVGGGTWTCTAAT-5'。完成预试验后,PCR 正式试验采用TransGen AP221-02:TransStart Fastpfu DNA Polymerase,20 μL反应体系。每个样本3个PCR重复,将3个重复的PCR产物混合;使用2%琼脂糖凝胶电泳检测产物。使用AxyPrep DNA 凝胶提取试剂盒回收PCR 产物,将PCR 产物用Quantus™Fluorometer 进行检测定量。按照每个样本的测序量要求进行相应比例的混合。
构建Miseq 文库与测序:使用NEXTFLEX Rapid DNA-Seq Kit进行建库:①接头链接;②使用磁珠筛选去除接头自连片段;③利用PCR扩增进行文库模板的富集;④磁珠回收PCR产物得到最终的文库。利用Illumina 公司的Miseq PE300/NovaSeq PE250 平台进行测序。按照平台说明进行操作:①DNA片段的一端与引物碱基互补,固定在芯片上;②另一端随机与附近的另外一个引物互补,也被固定住,形成“桥”;③扩增,产生DNA 簇;④DNA 扩增子线性化成为单链;⑤加入改造过的DNA 聚合酶和带有4 种荧光标记的dNTP,每次循环只合成一个碱基;⑥用激光扫描反应板表面,读取每条模板序列第一轮反应所聚合上去的核苷酸种类;⑦将“荧光基团”和“终止基团”化学切割,恢复3′端黏性,继续聚合第二个核苷酸;⑧统计每轮收集到的荧光信号结果,获知模板DNA 片段的序列。使用FLASH 软件对测序序列进行拼接,同时使用fastp 软件进行质量控制,随后使用UPARSE 软件,根据97%的相似度对序列进行OTU聚类。
生物多样性分析:将相似性大于97%的序列定义为一个OTU,每个OTU 对应不同的微生物物种。对16个样本在OTU分类水平进行PCA分析以及相关性分析。同时对各组样本的有效序列分别在门水平和属水平上进行微生物群落分析。
1.3 统计分析
采用Origin 9.0软件中的“ANOVA andt-test”进行差异显著性分析,以P<0.05作为差异显著的判断标准。
2 结果与分析
2.1 不同处理对蛋鸡盲肠微生物体外发酵氨气产量的影响
由图1 可知,CT 组的氨气产量最高为(14.73±0.42)μg,显著高于A、B、C组(P<0.05);C组的氨气产量最低为(10.84±0.11)μg,显著低于另外3组(P<0.05);A组氨气产量大于B组,但两组间差异不显著(P>0.05)。
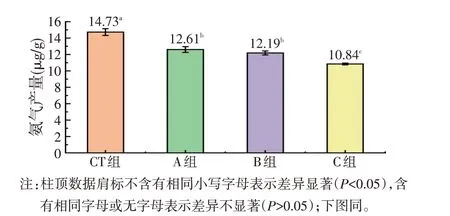
图1 不同处理对蛋鸡盲肠内容物体外发酵氨气产量的影响
2.2 不同处理对蛋鸡盲肠内容物体外发酵脲酶活性的影响
图2 表明,CT 组的脲酶活性最高为(0.68±0.01)mg/mL·24 h,显著高于A、B、C组(P<0.05);C组的脲酶活性最低为(0.45±0.03)mg/mL·24 h,显著低于另外3 组(P<0.05);A 组与B 组间脲酶活性差异不显著(P>0.05)。
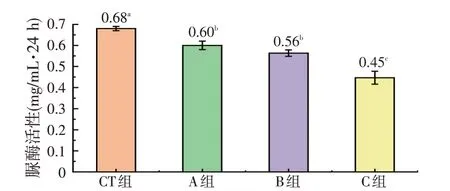
图2 不同处理对蛋鸡盲肠内容物体外发酵脲酶活性的影响
2.3 不同处理对蛋鸡盲肠内容物体外发酵尿素产量的影响
如图3 所示,C 组发酵液中的尿素含量最高为(0.175±0.010)mmol/L,显著高于其他3 组(P<0.05);CT、A组和B组间的差异不显著(P>0.05)。

图3 不同处理对蛋鸡盲肠内容物体外发酵尿素含量的影响
2.4 不同处理对蛋鸡盲肠内容物体外发酵液中微生物群落的影响
2.4.1 样品信息统计
如表2所示,本试验共获取575 267条有效序列,在检测的16个样本中共检出822个微生物OTU,涵盖16门、26纲、50目、90科、199属、366种。

表2 样本检测信息统计
2.4.2 样本OTU的主成分分析
由图4可知,CT组与3个处理组均分开,3个处理组之间也互相分开没有聚集,说明不同的添加处理改变了发酵液中的微生物组成。
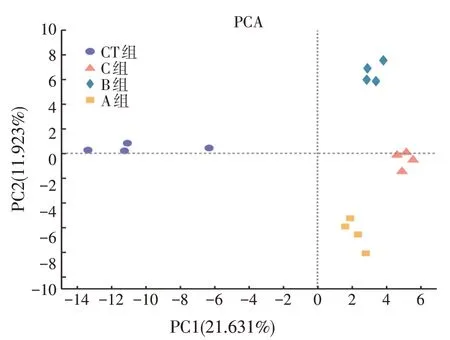
图4 不同处理蛋鸡盲肠内容物体外发酵液的PCA分析
2.4.3 样本的相关性分析
由图5 可知,热图中各处理的聚类较为规律,CT组独自聚类在一起,而C 组也聚类在一起,与OTU 的主成分分析的结果相同,反映了不同的添加处理会影响到发酵液的微生物组成。
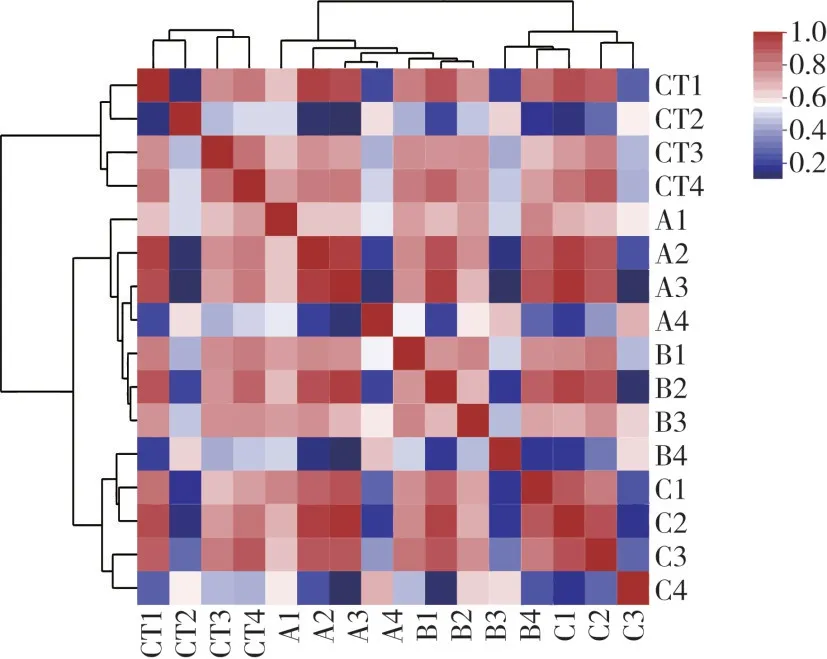
图5 不同处理蛋鸡盲肠内容物体外发酵液的相关性热图
2.4.4 门(Phylum)分类水平上的细菌菌落分析
将所有样本的有效序列在门分类水平进行细菌群落分析,组内样本计算平均值。由图6和表3可知,在A、B、C 组中,拟杆菌门是优势菌门,相对丰度分别为40.8%、39.3%及41.0%;A、C组的拟杆菌门丰度显著大于B组(P<0.05);而CT组拟杆菌门相对丰度只有37.0%显著低于A、B、C组(P<0.05)。CT组的优势菌门是厚壁菌门,相对丰度为39.1%,显著大于A、B、C组(P<0.05),同时A组和B组在厚壁菌门的相对丰度上差异不显著(P>0.05),C组的厚壁菌门相对丰度在所有分组中最低。

图6 不同处理蛋鸡盲肠内容物体外发酵液门水平上的细菌群落结构分析
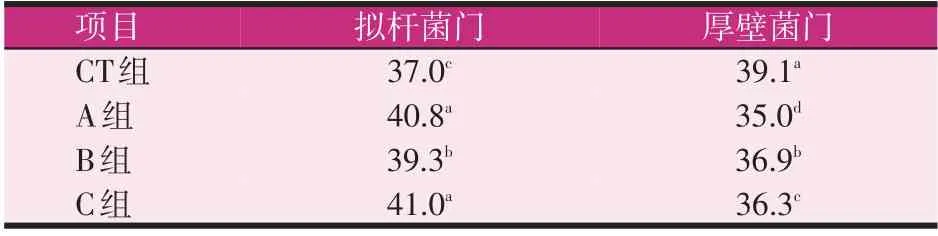
表3 门水平主要细菌结构变化(%)
2.4.5 属(Genus)分类水平上的细菌菌落分析
由表4和图7可知,在属分类水平上,4个组的优势菌属都是拟杆菌属,但A、B、C组的拟杆菌属相对丰度均显著大于CT 组(P<0.05);其中C 组的拟杆菌属相对丰度为20.6%,显著大于其他组(P<0.05)。脱硫弧杆菌属在CT组中的相对丰度最大为3.87%,显著大于其他3 组(P<0.05)。乳酸杆菌属方面,C 组的相对丰度最大为4.24%,显著大于其他3组(P<0.05),CT组相对丰度最低(1.76%),显著低于A、B组(P<0.05)。

表4 属水平主要细菌结构变化(%)

图7 不同处理蛋鸡盲肠内容物体外发酵液属水平上的细菌群落结构分析
3讨论
本试验研究结果表明,添加0.15%丁酸钠可减少氨气释放(P<0.05),其原因可能为:在蛋鸡体内的盲肠发酵阶段,氨气的产生途径主要来自尿酸的降解或盲肠微生物的分解代谢[1];其中盲肠中的尿酸是氨气产生的主要底物[16]。尿酸降解包含了一系列的酶促反应,其中尿酸酶和脲酶是该过程的限速酶[17]。在蛋鸡盲肠内容物体外发酵时添加丁酸钠可以有效抑制尿酸酶与脲酶活性,限制尿酸降解,进而减少氨气产生,王安平[1]的研究中有类似报道。
本试验研究还发现,1.5%沸石的添加也可减少氨气释放(P<0.05),罗一鸣等[12]有相似研究结果,他们发现在鸡粪堆肥时添加沸石可以吸附固定氨态氮达到减少氨气挥发的效果。除此以外,沸石具有比表面积大、吸附性能好、表面呈负电性、有利于络合吸附重金属离子等特性[18-19];而脲酶的活化与催化均需要镍离子的参与[20],沸石可吸附固定镍离子,进而有效抑制脲酶活性,减少氨气的产生。
值得注意的是,相对于其他所有组,0.15%丁酸钠和1.5%沸石的混合添加组在减少氨气排放上效果最佳,与对照组(脲酶活性0.68 mg/mL·24 h)相比,混合添加处理组对脲酶活性的抑制率达到了33.82%。基于此,推测丁酸钠与沸石可能存在协同作用,与单一添加丁酸钠或沸石相比,能进一步降低脲酶活性,抑制尿酸—尿素—氨气的分解过程,减少氨气产生。通过脲酶活性与尿素含量检测结果表明,混合添加丁酸钠和沸石可增强脲酶的抑制效果。而其协同作用发生的原因可能与沸石通过分子间作用力将正丁酸吸附到其表面[21]相类似,沸石吸附丁酸钠进而使其表面的负电性增大,增强了对镍离子的吸附作用,使镍离子络合固化增多而进一步抑制了脲酶活性。
本研究中样本的主成分分析(图4)与相关性分析(图5)的结果均表明,三种不同的添加处理均可以影响体外发酵体系的微生物群落结构。王安平[1]有相似的探究结果,其研究发现在蛋鸡盲肠内容物发酵时,微生物组成会随丁酸钠添加量的改变而变化。另外有研究表明,在日粮中添加沸石对肉鸡盲肠微生物结构有一定影响[22]。上述研究与本研究结果基本相符。在3个添加处理组的发酵液样品中,拟杆菌门与拟杆菌属是优势菌。Zocco等[23]研究指出,拟杆菌属以碳水化合物作为主要的能量来源,对氨基酸的分解利用较少,因此产氨较少。有研究中提到丁酸钠可提高部分碳水化合物发酵菌的相对丰度,丁酸钠干预后产氨能力弱的菌属相对丰度增加,产氨能力强的菌属相对丰度下降[1]。
强产氨菌属相对丰度的下降可能是自身合成的脲酶活性被抑制,无法分解产生氨气来维持其生存环境平稳所致。Ricci等[24]曾报道过幽门螺旋杆菌分解代谢产生氨气并利用其提高肠道pH 来存活。Kuever 等[25]报道脱硫弧杆菌属在代谢时可以将硝酸盐还原成氨。本研究结果表明,脱硫弧杆菌相对丰度随着添加处理的不同而改变,其中混合添加组的相对丰度最低。很可能是因为混合添加组对脲酶活性存在更强的抑制作用,最终导致其相对丰度下降。这与上述的研究推测相符。当大部分产氨的有害菌增殖被抑制后,部分有益菌的相对丰度会出现上升。周岩民等[26]研究证明了不同粒度的沸石均有降低肉鸡空肠中大肠杆菌数、提高乳酸杆菌数的效果。而郭传珍等[27]也曾报道了饲粮中混合100 g/t的丁酸钠,在肉鸡42日龄时可以显著提高乳酸杆菌数量,抑制大肠杆菌的繁殖。本研究结果与上述研究基本相符,并且混合添加组对乳酸杆菌属的提高效果最好,侧面表明混合添加组对产氨有害菌的抑制效果更强,且其原因也极有可能是丁酸钠与沸石之间发生协同作用,使沸石表面对镍离子等的吸附作用增强,进而抑制脲酶活性所致。当然,对于其是否如此,需要进一步对丁酸钠与沸石做电镜观察研究。
4 结论
本研究通过体外发酵试验对混合添加丁酸钠与沸石减氨排放的可行性进行了验证。试验结果表明,在体外发酵体系内混合添加丁酸钠和沸石可以显著抑制脲酶活性进而显著减少氨气产生,丁酸钠与沸石在抑制脲酶活性减少氨排放上可能存在协同作用。此外混合添加处理还对发酵体系中的微生物群落结构造成影响,产氨的脱硫弧杆菌属的增殖被抑制,而以拟杆菌属为主的碳水化合物发酵菌相对丰度上升成为优势菌属;乳酸杆菌属的相对丰度也有明显上升。
猜你喜欢
煤气与热力(2021年9期)2021-11-06
化学工业与工程(2021年5期)2021-11-03
湖南饲料(2021年3期)2021-07-28
生物技术通报(2020年12期)2020-12-21
腐植酸(2020年1期)2020-11-29
渔业现代化(2019年3期)2019-07-11
天然气化工—C1化学与化工(2019年6期)2019-02-18
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07
植物营养与肥料学报(2011年3期)2011-10-24
