
不同乳酸菌发酵甘薯渣产物组分分析
2023-01-14 07:59张丽君木泰华马梦梅
核农学报 2023年1期
张丽君 木泰华 马梦梅
(1青岛农业大学食品科学与工程学院,山东 青岛 266109;2中国农业科学院农产品加工研究所,北京 100193;3青岛特种食品研究院,山东 青岛 266109)
我国甘薯资源丰富,种植面积和年产量分别为232万公顷和5 126.4万吨,均居世界首位[1]。现阶段,我国甘薯主要用于加工淀粉及其制品(粉丝、粉条),在此过程中会产生大量的甘薯渣。据统计,每生产1 吨淀粉可产生6.5~7.5 吨湿薯渣[2-3]。然而,目前只有极少的甘薯渣被用作廉价动物饲料,大部分被作为废弃物直接丢弃,造成严重的资源浪费和环境污染[4]。前期研究表明,甘薯渣中依然存在大量的淀粉以及丰富的膳食纤维,具有很高的利用价值[5]。因此,亟需以甘薯渣为原料开发新型产品,延长甘薯加工产业链,提高甘薯附加值。
乳酸菌是一类无芽孢、革兰氏染色阳性细菌的总称,因能够利用葡萄糖或乳糖进行发酵产生乳酸而得名[6]。研究发现,乳酸菌具有维持人体肠道菌群微生态平衡、促进肠道吸收营养物质、增强机体免疫力、降低血清胆固醇等作用[7]。近年来,随着乳酸菌生理活性得到广泛认知,其在食品领域的应用也从乳制品逐渐扩展到植物性原料领域[8]。例如,杨立启等[9]比较了植物乳杆菌、嗜酸乳杆菌和副干酪乳杆菌在柑橘全果汁中的发酵性能,结果表明,与未发酵果汁相比,植物乳杆菌发酵的蜜桔果汁中黄酮和多酚含量更高;Wu 等[10]研究显示,植物乳杆菌、嗜热链球菌发酵可提高蓝莓汁中的总酚含量与抗氧化活性;马建功等[11]采用植物乳杆菌和鼠李糖乳杆菌发酵小米浆,结果显示发酵后小米浆的总酚、还原糖、氨基酸态氮、矿物质元素含量均显著上升;Weng 等[12]利用副干酪乳杆菌发酵豆渣,结果显示发酵后产物的可溶性膳食纤维、有机酸含量显著提高,风味物质显著增多;Pejin 等[13]以啤酒厂废谷物、麦芽和豆粕为原料,利用鼠李糖乳杆菌进行发酵,评估发现发酵产物中的乳酸、氨基酸及矿物质元素含量显著升高。由此可见,乳酸菌发酵可显著提高果蔬、谷物、豆类及其加工副产物中营养与功能成分的含量及生理活性,并改善口感和风味。然而,微生物的生长和发酵能力不仅与菌种本身有关,还与发酵基质的成分及发酵条件有关,目前尚鲜见不同乳酸菌应用于甘薯渣的报道。
本研究以植物乳杆菌、干酪乳杆菌、保加利亚乳杆菌、戊糖片球菌、嗜热链球菌、商业植物乳杆菌分别对甘薯渣进行发酵,收集发酵后的上清液,并对其理化特性(pH 值、总酸含量)及营养功能成分(乳酸及短链脂肪酸、总膳食纤维、可溶性膳食纤维、不溶性膳食纤维、蛋白质、灰分、还原糖及总糖、总酚、游离氨基酸、矿物质元素含量)进行系统分析,采用灰色理论加权关联度对不同乳酸菌发酵甘薯渣产物的综合营养价值进行评价,旨在为基于乳酸菌发酵的新型甘薯饮品设计提供基础数据和理论支撑。
1 材料与方法
1.1 材料与试剂
新鲜甘薯(济薯26),市售;耐高温α-淀粉酶(酶活:40 000 U·g-1)、碱性蛋白酶(酶活:105U·mL-1),北京萃锋科技有限公司;乳酸菌菌种:植物乳杆菌(BNCC 195072)、干酪乳杆菌(BNCC 134415)、保加利亚乳杆菌(BNCC 336436)、嗜热链球菌(BNCC 186558),北京绿源伯德生物科技有限公司;戊糖片球菌(CICC 21862),中国微生物菌种保藏中心;商业植物乳杆菌,山东中科嘉艺生物工程有限公司。
食品级柠檬酸、食品级氢氧化钠,广州焙考林食品有限公司;氨基酸分析标准品、乳酸标准品,美国Sigma公司;挥发性脂肪酸标准品,北京同源科创科技有限公司;矿物质元素标准品,北京百灵威科技有限公司;葡萄糖标准品、福林酚试剂,北京索莱宝科技有限公司;MRS 肉汤(MRS broth)、3,5-二硝基水杨酸(3,5-dinitrosalicylic acid,DNS)试剂、甲醇(色谱纯)、无水乙醇(色谱纯)、氢氧化钠(分析纯)、无水碳酸钠(分析纯),北京萃锋科技有限公司;盐酸、硝酸、丙酮、硫酸(分析纯),北京国药集团有限公司。
1.2 仪器与设备
BPG-9240A 精密鼓风干燥箱,北京天林恒泰科技有限公司;SH2-300A 水浴摇床,苏州培英实验设备有限公司;LDZW-40KCS 立式压力蒸汽灭菌锅,上海申安医疗器械厂;恒温培养箱,太仓市华利达实验设备有限公司;SW-CJ-1FD 超净工作台,苏州安泰空气技术有限公司;ST3100 pH 计,上海奥豪斯仪器有限公司;UV1101 分光光度计、L-8900 全自动氨基酸分析仪,日本日立公司;KJELTEC 2300 自动凯氏定氮仪,丹麦FOSS 公司;MARS 5 微波消解仪、ICP-MS7700 电感耦合等离子体质谱仪、LC-MS1100 液相色谱-质谱联用仪,美国安捷伦公司;ICS-3000 离子色谱仪、Lindberg/Bliue马弗炉,美国赛默飞世尔科技公司。
1.3 试验方法
1.3.1 甘薯渣乳酸菌发酵产物的制备 菌种活化:在无菌条件下,分别取植物乳杆菌(Lactobacillus plantarum,Lp)、干酪乳杆菌(Lactobacillus casei,Lc)、保加利亚乳杆菌(Lactobacillus bulgaricus,Lb)、戊糖片球菌(Pediococcus pentosus,Pp)、嗜热链球菌(Streptococcus thermophilus,St)冻干粉接种在MRS 肉汤培养基中,37 ℃活化培养48 h。
原料预处理:将新鲜甘薯清洗、去皮、分切,与水按1∶1(w/w)进行打浆,离心、干燥(65 ℃)后得甘薯渣。将甘薯渣与水按1∶10(w/v)混合均匀后,加入0.5%耐高温α-淀粉酶,95 ℃条件下酶解30 min,冷却调节pH 值至7.8±0.2,加入0.1%碱性蛋白酶,60 ℃下酶解30 min,调节pH值至6.0±0.2,在121 ℃下高温灭菌15 min,冷却至37 ℃。
接种及发酵:在无菌条件下,将活化好的植物乳杆菌(Lp)、干酪乳杆菌(Lc)、保加利亚乳杆菌(Lb)、戊糖片球菌(Pp)、嗜热链球菌(St)、商业植物乳杆菌(Shang Zhi,SZ)菌悬液按1%(v/v,8 lg CFU·mL-1)分别接入甘薯渣中,混匀,恒温培养箱中37 ℃发酵36 h 后高温灭菌停止发酵,冷却后7 000 r·min-1离心15 min,取上清液。
1.3.2 理化特性测定
1.3.2.1 pH 值的测定 取10 mL 发酵液,玻璃棒搅拌均匀,直接用pH计测定。
1.3.2.2 总酸的测定 按照《GB 12456-2021 食品安全国家标准食品中总酸的测定》[14]测定(以乳酸计)。
1.3.3 营养功能成分测定
1.3.3.1 乳酸的测定 采用液相色谱-质谱联用仪(liquid chromatography - mass spectrometry,LC-MS),参考于勇等[15]的方法进行测定,并稍作修改。色谱条件:色谱柱为Waters T3(150 mm×2.1 mm,5 m);流动相A:0.1%甲酸水,流动相B:甲醇;载气温度:350 ℃,鞘气温度:300 ℃,流速:0.3 mL·min-1;进样量:1 µL。质谱条件:采用气动辅助电喷雾离子化方式电离,检测方式为负离子多反应监测,扫描段:预离子89;碎片离子45、43、42;裂解电压为73 V;碰撞电压10 eV。样品处理:取1 mL 发酵后的上清液至100 mL 容量瓶中,用超纯水混匀、定容,静置30 min,取1.2 mL 上清液,在4 ℃、14 000 r·min-1条件下离心10 min,过0.22µm滤膜,供LC-MS测定。
1.3.3.2 短链脂肪酸的测定 采用气相色谱法,参考包娜然等[16]的方法进行测定,并稍作修改。色谱柱为DB-FFAP(30 m×0.53 mm×0.5 µm,Agilent)。采用FID 检测器(280 ℃),分流比30∶1,进样量1 µL。色谱柱程序升温:初始温度70 ℃保持2 min,然后以10 ℃·min-1升温至115 ℃保持9 min,再以45 ℃·min-1升温至200 ℃保持2 min。载气:高纯氦气2 mL·min-1。标准溶液制备:取1 mL短链脂肪酸混合(100µg·mL-1)标准贮备液于离心管中,加入100 µL 25%偏磷酸溶液,充分混合,过0.22 µm 滤膜注入进样小瓶,用于绘制标准曲线。样品前处理:取1.0 mL 样品溶液至1.5 mL 离心管中,加入100µL 25%偏磷酸溶液,混匀,静置1 h 后,在4 ℃、14 000 r·min-1条件下离心10 min,将上清液过0.22µm滤膜注入进样小瓶,上机检测。
1.3.3.3 基本成分的测定 总膳食纤维、可溶性膳食纤维、不溶性膳食纤维含量按照AOAC Method 991.43[17]测定;蛋白质含量按照AOAC Method 955.04[18]测定;灰分含量按照AOAC Method 942.05[19]测定。
1.3.3.4 总糖和还原糖含量测定 参考赵凯等[20]的方法并稍作修改,采用3,5-二硝基水杨酸法测定。吸取1 mL样品溶液,稀释至100倍,吸取1 mL稀释样品溶液于试管中,加入2 mL DNS试剂,沸水浴显色3 min,在540 nm 处测定其吸光值,还原糖含量以葡萄糖含量计(葡萄糖标准曲线的制作:配置葡萄糖标准溶液0.20、0.25、0.30、0.35、0.40、0.45 mg·mL-1,线性方程为:y=2.479 7x-0.036 0,R2=0.993)。
1.3.3.5 总酚含量测定 参考刘聪[21]的方法并稍作修改,采用福林酚比色法测定。取1 mL 稀释10 倍的样品溶液,加入2 mL福林酚试剂混合,加入2 mL 10%的Na2CO3溶液,摇匀,静置避光1 h后,在700 nm处测定其吸光值,总酚含量以没食子酸当量计(没食子酸标准曲线的制作:配置质量浓度为5、10、15、20、25 mg·mL-1的没食子酸标准溶液,线性方程为y=0.010 6x+0.002 4,R2=0.999 8)。
1.3.3.6 游离氨基酸含量测定 参考姜荷等[22]的方法,采用全自动氨基酸分析仪进行测定。将1 mL样品与1 mL 8%磺基水杨酸混合,10 000 r·min-1离心15 min,吸取1 mL 上清液氮吹至干燥,加入1 mL 0.02 mol·L-1HCl溶液复溶,过0.22µm水相滤膜,上机测试。
1.3.3.7 矿物质元素含量测定 参考李承范等[23]的方法并稍作修改,采用电感耦合等离子体质谱(inductively coupled plasma mass spectrometry,ICP-MS)检测。准确称取样品0.25 g(精确至0.000 1)于微波消解罐中,加入浓硝酸6 mL 预消解1 h,加入过氧化氢2 mL,上机进行微波消解,消解完成后转移至100 mL 容量瓶中,超纯水定容,摇匀,测定各矿物质元素的含量。
1.3.4 灰色理论加权关联度分析 根据乳酸菌发酵甘薯渣产物的理化特性及营养功能成分的测定结果筛选“理想菌种”,确定正负相关营养指标,正相关指标:总酸含量、乳酸及短链脂肪酸、蛋白质、灰分、总酚、总膳食纤维、可溶性膳食纤维、游离必需氨基酸/游离氨基酸总量(free essential amino acids/total free amino acids,FEAA/TFAA)、矿物质元素;负相关指标:pH 值、不溶性膳食纤维、还原糖、总糖,结合正相关指标上限和负相关下限建立最优菌种并构建“理想菌种”;“归一化”处理:对于正相关指标,归一化值以所有样品各指标占理想菌种对应指标的比值表示,负相关指标归一化值则以所有样品各指标占理想菌种对应指标比值的倒数表示,最终将所有指标转化为(0,1)之间的相关数列;灰色理论关联度分析:把发酵甘薯渣产物的理化指标及营养功能成分归一化得到的数列分别看做一个灰色系统,根据灰色理论公式分析计算[19];将每个样品得到的关联度结果用Microsoft Excel 2019软件按照Hu等[24]的方法制作成热图,以便了解每个指标对最终结果的贡献程度。最后将得到的样品加权关联度按照从高到低进行排序,即得到不同菌种发酵甘薯渣产物的综合功能营养价值综合排序。
1.4 数据处理
所有试验重复测定3次,结果以平均值±标准差表示。采用SPSS 22.0 软件进行显著性分析,P<0.05 表示差异显著。
2 结果与分析
2.1 不同乳酸菌发酵甘薯渣产物的理化特性
pH 值和总酸含量是乳酸菌发酵过程中用来评判发酵活性的重要指标之一,常用来表征乳酸菌的产酸能力,某些情况下用来反映其生长代谢情况[25]。由表1可知,未发酵甘薯渣的pH 值为5.70、总酸含量为0.27 g·100 mL-1。采用6 种乳酸菌进行发酵后,甘薯渣发酵产物的pH 值和总酸含量均发生了显著变化,说明6 种菌株产酸能力有显著差异,其中Lb发酵甘薯渣产物的pH值最低,为3.15;Lc 发酵甘薯渣产物的总酸含量最高,为2.79 g·100 mL-1。

表1 不同乳酸菌发酵甘薯渣产物的pH值和总酸含量Table 1 pH value and total acidity of sweet potato residue fermented by different lactic acid bacteria
2.2 不同乳酸菌发酵甘薯渣产物的营养功能成分
2.2.1 不同乳酸菌发酵甘薯渣产物的乳酸及短链脂肪酸含量 乳酸及短链脂肪酸主要是乳酸菌利用底物中的葡萄糖、膳食纤维、低聚糖等发酵产生的代谢物[26]。由表2可知,与对照相比,乳酸菌发酵甘薯渣产物的乳酸、乙酸、丙酸含量升高,且不同菌种发酵产物的乳酸及短链脂肪酸含量整体差异显著(P<0.05),表明不同菌种利用碳源进行发酵的能力显著不同[27],其中Lb组甘薯渣发酵产物中乳酸含量最高,为11.60 mg·mL-1;SZ组甘薯渣发酵产物中乙酸含量最高,为66.99µg·mL-1;Lc组甘薯渣发酵产物中丙酸含量最高,为4.41µg·mL-1。

表2 不同乳酸菌发酵甘薯渣产物的乳酸及短链脂肪酸含量Table 2 Lactic acid and short chain fatty acids of sweet potato residue fermented by different lactic acid bacteria
2.2.2 不同乳酸菌发酵甘薯渣产物的基础成分含量 乳酸菌发酵期间会产生丰富的酶类物质,如纤维素酶、半纤维素酶、蛋白酶等,这些酶类物质可将发酵基质中的大分子物质水解为小分子物质,进而提高发酵产物的营养价值和生理活性[28]。由表3可知,不同乳酸菌均可不同程度地提高甘薯渣发酵产物中可溶性膳食纤维、总膳食纤维含量,显著提高蛋白质及灰分含量(P<0.05),降低不溶性膳食纤维的含量。其中,以Lb 组发酵产物中总膳食纤维、可溶性膳食纤维和蛋白质含量最高,分别是对照组的1.64、2.00和3.42倍
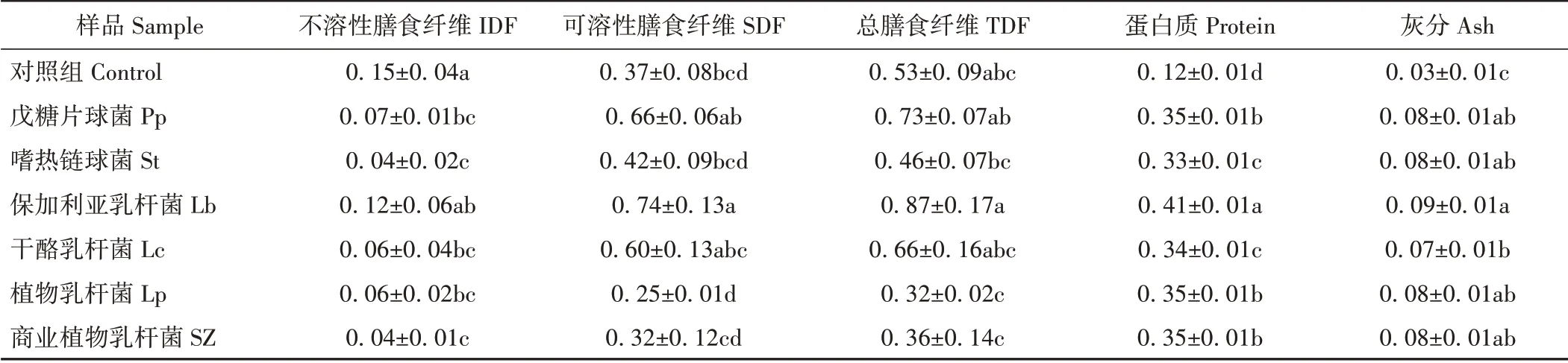
表3 不同乳酸菌发酵甘薯渣产物的基础成分含量Table 3 The basic components content of sweet potato residue fermented by different lactic acid bacteria /(g·100 mL-1)
2.2.3 不同乳酸菌发酵甘薯渣产物的总糖和还原糖含量 乳酸菌可利用单糖和低聚糖进行糖代谢,因此甘薯渣发酵前采用耐高温α-淀粉酶进行酶解,产生丰富的短链糊精、低聚糖和单糖,可为乳酸菌发酵提供必需的碳源[29],导致酶解液(E组)中总糖及还原糖含量显著提高。由图1-A可知,与对照及酶解组相比,除St 组外,其余5种乳酸菌均可提高甘薯渣发酵产物中的总糖含量,其中Pp组总糖含量最高,为58.66 mg·mL-1。由图1-B可知,在对照组和酶解组中,甘薯渣中还原糖含量分别为8.13 和29.79 mg·mL-1,经过乳酸菌发酵后,除SZ 组外,其余发酵组中的还原糖含量较酶解组显著下降(P<0.05),其中Pp 组还原糖含量最低,为18.67 mg·mL-1。

图1 不同乳酸菌发酵甘薯渣产物的总糖(A)和还原糖(B)含量Fig.1 Total sugar(A)and reducing sugar(B)of sweet potato residue fermented by different lactic acid bacteria
2.2.4 不同乳酸菌发酵甘薯渣产物的总酚含量 由图2可知,与对照组相比,不同乳酸菌均可提高甘薯渣发酵产物中的总酚含量,其中以St 组中总酚含量最高,为146.87µg GAE·mL-1,其次为Lp组,总酚含量为134.61µg GAE·mL-1。

图2 不同乳酸菌发酵甘薯渣产物的总酚含量Fig.2 Total polyphenol content of sweet potato residue fermented by different lactic acid bacteria
2.2.5 不同乳酸菌发酵甘薯渣产物的游离氨基酸含量 由表4可知,6 种乳酸菌发酵产物和对照组中检测出16 种游离氨基酸,但其含量存在明显差异。与对照组相比,发酵产物中的游离氨基酸总量(FTAA)和游离必需氨基酸含量(FEAA)均有不同程度的增加,分别提高了0.80~3.71倍和1.45~2.00倍,其中Lc组含量最高,分别为53.61 mg·100 mL-1和9.54 mg·100 mL-1。游离氨基酸总量变化的主要贡献者是谷氨酸,游离必需氨基酸含量变化的主要贡献者为缬氨酸、蛋氨酸、苯丙氨酸和亮氨酸。

表4 不同乳酸菌发酵甘薯渣产物的游离氨基酸含量Table 4 Free amino acids content of sweet potato residue fermented by different lactic acid bacteria /(mg·100 mL-1)
2.2.6 不同乳酸菌发酵甘薯渣产物的矿物质元素含量 由表5可知,甘薯渣经过发酵后,其产物富含Na、Mg、K、Ca等常量元素以及Fe、Zn、Se等微量元素,其中Na 含量较对照提高了0.80~9.00 倍、Mg 含量提高了0.80~1.40 倍、Ca 含量提高了0.90~2.00 倍、Fe 含量提高了0.95~3.50倍、Zn含量提高了0.73~1.45倍,Lc组的Na、Fe、Zn 元素含量最高、Lp 组的Mg 元素含量最高、SZ 组的Ca 和Se 元素含量最高。上述结果表明甘薯渣经发酵后,可显著提高Mg、Ca、Fe、Zn 等对人体有益的矿物质含量,尤其以Lc组总矿物质元素含量最高。

表5 不同乳酸菌发酵甘薯渣产物的矿物质元素含量Table 5 The mineral elements of sweet potato residue fermented by different lactic acid bacteria
2.3 不同乳酸菌发酵甘薯渣产物的综合营养评价灰色关联度排名
由表6可知,不同乳酸菌发酵甘薯渣产物的灰色关联度评分为0.701~0.823,从排名结果来看,保加利亚乳杆菌和干酪乳杆菌发酵甘薯渣产物综合营养价值较高。热图分析结果(图3)可以更直观地反映每个指标对最终结果的贡献程度,从而解释样品加权关联度之间的差异。排名第一的保加利亚乳杆菌和排名第二的干酪乳杆菌具有更多的绿色区域,保加利亚乳杆菌具有优势的指标主要包括乙酸、乳酸、pH 值、可溶性膳食纤维、总膳食纤维、蛋白质、灰分、EAA/TAA、K,说明该菌种与理想菌种更接近;而排名最后的戊糖片球菌发酵产物中具有更多的红色区域,表明该菌种的大部分指标在综合排序中不占优势,是该菌种最终排名较低的原因。

表6 不同乳酸菌发酵甘薯渣产物的综合营养评价灰色关联度排名表Table 6 Grey correlation ranking of comprehensive nutritional evaluation of sweet potato residue fermented by different lactic acid bacteria
3 讨论
甘薯渣经酶解、6种乳酸菌发酵后,所得产物的pH值降低、总酸含量升高,以Lb 组的pH 值最低,Lc 组总酸含量最高(表1)。进一步分析显示,发酵产物中的酸类物质以乳酸、乙酸为主,其中Lb组乳酸含量最高,SZ、Lb 和Lc 组乙酸含量无显著差异(表2),该结果与Weng 等[12]、赵昕琪[30]的研究结果一致。这是因为乳酸菌可利用底物中的小分子糖(如葡萄糖等)为碳源进行发酵,且不同种类乳酸菌的发酵能力存在显著差异,这也解释了发酵产物中还原糖含量下降的现象。同时,发酵产物中可溶性膳食纤维等营养成分含量较对照显著增加,以Lb 组最高,Pp 组、Lc 组次之,是因为乳酸菌发酵过程中产生有机酸等代谢产物,使甘薯渣中不溶性膳食纤维的糖苷键断裂,部分转化为可溶性膳食纤维[31]。
乳酸菌在发酵过程中可利用甘薯渣中的蛋白质及水解产物作为氮源,生成低分子肽及氨基酸等,因此,6 种甘薯渣发酵产物中蛋白质和游离氨基酸含量整体显著提高,这与刘秋豆等[32]的研究结果一致。究其原因,可能是乳酸菌发酵可使蛋白质由结合态转化为游离态释放到上清液中。此外,不同发酵甘薯渣产物的氨基酸组成存在明显不同,这可能是由于不同乳酸菌具有不同的转氨酶活性,使得氨基酸之间的转化不同[33]。
甘薯渣发酵产物中总酚含量较对照显著提高,以St组含量最高(图2),这是因为酚类物质被碳水化合物、蛋白质和脂质等大分子包裹,经过酶和微生物发酵的去酯化、水解等作用转换为游离态释放到液体中[34-35]。在发酵过程中,部分酚类物质与矿物质的螯合作用被微生物的酶类体系解除,乳酸菌也会发生细胞自溶,释放细胞质成分,导致矿物质元素含量增多[36-37];而经过发酵后P元素含量呈现下降的趋势,主要是因为乳酸菌对有机磷具有降解作用[38]。
综上,采用灰色理论加权关联度对6 种乳酸菌发酵甘薯渣产物的理化特性及营养功能成分进行归一化处理与分析,筛选出保加利亚乳杆菌和干酪乳杆菌发酵甘薯渣产物的综合营养价值较高,后续可进一步深入研究上述两种乳酸菌复配应用于甘薯渣发酵的加工工艺及其产物的功能活性。
4 结论
本试验系统研究了不同种类乳酸菌对甘薯渣发酵产物的pH 值、总酸含量、乳酸及短链脂肪酸、膳食纤维、蛋白质、总酚、游离氨基酸、矿物质元素等营养功能成分的影响。结果显示,乳酸菌可提高甘薯渣发酵产物中乳酸、乙酸、丙酸、总膳食纤维、可溶性膳食纤维、总酚、游离氨基酸及矿物质元素的含量,且不同菌种对甘薯渣的发酵能力存在显著差异。对不同菌种发酵产物进行灰色理论加权关联度分析,结果显示保加利亚乳杆菌和干酪乳杆菌发酵甘薯渣产物的综合营养价值较高。
猜你喜欢
湖北农机化(2021年11期)2021-12-07
军事文摘(2021年18期)2021-12-02
保鲜与加工(2021年1期)2021-02-06
中国酿造(2020年10期)2020-11-04
当代水产(2019年11期)2019-12-23
今日农业(2019年16期)2019-01-03
小学阅读指南·低年级版(2016年10期)2016-09-10
中国洗涤用品工业(2015年7期)2015-02-28
中国酿造(2014年9期)2014-03-11
中国酿造(2014年9期)2014-03-11
